链格孢引起的病害严重危害农作物生产并危及农产品安全
2017-08-09翟伟卜齐放军
王 婧, 翟伟卜, 高 环, 韩 榕, 齐放军*
(1. 山西师范大学生命科学学院, 临汾 041004; 2. 中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室, 北京 100193)
链格孢引起的病害严重危害农作物生产并危及农产品安全
王 婧1,2, 翟伟卜2, 高 环2, 韩 榕1*, 齐放军2*
(1. 山西师范大学生命科学学院, 临汾 041004; 2. 中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室, 北京 100193)
链格孢属AlternariaNees真菌是一类在自然环境中广泛分布的真菌,该属的一些种可引起多种重要农作物病害发生,严重危害农作物生产并造成巨大损失。另外,这些链格孢产生的毒素还积聚在农产品中,危及农产品食用安全。为此有必要加强有关链格孢病害的研究和治理。本文介绍了有关链格孢分类研究进展,近些年来链格孢危害农作物的主要种类及危害情况,以及世界范围内链格孢毒素所引发的农产品食用安全问题。在此基础上,也综述了链格孢病害的主要防控策略和治理技术。
链格孢病害; 毒素; 农作物生产; 农产品安全
链格孢属于子囊菌Ascomycota座囊菌纲Dothideomycetes格孢腔菌目Pleosporales格孢腔菌科Pleosporaceae链格孢属Alternaria[1]。链格孢广泛分布在土壤、空气和农作物残体中[2],主要以腐生、内生或致病三种形式存在。其中植物病原菌可侵染多种农作物,如小麦、水稻和高粱等谷类作物;油菜、大豆、向日葵等油料作物;番茄、马铃薯、茄子、辣椒、苹果、梨等蔬菜瓜果;芦荟、菊花等观赏花卉;人参、甘草、田七等中草药材;还有棉花、烟草等经济作物,给农业生产带来巨大损失[3]。链格孢除了引起农作物病害外,还极易引起农产品霉变,导致蔬菜、瓜果和储存食品腐败变质[4]。有些链格孢还具有产毒能力,它们产生的植物毒素主要分为两类:即寄主专化性毒素(host-selective toxins, HSTs)和非寄主专化性毒素(nonhost-selective toxins, NHSTs)。这些毒素累积在农产品和食物中,严重威胁农产品的食用安全。如欧洲曾对小麦中的链格孢毒素进行了长达十年的监测,发现各国小麦样品被毒素污染的情况较为严重[5]。一些谷类作物:小麦、高粱、大麦、燕麦等;瓜果蔬菜:苹果、柑橘、番茄、辣椒等;酒水饮品:苹果汁、葡萄汁、红酒等;食用油:橄榄油、菜籽油、葵花油、芝麻油等,以及其他农产品中均检测出链格孢毒素。此外,链格孢的孢子还可以释放过敏原[6]引发哮喘病[7]等,严重威胁人类的健康。链格孢引起的农作物病害危害如此严重,加强有关链格孢的研究并建立起有针对性的防控策略和技术,对保障农业生产、保证农产品食用安全显得尤为重要。
1 链格孢属的分类研究
1816年Nees[8]以细链格孢Alternariatenuissima为模式种,最先提出并描述了链格孢属的特征:暗色,具横、纵隔膜的孢子成链着生,在较细的顶端具喙。该属提出时,分类地位暂不明确,随后该属的分类地位又历经变迁。1832年Fries[9]在他所写的《Systema Mycologicum》中否认了Nees对于链格孢Alternaria的描述,并把其与Torulaalternate合并为一类,并提出一个新属Macrosporium。1912年Keissler[10]在Nees和Fries的基础上,又将A.alternata与A.tenuissima和Torulaalternate归为一类。同时,Stemphylium和Ulocladium属也被描述为丝状菌类中的有隔孢子类型。此后,因为越来越多新种的发现,使该真菌类群的分类更加复杂化。直至1917年,Elliott[11]研究了链格孢属各种类后,界定该属的典型特征是分生孢子倒棒状,具长喙,而孢子成链的特征不稳定,易受环境条件影响,有关链格孢属这一形态特征的界定得到人们认可。1933年Wiltshire[12]在前人研究的基础上,进一步比较了Alternaria和Macrosporium的基本种,建议将Macrosporium废除,使用Alternaria为该属名称,该建议为大家所认同。至此,链格孢属的分类地位大致确定下来。基于对链格孢属系统的研究,Simmons对链格孢属的形态学特征进行了全面概括和归纳[13]。我国张天宇也出版了专著《中国真菌志-链格孢属》[14]。
一直以来,链格孢种级分类难度较大。传统的种级分类主要是依据分生孢子和产孢结构的形态,包括分生孢子的形状、大小、分隔、颜色、表面纹饰、喙的有无及形态。菌丝特征、菌落特征、寄主范围等作为分类的辅助依据。但来自寄主和来自培养基上的链格孢(特别是小孢子种)分生孢子形态具有较大差异。在培养条件下,分生孢子形态易受温湿度、光照等培养条件的影响,而且小孢子种的孢子形态有趋同的倾向,这给准确的种级分类鉴定带来了很大困难[15-16]。
鉴于从形态学特征难以对链格孢进行种级划分,随着分子生物学技术的发展,分子序列特征分析已发展成为对链格孢属种类进行鉴定和分类的重要手段。目前多采用保守基因序列对Alternaria进行系统学分析,且逐渐由单基因分析向多基因联合分析发展(表1)。
表1 链格孢分类中主要采用的基因序列
Table 1 Gene sequences mainly used in classification ofAlternaria
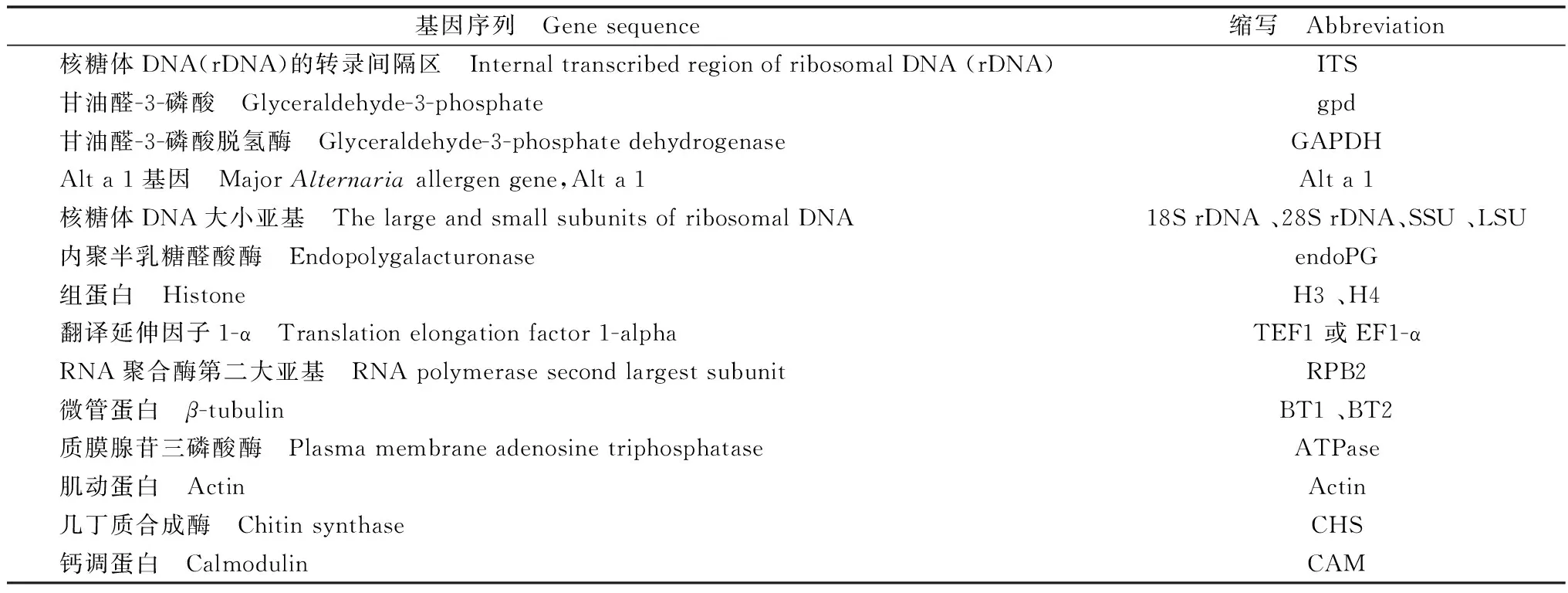
基因序列 Genesequence缩写 Abbreviation核糖体DNA(rDNA)的转录间隔区 InternaltranscribedregionofribosomalDNA(rDNA)ITS甘油醛-3-磷酸 Glyceraldehyde-3-phosphategpd甘油醛-3-磷酸脱氢酶 Glyceraldehyde-3-phosphatedehydrogenaseGAPDHAlta1基因 MajorAlternariaallergengene,Alta1Alta1核糖体DNA大小亚基 ThelargeandsmallsubunitsofribosomalDNA18SrDNA、28SrDNA、SSU、LSU内聚半乳糖醛酸酶 EndopolygalacturonaseendoPG组蛋白 HistoneH3、H4翻译延伸因子1-α Translationelongationfactor1-alphaTEF1或EF1-αRNA聚合酶第二大亚基 RNApolymerasesecondlargestsubunitRPB2微管蛋白 β-tubulinBT1、BT2质膜腺苷三磷酸酶 PlasmamembraneadenosinetriphosphataseATPase肌动蛋白 ActinActin几丁质合成酶 ChitinsynthaseCHS钙调蛋白 CalmodulinCAM
Woudenberg等曾利用GAPDH、RPB2和TEF1三种基因序列的数据,对121株链格孢进行了系统分类学分析并建立了系统发育树[1]。Lawrence等结合gpd、Alt al、Actin、Plasma membrane ATPase和Calmodulin 5个蛋白基因编码位点的数据,对176个从多个资源库中收集到的链格孢种及其相近种进行系统学分析并建树,揭示了8个特征区分明显的链格孢无性繁殖系[17]。Hou等对两株从烟草中分离得到的链格孢进行了全基因组测序分析和系统发育分析,发现A.longipescx1和A.alternatacx2在330万年前就发生了分离,并还发现了一些可能与非寄主专化性毒素合成有关的基因[18]。代谢组学也开始应用于链格孢的分类研究中。Andersen等利用HPLC技术对分离得到的153株链格孢产生的次级代谢产物进行分析,最终根据所产生次级代谢产物的不同,结合形态学特征,将其分为三个链格孢菌群A.infectoriaspecies-group,A.arborescensspecies-group和A.tenuissimaspecies-group[19]。
2 链格孢对主要农作物的侵染危害情况
据不完全统计,链格孢属病原菌可侵染的农作物和林果植物多达200种以上。下面分别介绍此类病原菌对谷类作物、油料作物、蔬菜瓜果、观赏花卉、中草药材以及其他经济作物的侵染危害情况。
2.1 谷类作物
小麦是重要的粮食作物,近年来,随着我国矮秆小麦品种的推广种植,土壤肥力的提高,肥水投入的增加及成熟期气候变暖,链格孢引起的小麦黑胚病在我国不时发生,严重影响小麦的产量和品质。据报道我国小麦主产区河南省有些品种受到链格孢A.alternata、细极链格孢A.tenuissima、小麦链格孢A.triticina的侵染,经济损失严重[20-21]。Webley等于1995-1996年调查了澳大利亚小麦、大麦、燕麦和高粱等作物受链格孢侵染的情况,发现这些作物均不同程度受到危害[22]。小麦黑胚病还曾在德国和阿根廷等国家暴发,导致小麦产量损失惨重,为此欧洲长期监测链格孢对小麦的危害。
2.2 油料作物
油菜黑斑病是油菜上常见病害之一,目前已知的致病菌有芸薹链格孢A.brassicae、甘蓝链格孢A.brassicicola和萝卜链格孢A.raphani3种,在我国各油菜产区均有发生。西藏因其特殊的高原气候,当地油菜品种具有抗寒、抗旱、耐贫瘠、含油量高等特点,但随着种植面积的扩大,西藏地区的油菜黑斑病也不断加重[23]。
向日葵为世界第五大油料作物。在我国向日葵是第四大油料作物,其栽培面积仅次于大豆、油菜和花生,约为117万hm2,主要种植地区为内蒙古、黑龙江、山西、新疆、吉林等省/自治区[24]。引起向日葵黑斑病的病原菌有8种,其中向日葵链格孢A.helianthi为优势病原菌[25]。而在印度,向日葵黑斑病同样危害严重,可导致结籽率降低30%~80%,出油率下降30%左右[26]。泰国首次发现长柄链格孢A.longipes可侵染油棕并致其发生叶斑病[27]。
2.3 蔬菜瓜果
番茄、马铃薯早疫病,又称为“轮纹病”,是由茄链格孢A.solani所导致的一种危害严重的世界性植物真菌病害[28];茄子、辣椒早疫病的病原菌也是茄链格孢。我们日常餐桌上的胡萝卜[29-30]、洋葱[31]、卷心菜[32]、芥菜等蔬菜也均可被链格孢属真菌侵染。
苹果落叶病这一先前危害并不严重的病害,目前在我国很多果树种植区呈现严重的发病态势。据调查不少果园平均病株率高达55.6%~72.8%,病株率达100%的果园也很常见,该病害一旦发生,大量叶片过早脱落,树势趋衰[33]。链格孢引起的苹果落叶病对意大利南部果园危害同样严重[34]。Zhao等在北京首次发现了由细极链格孢A.tenuissima引起的甜瓜叶部病害,且发病率达到33.3%~87.5%[35]。 我国新疆的库尔勒香梨[36]、河北鸭梨[37]、中华猕猴桃[38]、大樱桃[39]、柑橘[40]等瓜果上也发现了链格孢引发的病害。美国加利福尼亚州的开心果[41]、阿根廷的蓝莓[42]受链格孢危害严重。
2.4 观赏花卉
银胶菊Partheniumhysterophorus是印度主要鲜切花之一,具有重要的商业价值,近年来银胶菊早疫病发病率呈增长趋势,Kaur等首次分离并鉴定其病原菌是大孢链格孢A.macrospora[43]。而同为菊科的万寿菊Tageteserecta是一种集观赏、药用、抗菌等多种用途的作物,在我国种植面积呈逐年扩大的趋势,而链格孢引起的万寿菊黑斑病发病严重,种植田块发病率一般在20%~30%,严重时可达95%以上,尤其是7月中旬以后显著加重,而且蔓延迅速,短时间内即可蔓延至全田,造成严重损失[44-45]。
芦荟Aloeveravar.chinensis(Haw.) Berg是集观赏、医疗、食用等多种功能于一身的热带植物,也是近年来出现的具有巨大发展潜力的药用花卉资源之一。有研究表明,倒梨形链格孢A.obpyriformis侵染导致芦荟黑斑病发生[46]。
2.5 中草药材
甘草这一具有重要经济价值的常用中药材,随着近些年来种植面积增加,由豆链格孢A.azukiae引起的甘草叶斑病的发生也日趋严重,成为影响甘草生产的主要障碍。甘草叶斑病严重发生时病叶率高达80.0%以上,光合作用受阻,产量明显降低,给种植农户造成了严重的经济损失。豆链格孢A.azukiae还可侵染曼陀罗、藤三七、独活等药材[47]。2008年―2010年,南京地区种植的传统中药材千金子早疫病发生严重,其致病菌也是链格孢A.alternata[48]。在韩国,人参链格孢A.panax对高丽参的危害一直是这一重要中药材种植所面临的难题[49]。
2.6 其他经济作物
Zhao等发现,链格孢A.alternata易于侵染受低温伤害的棉花叶片[50],引起棉花叶片轮纹斑病,最终导致棉花早衰。低温和链格孢复合作用引起的棉花早衰,一度遍及我国各主要棉区,严重危害我国棉花生产,造成巨大损失[51]。而在澳大利亚北部地区,由链格孢引起的棉花叶部病害[52]也曾广泛流行,且该病害可贯穿于棉花生长的整个周期,危害十分严重。Bhuiyan等采用喷洒农药代森锰锌和硝酸钾的方法来控制病害的发生,并取得了一定的成效[53]。链格孢A.alternata还可侵染烟草,引起烟草赤星病的暴发,严重威胁烟草生产[54-55]。
3 链格孢毒素
一些链格孢产生的链格孢毒素还严重危及农产品的食用安全。人或动物一旦摄入链格孢毒素污染的食物,可能导致急性或慢性中毒,某些链格孢毒素还有致畸、致癌、致突变作用[56]。
3.1 寄主专化性毒素
由植物病原真菌产生的寄主专化性毒素(host-selective toxins, HSTs)多为低分子量的次生代谢物,有多种多样的结构,在植物-病原互作中往往作为病原菌的效应子,影响病原菌的致病性[57]。
1933年,Tanaka首次在日本梨黑斑病中发现了参与植物-病原物互作的HST。梨黑斑病菌专化性侵染‘Nijisseiki’梨品种。‘Nijisseiki’梨是1888年在日本千叶县一个潮湿的院子里发现的自然突变株,在其被发现20年后才开始经济栽培。因‘Nijisseiki’对菊池链格孢A.kikuchiana极其敏感,黑斑病一直严重威胁着‘Nijisseiki’梨的生产。Tanaka还发现,真菌滤液中含有危害‘Nijisseiki’梨的毒素,但该毒素并不危害抗性梨品种。这是第一个通过培养滤液的方法证实专化性毒素的例子,随后该毒素被分离并鉴定为AK-toxin[57-59]。
目前仅在真菌中发现寄主专化性毒素(HSTs),且不超过20种,它们都是一些小分子的次生代谢产物,其中由链格孢A.alternata产生的HSTs多达7种(表2)。这7种毒素分别由其相应的病原菌致病型产生,而这些病原菌致病型则是依据其所侵染的寄主命名的[60]。
表2 已知的由链格孢产生的寄主专化性毒素(HSTs)
Table 2 Host-selective toxins (HSTs) produced byAlternariaalternata

病害 Disease致病型 Pathotype毒素 Toxin苹果斑点落叶病 Alternariablotchofapple苹果致病型 ApplepathotypeAM-toxinⅠⅡ,Ⅲ草莓黑斑病 Blackspotofstrawberry草莓致病型 StrawberrypathotypeAF-toxinⅠⅡ,Ⅲ日本梨黑斑病 BlackspotofJapanesepear日本梨致病型 JapanesepearpathotypeAK-toxinⅠ,Ⅱ柑橘褐斑病 Brownspotoftangerine橘致病型 TangerinepathotypeACT-toxinⅠ,Ⅱ粗柠檬叶斑病 Leafspotofroughlemon粗柠檬致病型 RoughlemonpathotypeACR-toxinI番茄早疫病 Alternariastemcankeroftomato番茄致病型 TomatopathotypeAAL-toxinTa,Tb烟草赤星病 Brownspotoftobacco烟草致病型 TobaccopathotypeAT-toxin
HSTs作为专化性毒素,只危害特定种类植物,而对于其他种类的植物,HSTs则作为效应子激发植物防御反应的产生。HSTs毒素在病原菌分生孢子萌发过程中产生,并在侵染过程中发挥作用。目前在植物细胞中明确了HSTs毒素的4个作用靶点:质膜、线粒体、叶绿体和内质网上的鞘磷脂合成酶(ceramide synthase)。其中AM-toxin作用于叶绿体、质膜;AK-toxin、AF-toxin、ACT-toxin作用于质膜;ACR-toxin、AT-toxin作用于线粒体;AAL-toxin作用于内质网的鞘磷脂合成酶[58]。
3.2 非寄主专化性毒素
链格孢产生的各类非寄主专化性毒素(nonhost-selective toxins, NHSTs)可导致各种农产品腐烂变质,并危及农产品的食用安全[61]。链格孢产生的NHSTs主要有7种,分别是:链格孢酚(alternariol, AOH)、交链孢酚单甲醚(alternariol monomethyl ether, AME)、链格孢霉素(altenuene, ALT)、细格菌毒素Ⅰ,Ⅱ,Ⅲ(altertoxins Ⅰ,Ⅱ,Ⅲ, ATX-Ⅰ,Ⅱ,Ⅲ)、细交链孢菌酮酸(tenuazonic acid, TeA)[4]
4 链格孢毒素引发的农产品安全问题
链格孢毒素存在谷类作物中,危及主粮的食用安全。Müller等[5]于2001年―2010年,调查了1 064个从德国勃兰登堡地区收获的冬小麦样本,结果发现有30.3%的样本检测出细交链孢菌酮酸(TeA),8.1%的样本检测出链格孢酚(AOH),3.1%的样本检测出交链孢酚单甲醚(AME),2.6%的样本检测出链格孢霉素(ALT)。TeA是该地区最为常见的毒素,每年都能在小麦样本中检测到,2001-2010年十年间,该毒素从小麦样本的检出率从1%~4%突增至100%,呈现暴发式发展趋势[5]。
而Hajnal等于2011年―2013年对塞尔维亚地区小麦含链格孢毒素情况进行了检测,发现68.5%的样本含细交链孢菌酮酸(TeA),12%的样本检出链格孢酚(AOH),6.5%的样本检出交链孢酚单甲醚(AME)[62]。
阿根廷是欧洲新鲜水果和加工食品的第二大出口国,从阿根廷出口的谷类、水果和蔬菜中均检出链格孢毒素和其他一些次生代谢产物。Andersen等对阿根廷地区番茄、小麦、蓝莓和核桃中链格孢的产毒能力进行了研究,结果发现,在其所检测的菌系中至少有75%有潜在的产毒能力[63]。
Scott报道了链格孢毒素在各类农产品中的检出情况。其中AOH、AME和TeA三种毒素通常可以在苹果、苹果制品、苹果汁、柑橘、橄榄、辣椒、红辣椒、番茄、番茄制品、向日葵籽、高粱、小麦、阿根廷蓝莓和食用油(橄榄油、菜籽油、芝麻油、葵花籽油)中检测到。AOH和AME在西梅汁、树莓、红醋栗、大麦、燕麦、日本梨、柑橘类和胡萝卜中检测到。AME和TeA在柠檬中被检测到。AOH在黑莓、草莓、蔓越莓汁、葡萄汁、覆盆子汁等果汁和红酒中一般也可以检测到。ATX-Ⅰ和ATX-Ⅱ在链格孢侵染的苹果、高粱和小麦中也被检测到[64-65]。
5 链格孢病害的防控策略
链格孢引起农作物病害的暴发流行与品种、气候、逆境、土壤、耕作模式等许多因素有关。近年来报道的危害作物的链格孢种类逐年增多,危害越发严重,加上链格孢产生的毒素还严重危及各类农产品的食用安全,为此防控链格孢病害的发生和危害已成为保障农业安全生产的重要研究课题之一。对于不同种类不同经济价值的作物,可以采取不同的防控策略。
在北欧马铃薯早疫病呈现逐年增长的趋势。Runno-Paurson等对2010年和2011年当地种植的马铃薯进行了调查,发现感病品种发病极为严重,2010年马铃薯幼苗的发病率达到了70%,而2011年马铃薯幼苗早疫病的发病更为严重,发病率从2%暴增至91%,五周之内马铃薯幼苗全部死亡。相比较而言,抗病品种基本不发病或发病轻。由此可见,培育种植抗病的马铃薯品种是早疫病防治的首要任务[66]。在韩国,分别利用防和治两种方法控制高丽参发生链格孢病害。Kim等先利用避雨遮阳的工具、除去死掉的植株等方法预防病害发生,再喷洒一些杀菌剂控制病情的发展[49]。冬枣在贮藏过程中经常会因链格孢侵染而腐烂,造成相当大的损失,Yan等研究表明,将采摘后的冬枣蘸取一定量的氨基丁酸(β-aminobutyric acid, BABA),可以有效地控制冬枣腐烂病害的发生,并且能提高贮藏冬枣的品质[67]。de Lima等发现大蒜提取物和橘子精油对胡萝卜链格孢A.dauci和链格孢A.alternata有一定控制作用[68]。
链格孢种类繁多,分布范围广泛,引起的病害种类多,危害广,已成为世界范围的病害,并直接影响农产品国际贸易。就我国而言,由链格孢引起的病害暴发在近些年显得尤为突出。由于对该病害的认识不足,研究投入缺乏,使得广大农户在病害发生后不仅不能明晰病因,也缺乏有效的防控措施,导致病情逐年递增,损失惨重,已成为我国农业生产中病害防控的突出问题之一。更何况链格孢毒素直接危及农产品食用安全,如何有效解决这一问题理应引起足够的重视。
[1] Woudenberg J H, Groenewald J Z, Binder M, et al.Alternariaredefined [J]. Studies in Mycology, 2013, 75(1): 171-212.
[2] 孙霞. 链格孢属真菌现代分类方法研究[D]. 泰安: 山东农业大学, 2006.
[3] Thomma B P H J.Alternariaspp.: from general saprophyte to specific parasite [J]. Molecular Plant Pathology, 2003, 4(4): 225-236.
[4] Ostry V.Alternariamycotoxins: an overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs [J].World Mycotoxin Journal,2008,1(2):175-188.
[5] Müller M E H, Korn U.Alternariamycotoxins in wheat—A 10 years survey in the Northeast of Germany [J]. Food Control, 2013, 34(1): 191-197.
[6] Mitakakis T Z, Barnes C, Tovey E R.Spore germination increases allergen release fromAlternaria[J]. The Journal of Allergy and Clinical Immunology, 2001, 107(2): 388-390.
[7] Bush R K, Prochnau J J.Alternaria-induced asthma [J]. The Journal of Allergy and Clinical Immunology, 2004, 113(2): 227-234.
[8] Nees von Esenbeck C G D.Das system der Pilze und Schwämme: ein versuch [M]. Wurzburg, Germany, 1817.
[9] Fries E M. Systema mycologicum […] Volumen Ⅲ[M]. Germany: Gryphiswaldae [sic]: Sumtibus Ernesti Maurittii, 1829: 419-425.
[10]Keissler K. Zur kenntnis der pilzflora krains [J]. Beihefte zum Botanischen Zentralblatt, 1912, 29: 395-440.
[11]Elliott J A.Taxonomic characters of the generaAlternariaandMacrosporium[J]. American Journal of Botany, 1917, 4(8): 439-476.
[12]Wiltshire S P.The foundation species ofAlternariaandMacrosporium[J]. Transactions British Mycological Society, 1933, 18: 135-160.
[13]Simmons E G.Alternaria: an identification manual [M]. The Netherlands Utrecht: CBS Fungal Biodiversity Centre, Utrecht, 2007.
[14]张天宇. 中国真菌志, 第16卷: 链格孢属[M]. 北京: 科学出版社, 2003.
[15]Roberts R G, Reymond S T, Andersen B.RAPD fragment pattern analysis and morphological segregation of small-sporedAlternariaspecies and species groups [J]. Mycological Research, 2000, 104(2): 151-160.
[16]Tymon L S, Peever T L, Johnson D A.Identification and enumeration of small-sporedAlternariaspecies associated with potato in the U.S. Northwest [J]. Plant Disease, 2016, 100(2): 465-472.
[17]Lawrence D P, Gannibal P B, Peever T L, et al. The sections ofAlternaria: formalizing species-group concepts [J]. Mycologia, 2013, 105(3): 530-546.
[18]Hou Yujie, Ma Xiao, Wan Wenting, et al. Comparative genomics of pathogens causing brown spot disease of tobacco:AlternarialongipesandAlternariaalternata[J]. PLoS ONE, 2016, 11(5): e0155258.
[19]Andersen B, Krøger E, Roberts R G.Chemical and morphological segregation ofAlternariaarborescens,A.infectoriaandA.tenuissimaspecies-groups [J]. Mycological Research, 2002, 106(2): 170-182.
[20]康业斌, 张有聚, 李会娟, 等. 我国小麦黑胚病研究现状[J]. 麦类作物学报, 1999(2): 58-60.
[21]代君丽, 于巧丽, 袁虹霞, 等. 河南省小麦黑胚病菌的分离鉴定及致病性测定[J]. 植物病理学报, 2011, 41(3): 225-231.
[22]Webley D J, Jackson K L, Mullins J D, et al.Alternariatoxins in weather-damaged wheat and sorghum in the 1995-1996 Australian harvest [J]. Australian Journal of Agricultural Research, 1997, 48(8): 1249-1255.
[23]岳海梅, 庄华, 巩文峰, 等. 一株西藏白菜型黄籽油菜黑斑病菌的鉴定及生物学特性[J]. 中国油料作物学报, 2015, 37(3): 377-382.
[24]中国农业年鉴编辑委员会. 中国农业年鉴[M]. 北京: 中国农业出版社, 2006: 462-464.
[25]兰巍巍, 陈倩, 王文君, 等. 向日葵黑斑病研究进展及其综合防治[J]. 植物保护, 2009, 35(5): 24-29.
[26]Udayashankar A C, Nayaka S C, Archana B, et al. Specific PCR-based detection ofAlternariahelianthi: the cause of blight and leaf spot in sunflower [J]. Archives of Microbiology, 2012, 194(11): 923-932.
[27]Suwannarach N, Kumla J, Lumyong S. First report ofAlternarialeaf blight disease on oil palm caused byAlternarialongipesin Thailand [J]. Phytoparasitica, 2014, 43(1): 57-59.
[28]Patel S J, Subramanian R B, Jha Y S. Biochemical and molecular studies of early blight disease in tomato [J]. Phytoparasitica, 2011, 39(3): 269-283.
[29]Soylu S, Kurt S, Soylu E M, et al. First report ofAlternarialeaf blight caused byAlternariadaucion carrot in turkey [J]. Plant Pathology, 2005, 54: 252.
[30]Farrar J J, Pryor B A, Davis R M.Alternariadiseases of carrot [J]. Plant Disease, 2004, 88(8): 776-784.
[31]Bihon W, Cloete M, Gerrano A S, et al. Draft genome sequence ofAlternariaalternataisolated from onion leaves in South Africa[J]. Genome Announcements, 2016, 4(5): e01022-16.
[32]Chen L Y, Price T V, Silvapulle M J.Dark leaf spot (Alternariabrassicicola) on Chinese cabbage: spatial patterns [J].Australian Journal of Agricultural Research, 2005, 56(7): 699-714.
[33]王岩, 孙丽梅. 苹果斑点落叶病的发生与防治[J]. 北方果树, 2015(2): 43.
[34]Rotondo F, Collina M, Brunelli A, et al. Comparison ofAlternariaspp. collected in Italy from apple withA.maliand other AM-toxin producing strains [J]. Phytopathology, 2012, 102(12): 1130-1142.
[35]Zhao Juan, Bao Shuwen, Ma Guoping, et al. Characterization ofAlternariaspecies associated with muskmelon foliar diseases in Beijing municipality of China[J]. Journal of General Plant Pathology, 2015, 82(1): 29-32.
[36]宋博, 朱晓锋, 徐兵强, 等. 库尔勒香梨果萼黑斑病病原鉴定及其ITS、GPD和EF-1α序列分析[J]. 园艺学报, 2016, 43(2): 329-336.
[37]严进, 施宗伟, 宋福, 等. 河北和山东鸭梨果实上链格孢菌鉴定[J]. 植物保护学报, 2009, 36(1): 37-43.
[38]赵金梅,高贵田,谷留杰,等.中华猕猴桃褐斑病病原鉴定及抑菌药剂筛选[J].中国农业科学,2013,46(23):4916-4925.
[39]赵远征, 刘志恒, 李俞涛, 等. 大樱桃黑斑病病原鉴定及其致病性研究[J]. 园艺学报, 2013, 40(8): 1560-1566.
[40]Akimitsu K, Peever T L, Timmer L W.Molecular, ecological and evolutionary approaches to understandingAlternariadiseases of citrus [J].Molecular Plant Pathology, 2003, 4(6): 435-446.
[41]Ma Zhonghua, Felts D, Michailides T J.Resistance to azoxystrobin inAlternariaisolates from pistachio in California[J]. Pesticide Biochemistry and Physiology, 2003, 77(2): 66-74.
[42]Greco M, Patriarca A, Terminiello L, et al. ToxigenicAlternariaspecies from Argentinean blueberries[J]. International Journal of Food Microbiology, 2012, 154(3): 187-191.
[43]Kaur M, Aggarwal N K. First report ofAlternariasp. causing leaf blight disease on parthenium weed in India [J]. New Disease Reports, 2015, 31: 25.
[44]冯倩倩, 陈东亮, 程曦, 等. 万寿菊黑斑病的研究进展[J]. 北方园艺, 2015(20): 181-185.
[45]Li Y, Shen J, Pan B H, et al. First report of leaf spot caused byAlternariaalternataon marigold (Tageteserecta) in Beijing, China [J]. Plant Disease, 2014, 98(8): 1153-1154.
[46]鲁红学, 赵明敏, 李建强. 芦荟黑斑病病原鉴定及培养特性的研究[J]. 华北农学报, 2007, 22(1): 156-160.
[47]阎合, 徐秉良, 梁巧兰, 等. 甘草叶斑病的发生与病原菌鉴定[J]. 植物保护, 2009, 35(3): 111-114.
[48]Yu X, Zhang W M, Zhao B T, et al. First report ofAlternariaalternatacausing a blight disease ofEuphorbialathyrisin China [J]. Journal of Plant Pathology, 2011, 93(4): 85.
[49]Kim Y C, Lee J H, Bae Y S, et al. Development of effective environmentally-friendly approaches to controlAlternariablight and anthracnose diseases of Korean ginseng [J]. European Journal of Plant Pathology, 2010, 127(4): 443-450.
[50]Zhao Jingqing, Li Sha, Jiang Tengfei, et al. Chilling stress—The key predisposing factor for causingAlternariaalternatainfection and leading to cotton (GossypiumhirsutumL.) leaf senescence [J]. PLoS ONE, 2012, 7(4): e36126.
[51]齐放军, 简桂良, 李家胜. 棉花早衰、红叶茎枯病与棉花轮纹斑病间关系辨析[J]. 棉花学报, 2013, 25(1): 81-85.
[52]Bhuiyan S A, Boyd M C, Martin C, et al. Development ofAlternarialeaf blight on north Australian cotton (Gossypiumhirsutum), species prevalence, and its control using mancozeb[J]. Australian Plant Pathology, 2007, 36(5): 488-497.
[53]Bhuiyan S A, Boyd M C, Dougall A J, et al. Effects of foliar application of potassium nitrate on suppression of Alternaria leaf blight of cotton (Gossypiumhirsutum) in northernAustralia[J].Australian Plant Pathology,2007,36(5):462-465.
[54]Sun Huanhuan, Wang Lei, Zhang Baoqin, et al. Scopoletin is a phytoalexin againstAlternariaalternatain wild tobacco dependent on jasmonate signalling [J]. Journal of Experimental Botany, 2014, 65(15): 4305-4315.
[55]Sun H, Hu X, Ma J, et al. Requirement of ABA signalling-mediated stomatal closure for resistance of wild tobacco toAlternariaalternata[J].Plant Pathology,2014,63(5):1070-1077.
[56]Bensassi F, Gallerne C, El Dein O S, et al. Cell death induced by theAlternariamycotoxin Alternariol [J]. Toxicology in Vitro, 2012, 26(6): 915-923.
[57]Akimitsu K, Tsuge T, Kodama M, et al.Alternariahost-selective toxins: determinant factors of plant disease [J]. Journal of General Plant Pathology, 2014, 80(2): 109-122.
[58]Tsuge T, Harimoto Y, Akimitsu K, et al. Host-selective toxins produced by the plant pathogenic fungusAlternariaalternata[J]. Federation of European Microbiological Societies Microbiology Reviews, 2013, 37(1): 44-66.
[59]Tanaka A, Shiotani H, Yamamoto M, et al. Insertional mutagenesis and cloning of the genes required for biosynthesis of the host-specific AK-toxin in the japanese pear pathotype ofAlternariaalternata[J]. Molecular Plant-Microbe Interactions, 1999, 12(8): 691-702.
[60]Walton J D.Host-selective toxins: Agents of compatibility [J]. Plant Cell, 1996, 8(10): 1723-1733.
[61]Lopez P, Venema D, Mol H, et al.Alternariatoxins and conjugates in selected foods in the Netherlands [J]. Food Control, 2016, 69: 153-159.

[63]Andersen B, Nielsen K F, Fernandez Pinto V, et al. Characterization ofAlternariastrains from Argentinean blueberry, tomato, walnut and wheat[J]. International Journal of Food Microbiology, 2015, 196: 1-10.
[64]Scott P M. Analysis of agricultural commodities and foods forAlternariamycotoxins[J]. Journal of AOAC International, 2001, 84(6): 1809-1817.
[65]Scott P M, Lawrence G A, Lau B P Y.Analysis of wines, grape juices and cranberry juices forAlternariatoxins [J]. Mycotoxin Research, 2006, 22(2): 142-147.
[66]Runno-Paurson E, Loit K, Hansen M, et al. Early blight destroys potato foliage in the northern Baltic region [J]. Acta Agriculturae Scandinavica, Section B: Soil and Plant Science, 2015, 65(5): 422-432.
[67]Yan Jiaqi, Yuan Shuzhi, Wang Chunyue, et al. Enhanced resistance of jujube (ZizyphusjujubaMill. cv. Dongzao) fruit against postharvestAlternariarot byβ-aminobutyric acid dipping [J]. Scientia Horticulturae, 2015, 186: 108-114.
[68]de Lima C B, Rentschler L L A, Bueno J T, et al. Plant extracts and essential oils on the control ofAlternariaalternata,Alternariadauciand on the germination and emergence of carrot seeds (DaucuscarotaL.) [J]. Ciência Rural, 2016, 46(5): 764-770.
(责任编辑:杨明丽)
Serious damage to crop production caused byAlternariadiseases and the safety of agricultural products
Wang Jing1,2, Zhai Weibo2, Gao Huan2, Han Rong1, Qi Fangjun2
(1.CollegeofLifeSciences,ShanxiNormalUniversity,Linfen041004,China; 2.StateKeyLaboratoryforBiologyofPlantDiseasesandInsectPests,InstituteofPlantProtection,ChineseAcademyofAgriculturalSciences,Beijing100193,China)
AlternariaNees is a widely distributed fungal genus in the natural environment. SomeAlternariaspecies are plant pathogens, causing a range of diseases on a large variety of important crops, and often leading to serious damage to the agricultural production. These plant pathogenicAlternariaspecies often produceAlternariatoxins, which may accumulate in agricultural products, and endanger the safety of edible agricultural products. Therefore, it is necessary to strengthen the studies on the occurrence and management ofAlternariadiseases. Here, progresses inAlternariaspecies classification, the main crops damaged byAlternariadiseases, and the worldwide issues of food and agricultural product safety caused byAlternariatoxins are introduced in this paper. Furthermore, strategies and technologies for management ofAlternariadiseases are also reviewed.
Alternariadisease; toxin; crops production; safety of agricultural products

专论与综述Reviews
2016-10-18
2016-12-07
国家自然科学基金(31371898); 植物病虫害生物学国家重点实验室开放基金(SKLOF201615)
S 432.44
A
10.3969/j.issn.0529-1542.2017.04.002
* 通信作者 E-mail: hanrong@dns.sxnu.edu.cn;fjqi@ippcaas.cn
