骨碎补活性单体成分调控骨质疏松症相关信号通路的研究进展
2017-08-06招文华沈耿杨任辉丘婷张志达唐晶晶陈康梁德姚珍松杨志东江晓兵
招文华 沈耿杨 任辉 丘婷 张志达 唐晶晶 陈康 梁德 姚珍松 杨志东 江晓兵*
1. 广州中医药大学,广东 广州 510405 2. 广州中医药大学第一附属医院,广东 广州 510405
骨质疏松症(osteoporosis,OP)是一种严重危害人类健康的代谢性骨病,具有高发病率、高致残率的特点[1]。目前,针对OP的治疗趋向多样化,但仍以西药治疗为主,如降钙素、双膦酸盐、氟化物等。然而,这些西药大多数都具有一定副作用,如降钙素可引起恶心、呕吐、面部潮红、手部麻刺感及低血钙造成的四肢搐搦等不良反应,当应用动物来源的降钙素时,还可引起过敏反应,而双膦酸盐除了可引起恶心、呕吐、腹痛、肌肉骨骼痛等不良反应外,还有可能引起颌骨坏死、食管癌和肾功能衰竭等严重不良反应。因而,寻求高效、安全、价格低廉的抗骨质疏松新药仍迫在眉睫。长期以来,临床运用中药治疗OP显示出独特优势,能明显缓解患者症状,提高骨密度,改善生活质量[2]。近年来,骨碎补在促进骨折愈合、防治骨质疏松等方面的药理作用逐渐引起了国内外学者的关注,同时大量实验研究显示,骨碎补能有效抑制骨吸收与促进骨形成[3,4]。而OP的发生,实质就是骨吸收和骨形成过程中的动态平衡被打破,引起骨组织生理代谢发生变化,继而使得骨组织结构和力学特征发生改变的过程。目前的研究显示,Wnt/β-catenin、骨保护素/细胞核因子kB受体活化因子配体/细胞核因子kB受体活化因子(OPG/RANKL/RANK)、组织蛋白酶K(CTSK)、骨形态发生蛋白(BMP)等分子信号通路在骨吸收和骨形成中起着重要的调控作用。基于以上信号通路,近年来研究者进行了大量有关骨碎补活性单体成分防治OP的研究,并取得可喜成果,本文将综述骨碎补活性单体成分调控OP相关信号通路的研究进展。
1 骨碎补抗OP的活性单体成分研究
研究表明,从骨碎补中分离得到的化学成分类型主要包括黄酮、三萜、酚酸及其苷类等[5],酚酸类也称苯丙素类。其中,关于骨碎补抗OP活性的研究主要集中于总黄酮。骨碎补中的黄酮类化合物主要有二氢黄酮、黄烷-3-醇(又称儿茶素类)、黄酮、黄酮醇、色原酮、查尔酮、橙酮类及其衍生物等。近年来,从骨碎补的乙醇提取物中已经分离得到40种黄酮类化合物。在对骨碎补总黄酮的单体成分进一步研究发现,其活性成分是以柚皮苷为主的二氢黄酮类化合物,而柚皮苷的药理作用主要是活血祛瘀、接骨疗伤和增强心肌细胞机能[6],但柚皮苷并非是骨碎补唯一的有效成分。目前,骨碎补总黄酮这一混合物中被证明具有抗OP作用的活性单体成分包括柚皮苷、柚皮素及新北美圣草苷。
1.1 柚皮苷
近年来,柚皮苷的研究是一个热点,其抗OP的活性获得了比较充分的肯定。Li 等[7]报道,柚皮苷可有效促进BMSCs的增殖和成骨分化,促进骨钙素(BGP)的表达,增强骨密度、增大骨体积及骨小梁密度从而有效逆转OP。徐展望等[8]研究发现,柚皮苷可促进体外培养BMSCs的Runx-2和 Osterix的表达,增强OP大鼠的骨强度,增大骨小梁面积。刘伟等[9]报道,1×10-5mol/L的柚皮苷可明显促进兔BMSCs的增殖并向OB分化,翟远坤等[10]发现此浓度的柚皮苷可显著促进大鼠乳鼠OB的增殖分化,并且该促进效果会被雌激素通路阻断剂ICI 182.780所抑制,表明柚皮苷属于植物雌激素,通过雌激素信号通路发挥其促骨形成活性,可以用于OP的防治。另外,李风波等[11]报道,柚皮苷可抑制OC分化、增殖和骨吸收功能,推测其机制可能是通过抑制OC分化过程中特异性基因表达实现的;他们的另一项研究[12]表明,柚皮苷可提高OP骨折大鼠的骨密度、骨体积、相对骨体积比、平均骨小梁厚度,改善骨代谢,从而增强OP骨折愈合后的力学强度。
柚皮苷除了存在于骨碎补中,还广泛存在于柚子、葡萄柚、酸橙及其变种的果皮中,我国是柚子生产大国,因而大部分的柚皮苷来源于柚子,但有研究发现,来源于骨碎补的柚皮苷的抗OP活性强于来源于柚子的柚皮苷,笔者认为今后有必要对两种来源不同的柚皮苷作进一步的研究,如它们的化学结构及活性等。
1.2 柚皮素
柚皮素是从骨碎补中分离得到的二氢黄酮类化合物,近年来研究表明其具有较明显的抗OP作用。宋双红等[13]研究报道,柚皮素可对抗卵巢切除引起的大鼠体重增加,抑制骨转换指标血清BGP和尿液脱氧吡啶啉(DPD)含量的升高,降低骨转换率,保持骨密度,改善骨应力和弹性模量等内在特性指标,同时显著改善骨小梁微结构。翟远坤等[14]通过比较研究柚皮苷及其代谢物柚皮素对体外培养乳鼠颅骨OB分化成熟的影响发现,柚皮苷和柚皮素均能明显提高乳鼠颅骨OB的碱性磷酸酶活性,刺激BGP、BMP-2、骨桥蛋白和I型胶原的分泌,增加钙化结节数量,并促进成骨相关基因mRNA表达,并发现柚皮素的活性明显高于柚皮苷,推测是柚皮苷经代谢后,其代谢产物柚皮素的促骨形成活性得到增强,这在抗OP新药的开发及剂型设计方面有一定的指导意义。
1.3 新北美圣草苷
新北美圣草苷是骨碎补中分离出的另一个主要的活性成分。实验表明,新北美圣草苷比柚皮苷在促进成骨样细胞MC3T3-E1的增殖和分化方面有更强的作用,且能够更有效的增加碱性磷酸酶的活性。通过RT-PCR发现柚皮苷和新北美圣草苷都可以上调成骨分化过程中Runx2、COLI和BGP这3个标志性基因的表达,但在新北美圣草苷作用下Runx2、COLI和BGP的表达量分别比在柚皮苷作用下的表达量上调了56%、37%和14%。此外新北圣草苷可以一定程度上逆转PD98059对细胞分化的抑制作用,可见新北圣草苷可能是骨碎补中另一个更有效的抗OP治疗的活性成分[15]。然而,针对新北美圣草苷与柚皮苷的对比研究,目前仅停留在细胞水平,缺乏动物实验支持,因而对该化合物开展分离纯化和活性研究的前景广阔。
目前,尽管骨碎补总黄酮的有效单体成分对BMSCs、OB、OC以及动物模型的作用得到深入探讨,但在相关分子信号通路上的研究仍较欠缺。
2 骨碎补调控分子信号通路抗OP的作用机制
研究发现,在骨吸收和骨形成过程中存在受多种调节因子调控的信号通路[16,17]。骨碎补通过抑制骨吸收信号通路中的调控蛋白,或促进骨形成信号通路中调控蛋白的合成和释放,改善OP中骨代谢不平衡的病理过程,从而对OP起到防治作用。目前发现骨碎补调控骨吸收的信号通路主要包括OPG/RANKL/RANK通路、CTSK通路,调控骨形成主要有Wnt/β-catenin通路和BMP通路。
2.1 OPG/RANKL/RANK信号通路
OPG/RANKL/RANK 信号通路是近年来发现在OC分化过程中起重要作用的信号传导通路[18]。OPG又称OC抑制因子,是肿瘤坏死因子受体超家族中的一员,是目前发现的唯一能直接负向调控OC的调控因子。骨组织中的OPG主要由OB谱系的各种细胞产生,并随细胞的分化成熟而增加。RANKL是OPG的配体,主要由骨组织中的OB和基质细胞分泌,在骨和骨髓中呈现高水平的表达。RANKL主要与OC表面的特异性受体RANK结合,进而刺激OC分化、活化和成熟,同时抑制OC凋亡。而OPG能与RANKL竞争性结合RANK,从而阻断由RANKL引起的OC前体分化、存活和融合,抑制成熟OC活化及骨吸收活性,最终导致OC凋亡[19-20]。因而,OPG的主要作用是抑制骨吸收,RANKL的主要作用是促进骨吸收,通过促进OPG的表达或者抑制RANKL的表达都可以抑制骨吸收,最终达到防治OP的目的。
赵晓燕等[21]报道,骨碎补水提液能上调OPG mRNA水平的表达。朱振标等[22]通过观察骨碎补总黄酮对去卵巢大鼠RANKL/OPG表达的影响发现,骨碎补总黄酮可增加去卵巢大鼠骨密度,抑制骨组织RANKL mRNA表达,促进OPG mRNA表达,最终造成骨组织RANKL含量减少,OPG含量增加,RANKL/OPG比值减小而使成骨增加。刘康等[23]用不同剂量骨碎补总黄酮对OP大鼠进行干预,发现不同剂量骨碎补总黄酮均影响OPG/RANKL/RANK轴系统,且随骨碎补总黄酮剂量的增加效果越明显,可能是通过调控OPG/RANKL/RANK轴系统使OPG表达增加、RANK和RANKL的表达下降来实现的。吴连国等[24]报道,骨碎补总黄酮可提高OP大鼠骨矿含量、降低血清RANKL水平,但不能提高血清雌二醇水平。Ang等[25]研究发现,柚皮苷可抑制OC形成和骨吸收,此过程是通过抑制RANKL表达,并且活化核因子κB(NF-κB)及磷酸化ERK实现的。
2.2 CTSK信号通路

图1 骨碎补有效成分调控OPG/RANKL/RANK信号通路Fig.1 Effective constituent of rhizoma drynariae regulating the OPG/RANKL/RANK signaling pathway
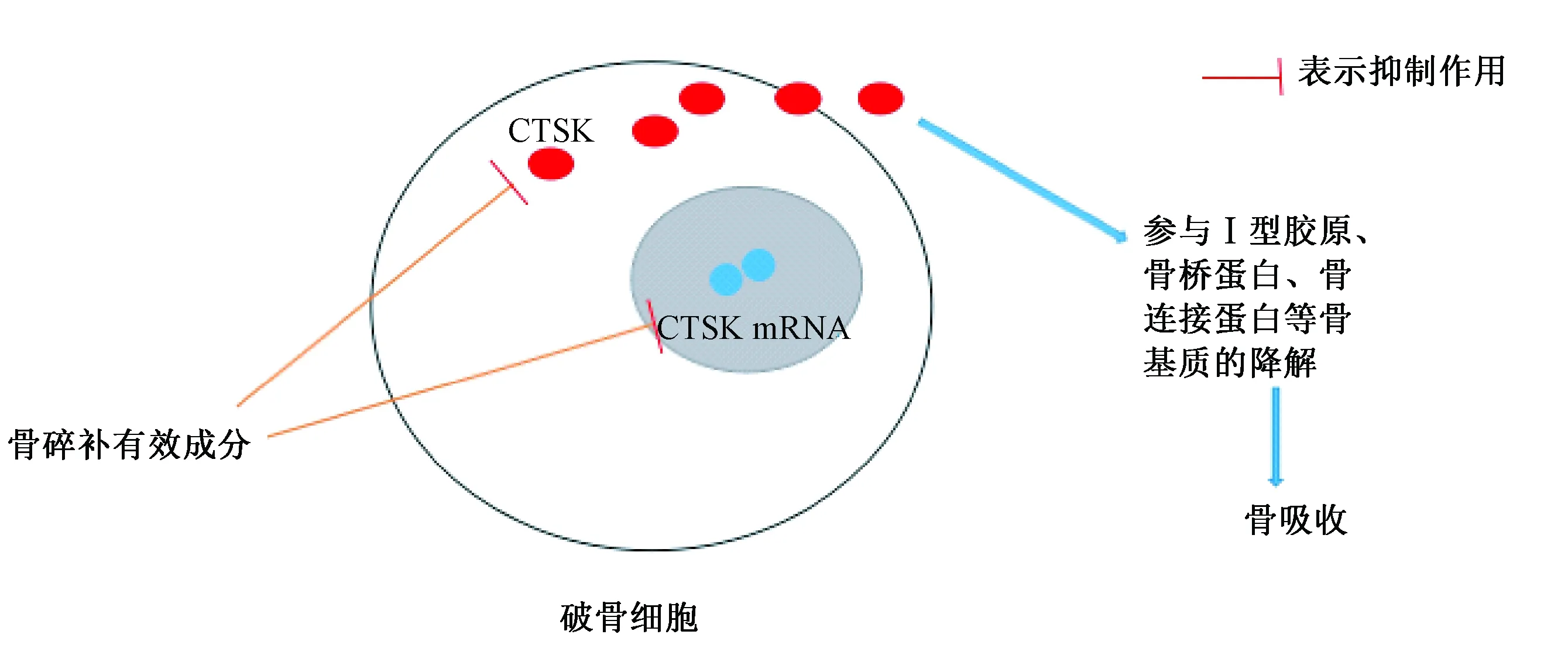
图2 骨碎补有效成分调控CTSK信号通路Fig.2 Effective constituent of rhizoma drynariae regulating the CTSK signaling pathway
CTSK是一种在OC中表达丰富的半胱氨酸蛋白酶,它是番木瓜蛋白酶家族中的一种蛋白水解酶,主要参与Ⅰ型胶原、骨桥蛋白、骨连接蛋白等骨基质的降解。CTSK主要通过调节骨胶原纤维降解来促进骨吸收,其主要作用靶点位于Ⅰ/Ⅱ型胶原纤维胶质的N端三股螺旋处[26]。因而部分学者认为通过抑制CTSK对骨组织的作用,可以达到延缓骨基质降解、改善OP的目的。
研究[27]表明,骨碎补提取物能抑制OC骨吸收,具体表现在骨吸收陷窝数和面积均呈剂量依赖性地减少,其对OC的作用可能是通过抑制细胞内CTSK的表达而起的,抑制作用与浓度呈依赖性关系。近来的一项研究[28]也认为CTSK在骨吸收中扮演着重要的角色,是骨质疏松症防治的潜在靶点,而骨碎补总黄酮能够降低CTSK的表达。而史晓林等[29]通过观察骨碎补总黄酮对去卵巢大鼠骨质疏松模型CTSK的血清浓度和左胫骨干骺端基因表达的影响发现,骨碎补总黄酮能够抑制去卵巢大鼠血清CTSK浓度、降低左胫骨干骺端CTSK mRNA表达量,倪力刚等[30]也得到一致结果。Jeong等[31]用OC/OB共培养物来考察骨碎补的有效成分在骨吸收中的作用,在实验中分别用磷脂酰肌醇3(PI3)激酶抑制药渥曼青霉素(wT)和蛋白抑制药、蛋白激酶抑制药(calphostin C)为对照考察骨碎补有效成分对OC介导CTSK在OC中的成熟与转运作用,发现骨碎补与wT作用类似,在骨吸收过程中抑制CTSK前体的胞内转运与表达。
2.3 Wnt/β-catenin 信号通路
Wnt 信号通路参与人体内多种器官和组织的生长、发育和分化调控,其在OP方面的研究目前已受到广泛的关注。目前已知的Wnt 细胞内信号通路传导途径有多条,其中 Wnt/β-catenin 通路最为经典,在骨吸收和骨形成过程中起重要作用[32]。Wnt/β-catenin通路作用机制为:Wnt蛋白与细胞膜受体卷曲蛋白(Fz)和辅助受体脂蛋白受体相关蛋白 5/6(LRP5/6)结合,活化Fz受体,通过骨架蛋白(Dv)和酪蛋白激酶1(CK1)传递信号,活化由轴蛋白(axin)、结肠腺瘤性息肉病蛋白(APC)和糖原合成酶激酶3(GSK3)组成的复合物,激活细胞内信号通路[33]。无Wnt信号刺激时,GSK 3 磷酸化β-catenin,β-catenin 通过泛素/蛋白酶体途径降解。而当有 Wnt 信号刺激时,蛋白复合物解离不发生磷酸化,catenin 在胞浆中积聚并转位入核,与转录因子T细胞因子/淋巴细胞增强因子(TCF/LEF)结合启动靶基因如 c-myc、cyclin D1的转录,促进细胞的增殖或活化,该过程对成骨非常关键[34]。
郭英等[35]探讨骨碎补总黄酮对BMSC成骨分化过程中对Wnt/β-catenin信号通路相关因子mRNA表达的影响发现,骨碎补总黄酮在分化早期促进β-catenin、淋巴增强因子1(LEF-1)、细胞周期蛋白D (cycline D) mRNA的表达,在晚期抑制其表达,认为在骨碎补总黄酮诱导BMSC成骨分化过程中,Wnt/β-catenin信号通路可能主要参与早期分化。Ma等[36]报道,柚皮苷可对抗切除坐骨神经所诱发的废用性骨质疏松,促进OB分化并抑制OC的分化,在活化 Wnt/β-catenin 信号通路的同时也增加Sema3A的表达。Lv等[37]也发现柚皮苷可通过活化Wnt/β-catenin 信号通路,增加骨膜蛋白的表达并抑制壳硬蛋白的表达从而阻止废用性骨质疏松的进程。Wang等[38]报道柚皮苷可对抗去卵巢性骨质疏松,增强骨强度,增加β-catenin的mRNA及蛋白的表达,并活化LEF、TCF转录因子。

图3 骨碎补有效成分调控Wnt/β-catenin 信号通路Fig.3 Effective constituent of rhizoma drynariae regulating the Wnt/β-catenin signaling pathway
2.4 BMP信号通路
BMPs属于转移生长因子TGF-β超家族中的一员,其主要生物学作用是诱导未分化的间充质细胞增殖及发生成骨分化,最终促进软骨和新生骨的形成。BMP信号通路主要是通过BMPs与其活化素激酶受体相结合,激活细胞内下游通路如Smad和MAPK等信号通路,从而发生一系列磷酸化或聚合酶链反应[39]。BMP作为调节OB生长最重要的系列生长因子之一,对骨形态的发生具有决定性作用。目前的研究[40]结果发现,BMP 具有很强的促进OB分化和诱导体外成骨的能力,在诱导人骨髓间充质干细胞发生成骨分化的同时还可抑制其向脂肪细胞和肌肉细胞转化。因此,通过促进骨组织中BMPs的表达来增加骨量可以达到改善OP的目的。其中BMP-2 是诱导成骨活性最强的BMPs之一,主要对未分化间充质细胞和骨系细胞起到募集和分化作用[41]。
张迪华等[42]通过摘除性成熟SD大鼠双侧卵巢,模拟绝经后期大鼠,给予强骨胶囊(主要含骨碎补总黄酮)进行干预,观察在雌激素撤退的情况下,BGP和BMP-2在骨组织中的表达情况,研究发现去卵巢大鼠口服强骨胶囊能促进BMP-2和BGP的表达。董福慧等[43]研究发现,在骨愈合过程中,骨碎补对TGF-β1mRNA、BMP-2 mRNA基因表达具有有益的调节作用,说明成骨可能与骨碎补上调BMP信号通路中TGF-β1mRNA、BMP-2 mRNA基因表达有关。而Yang 等[44]也发现,骨碎补水提液能有效的促进SD大鼠MSCs增殖和骨向分化,并且在MSCs骨向分化过程中,骨碎补水提液可上调TGF-β1、BMP-2的表达量。Wu等[45]报道,柚皮苷具有预防卵巢切除小鼠骨量丢失的潜能和作用,其作用机制是柚皮苷在OB中通过磷脂酰肌醇3激酶/蛋白激酶B通路,转录因子c-Fos/c-Jun和活化蛋白-1旁路诱导BMP-2表达。
上述提到,BMP信号通路的下游存在Smad信号通路和MAPK信号通路两条重要的通路,其对成骨的作用非常关键。朱慧锋等[46]通过手术切SD大鼠的双侧卵巢,建立绝经后骨质疏松模型,研究骨碎补总黄酮对骨质疏松大鼠Smad1、Smad5 mRNA 表达的影响,发现骨碎补总黄酮能够上调骨髓微环境中Smad1、Smad5 mRNA的表达水平,并以中剂量组最明显。汪甜等[47]研究表明,柚皮苷具有显著促进MSCs 骨向分化的能力,主要是通过激活MAPK信号通路中ERK通路、JNK 通路,以及上调 BMP-2的表达来促进 MSCs 骨向分化的,其中柚皮苷上调BMP-2的表达受MAPK通路中p38通路的影响较大。

图4 骨碎补有效成分调控BMP信号通路Fig.4 Effective constituent of rhizoma drynariae regulating the BMP signaling pathway
3 结语
寻找OP相关分子信号通路上的作用靶点并进行相关的靶向治疗,是有效防治OP的重要途径。在OP的临床治疗中,中药发挥整体调节和综合治疗的优势,能明显改善患者骨密度[48]。近些年来,骨碎补在防治OP方面取得了较为确切的临床疗效,而大量文献也报道骨碎补能促进骨形成与抑制骨吸收,同时发现相关信号通路在骨碎补有效成分抗OP过程中发挥重要调控作用。因此,深入探讨骨碎补活性单体成分的多通路分子生物学机制,对指导骨碎补抗OP治疗具有重要意义。
但目前骨碎补作用于骨代谢分子信号通路的研究尚处于起步阶段,就其研究广度而言,除了上述所报道的分子信号通路以外,还包括PTH通路、NF-κB通路、雌激素通路、谷氨酸通路等[49];就其研究深度而言,大部分分子机制研究仅限于单一通路,而骨代谢受到多条信号通路的调节,这些通路之间交叉重叠、相互联系,共同构成了一个影响骨稳态的网络信号调控系统。另外,由于传统分子生物学技术对单个或数个分子信号基因或蛋白检测的局限,以及基因芯片等高通量筛选技术的应用难度,导致研究效率不高,因而难以全面诠释骨碎补多靶点、多途径、多层次的作用机制,这些问题有待研究者们进一步的努力探索和解决,而通路间的相互交联(crosstalk)也将成为今后的研究重点之一。
其次,由于目前在研究分子信号通路的相关调控蛋白时,使用的干预药物主要是骨碎补总黄酮,骨碎补总黄酮作为一种混合物,难以明确其多个单体成分与信号通路调控靶点之间的关系,因此有效单体成分的分离、鉴定及相关体内、体外研究仍需广泛开展。
