色季拉山冬虫夏草适生地草甸地表节肢动物群落多样性
2017-08-02张剑霜吴文静张古忍2
喻 浩,张剑霜,吴文静,张古忍2,*
色季拉山冬虫夏草适生地草甸地表节肢动物群落多样性
喻 浩1,2,张剑霜3,吴文静2,4,张古忍2,3*
(1.贵州师范学院,化学与生命科学院,山地研究所,贵阳 550018;2.中山大学有害生物控制与资源利用国家重点实验室,广州 510275;3.中山大学食品与健康工程研究院,广州 510275;4.广东省生物资源应用研究所,广东省农业害虫综合治理重点实验室,广州 510275)
色季拉山;冬虫夏草;高寒草甸;地表节肢动物;多样性
冬虫夏草是我国传统名贵中药材,主要分布于海拔4500 m以上的青藏高原高寒草甸。复杂的气候条件,加之近年来过度放牧和无序采挖等人为干扰的介入,使得青藏高原地区的环境脆弱多变,进而会对冬虫夏草的发生产生影响(张古忍等,2011)。无疑,掌握高寒草甸的的生态条件和生物多样性水平,是冬虫夏草资源的可持续利用研究的需要(邹志文,2009;喻浩,2013)。
以昆虫为代表的节肢动物是草地生物多样性的重要组成部分,它们种类众多,数量巨大,分布广泛,与脊椎动物相比,节肢动物在自然界中占据了多样性更高空间尺度更小的生境,对生境的变化更敏感,可作为环境的指示种发挥重要的作用(蔡海等,2012)。虫草寄主钩蝠蛾需要在土壤中度过约3-4年的漫长幼虫期,期间被真菌中国被毛孢侵染即可能形成冬虫夏草(朱印酒等,2009;李峻锋等,2011),因此土壤表层区域的节肢动物可作为生态探针,用以反映冬虫夏草的微生境特征。基于以上理由,本文选取了位于西藏色季拉山的3类典型冬虫夏草适生地草甸,研究并比较其地表节肢动物群落及多样性特征,以期利用节肢动物的的生态指示及调控功能,探寻高寒草甸生态系统环境异质化的原因,为冬虫夏草适生地生物多样性水平的维持与保护提供科学建议及决策依据。
1 材料与方法
1.1 样地设计
根据色季拉山的地形地貌、生境特点和管理状况,将冬虫夏草适生草甸划分为3种生境类型:山坡草甸、林缘草甸与滩地草甸(表1),每种生境设计8个样点作为重复,每样点设置4口地面陷阱,采集范围约5 m×5 m,样点之间直线距离大于50 m,样点与生境边缘距离大于100 m,以消除边缘效应。

表1 色季拉山3种冬虫夏草适生草甸自然环境概况Table 1 Nature environment of different meadow types
1.2 调查方法
采用由两个一次性塑料杯相嵌埋入土壤作为陷阱。塑料杯使用一般市售的300 mL杯,口径7.2 cm,深度10.5 cm。陷阱溶剂具体配方为每1000 mL 4%福尔马林液加5 mL甘油和几滴洗衣粉溶液。杯口与地面平齐,为减少陷阱溶液的蒸发以及防止降雨和大的杂物落入,每个陷阱上都有铁丝支撑的塑料碗作棚。
采集的时间覆盖色季拉山的无雪季节,具体为2009年6月-2009年11月,2010年4月-2010年11月,每10天收集一次陷阱中的标本并更换陷阱溶液,标本先置于75%的酒精中进行固定,1天后换成80%的干净酒精保藏待检。共计收集34个批次。
标本鉴定的分类阶元级别为科,并区别开形态种。鉴定主要依据《昆虫分类》(郑乐怡和归鸿,1999)、《昆虫形态分类学》(忻介六等,1985)和《中国动物志·昆虫纲》(中国科学院中国动物志编辑委员会,2003)。标本鉴定由中山大学昆虫分类学研究室及北京中国科学院动物学研究所协助完成。
1.3 数据分析
抽样充分性:利用EstimateS8.2.0软件计算ACE(abundance-base coverage estimator)估计值,并通过Excel进行物种累积曲线(species accumulation curves)的绘制。根据曲线的特征,并结合物种丰富度(S)(物种数实测值)与ACE估计值的比率进行抽样充分性判断(李巧等,2009)。
相对多度模型:以“物种-多度曲线”(species-abundance curve)表现种的相对多度分布,横坐标为种数的自然分组,以对数倍程(个体数量取以2为底的对数)对个体数量进行统计,倍程1、2、3、4等分别对应个体数为1、2-3、4-7、8-15等等,纵坐标是每个倍程中的物种数。使用Preston对数正态分布模型拟合各生境相对多度,检验其分布是否符合对数正态分布,模型公式:S(R)=S0exp (-α2R2),式中S(R)指第R倍程中物种数,S0为众数倍程物种数,α是与分布有关的参数(May; 1975)。
各生境多样性指数及比较:分别计算每个样点的Margalef丰富度指数dma、Shannon-Wiener多样性指数H′、Pielou均匀度指数J、Simpson优势度指数λ。计算公式:dma=(S-1)/ lnN;λ=∑Pi2;H′=-∑PilnPi;J=H′/H′max。上述各式中,S为物种数,N为个体总数,Pi第i种占群落中个体总数N的比例,各多样性指数计算使用Past软件。由于每种生境的样点量有限且不服从方差岐性,使用非参数的克罗斯考尔和瓦里斯秩方差分析(Kruskal-Wallis ANOVA by Ranks),及平均等级排序多重比较分析,比较3种草甸类型上述各指数的差异性,非参数多重比较使用Statistica 10.0软件完成。
2 结果与分析
2.1 物种丰富度及抽样充分性估计
基于3种草甸类型地表节肢动物采样批次和个体数量,物种数量的物种积累曲线都接近水平渐近线(图1),说明继续增加取样数量物种数量增加的速率将会很低(León-Cortésetal., 1998),物种丰富度由高到低分别为林缘草甸、山坡草甸、滩地草甸,3类生境的物种数量都稳定在90-100种(图1A,表2)。滩地草甸比其他两种生境得到更多的个体,却属于更少的物种(图1B,表2),表明其节肢动物群落高度的不均匀,可能有少部分物种在此生境中占有极大优势。
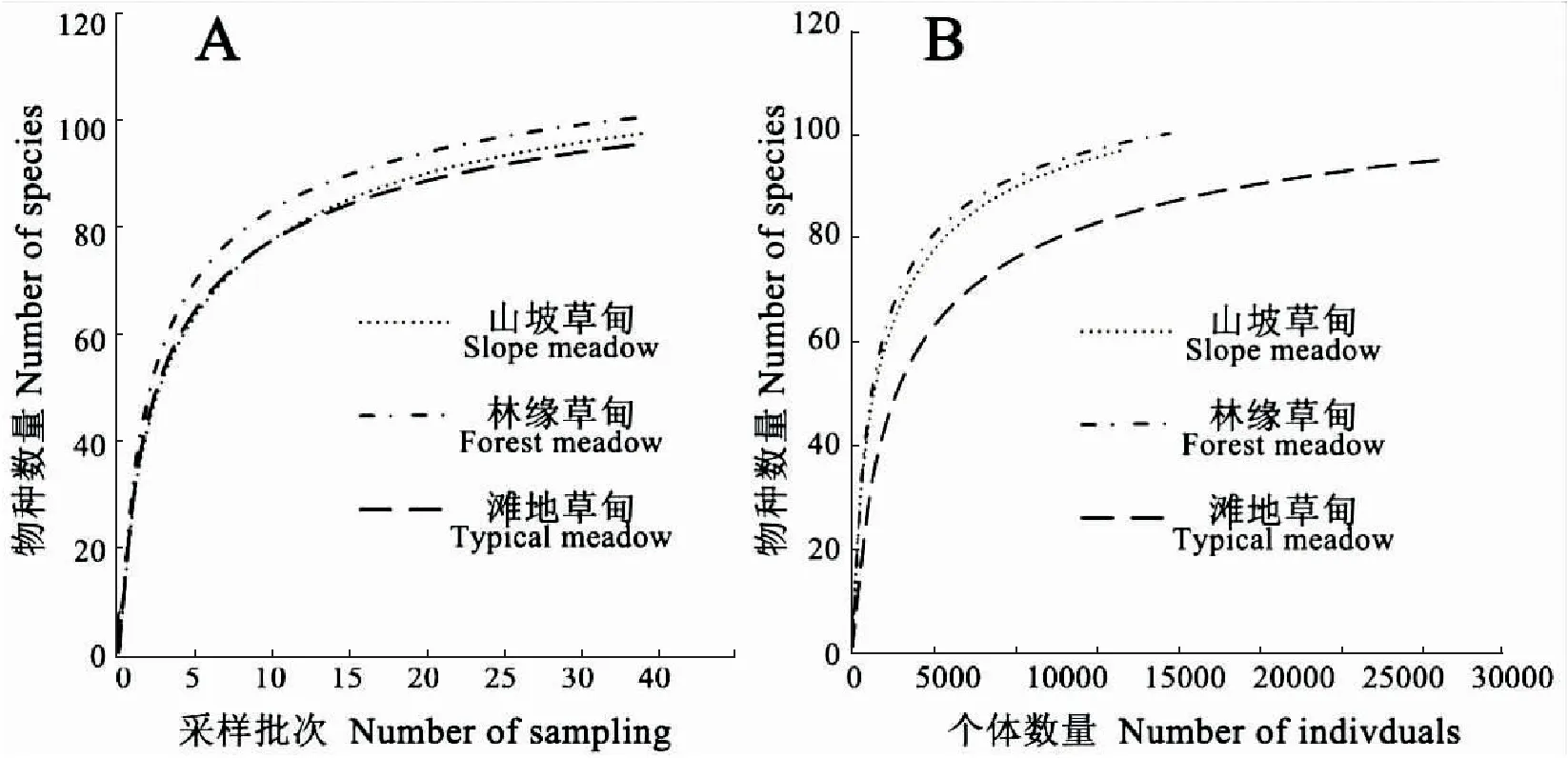
图1 色季拉山3种冬虫夏草生境草甸地表节肢动物的物种积累曲线Fig.1 Species-accumulation curves of ground-dwelling arthropods in three habitats types注:A,基于采样批次的物种累积曲线;B,基于个体数量的物种累积曲线。Note: A, Horizontal ordinate: Number of sampling; B, Horizontal ordinate: Number of individuals.

表2 色季拉山3种冬虫夏草生境地表节肢动物的物种实际采集量、种积丰富估值和抽样率Table 2 Species richness, ACE, ratio of S to ACE of ground-dwelling arthropods in three habitats types
从抽样效果来看,3种生境实际采集到的物种数目都超过全部物种(物种丰富度估计值)的90%以上(表2),采集几乎覆盖生境中的全部物种,效果较好。数据可进行后续分分析。
2.2 种类组成与数量分布
2.2.1 色季拉山冬虫夏草适生草甸的地表节肢动物群落组成
调查一共获得54757号可明确进行鉴定的标本,隶属14目、62科、114种。在科级水平上,双翅目昆虫的类群最为丰富,占全部类群24.19%,其次为鞘翅目、蜘蛛目和膜翅目。在物种水平,各目的种类比例与科级水平相似,但物种最多的为鞘翅目,占全部类群22.61%,其次为双翅目、膜翅目和蜘蛛目。襀翅目、革翅目和蜈蚣目的类群较为单一,均只统计到1科1种。个体数量最多的是弹尾目,占总个体数量的28.39%,其次为鞘翅目、双翅目、膜翅目等(表3)。
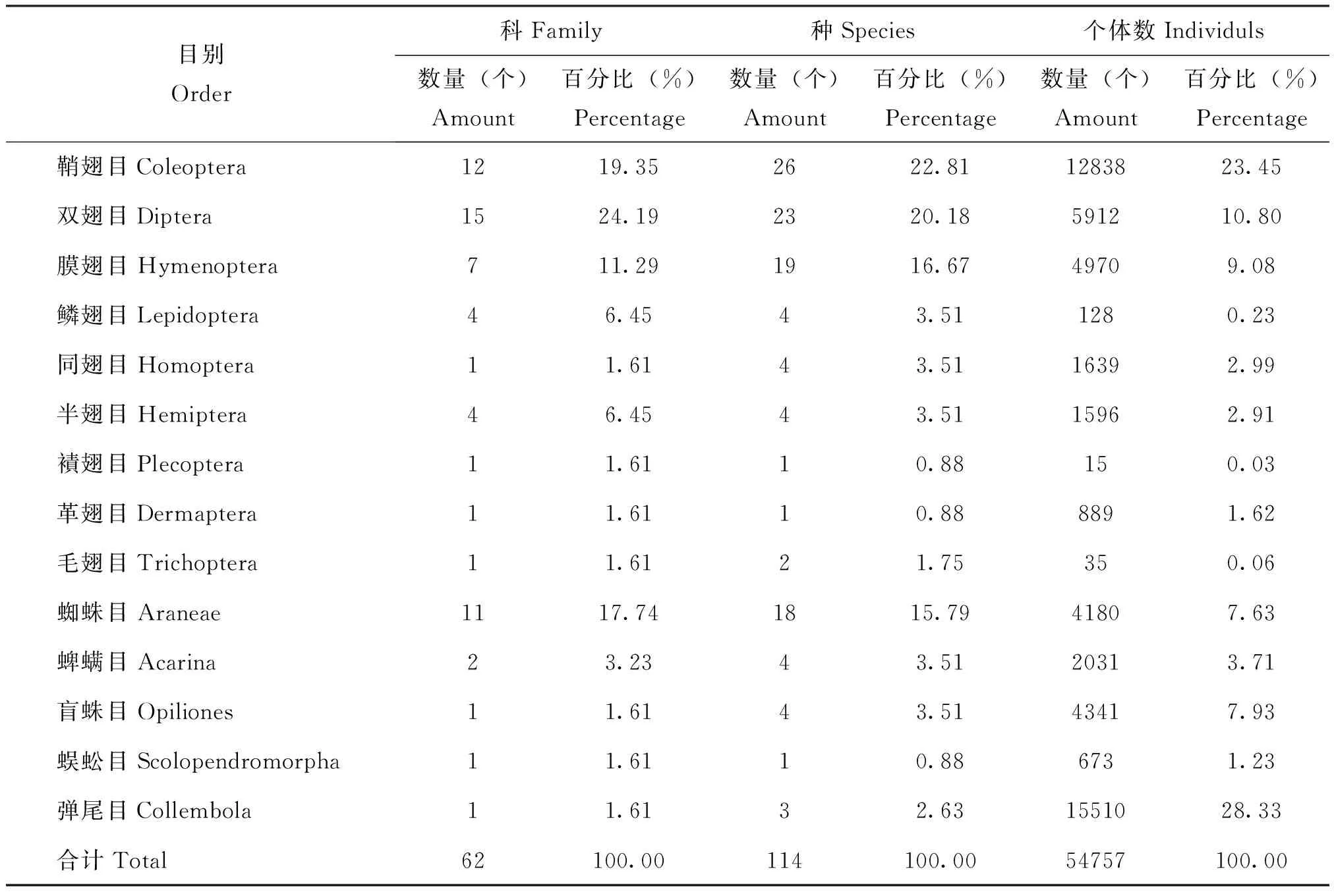
表3 色季拉山冬虫夏草生境地表节肢动物的种类组成Table 3 Ground-dwelling arthropods communities in habitats of Cordyceps sinensis in Mt.Segrila
2.2.2 不同生境类型区域地表节肢动物群落的数量特征
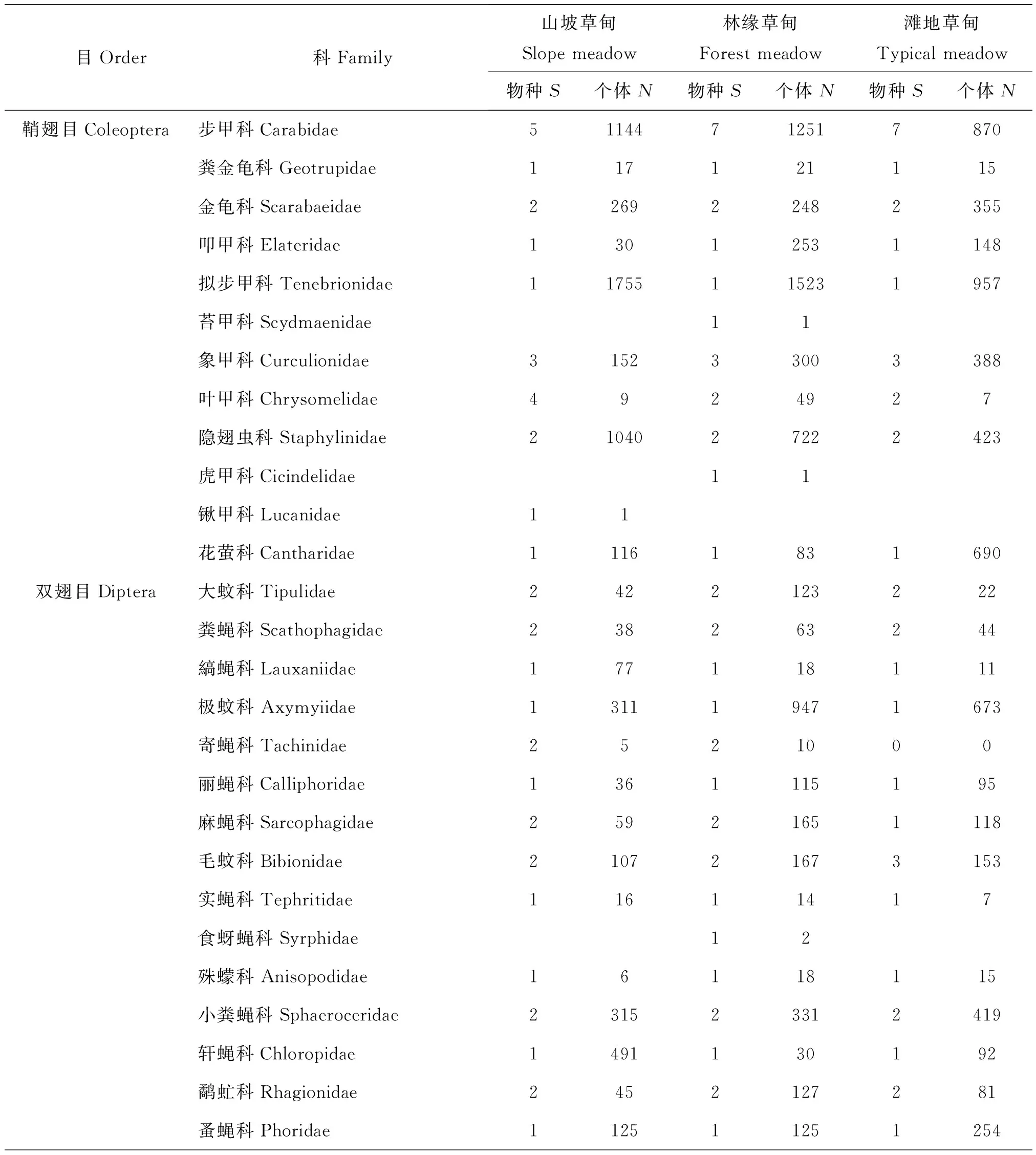
表4 色季拉山冬虫夏草不同生境类型的地表节肢动物的群落组成Table 4 Ground-dwelling arthropods communities in three habitats of Cordyceps sinensis in Mt.Segrila
续上表
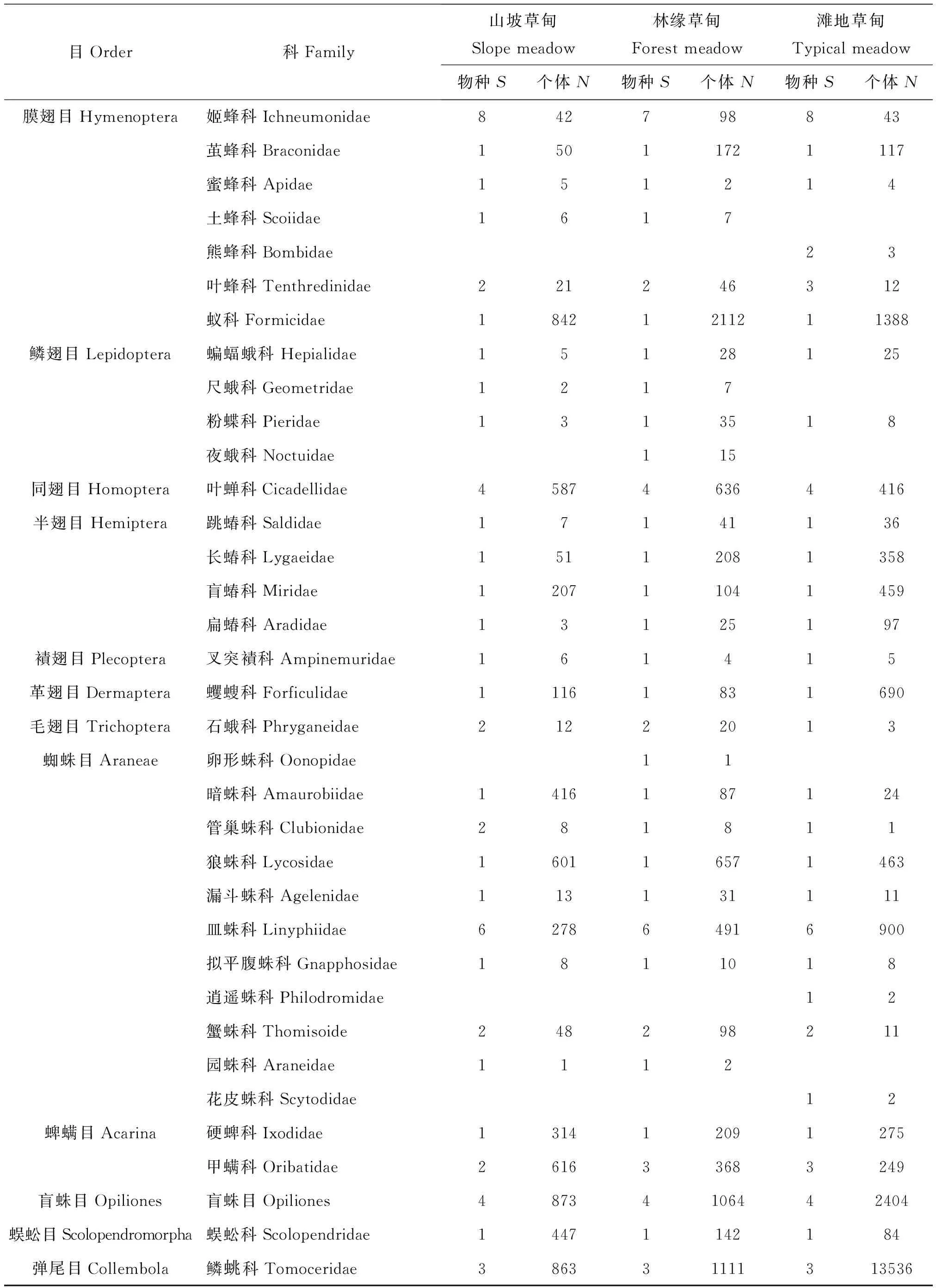
目Order科Family山坡草甸Slopemeadow林缘草甸Forestmeadow滩地草甸Typicalmeadow物种S个体N物种S个体N物种S个体N膜翅目Hymenoptera姬蜂科Ichneumonidae842798843茧蜂科Braconidae15011721117蜜蜂科Apidae151214土蜂科Scoiidae1617熊蜂科Bombidae23叶蜂科Tenthredinidae221246312蚁科Formicidae18421211211388鳞翅目Lepidoptera蝙蝠蛾科Hepialidae15128125尺蛾科Geometridae1217粉蝶科Pieridae1313518夜蛾科Noctuidae115同翅目Homoptera叶蝉科Cicadellidae458746364416半翅目Hemiptera跳蝽科Saldidae17141136长蝽科Lygaeidae15112081358盲蝽科Miridae120711041459扁蝽科Aradidae13125197襀翅目Plecoptera叉突襀科Ampinemuridae161415革翅目Dermaptera蠼螋科Forficulidae11161831690毛翅目Trichoptera石蛾科Phryganeidae21222013蜘蛛目Araneae卵形蛛科Oonopidae11暗蛛科Amaurobiidae1416187124管巢蛛科Clubionidae281811狼蛛科Lycosidae160116571463漏斗蛛科Agelenidae113131111皿蛛科Linyphiidae627864916900拟平腹蛛科Gnapphosidae1811018逍遥蛛科Philodromidae12蟹蛛科Thomisoide248298211园蛛科Araneidae1112花皮蛛科Scytodidae12蜱螨目Acarina硬蜱科Ixodidae131412091275甲螨科Oribatidae261633683249盲蛛目Opiliones盲蛛目Opiliones48734106442404蜈蚣目Scolopendromorpha蜈蚣科Scolopendridae14471142184弹尾目Collembola鳞科Tomoceridae386331111313536
2.2.3 不同生境地表节肢动物群落的物种-多度分布
分别对3种生境中的地表节肢动物多度进行了对数正态分布模型拟合。结果表明:林缘草甸的群落表现为典型的对数正态分布(P=0.81),山坡草甸和滩地草甸不符合对数正态模型(P<0.05),更接近对数级数分布。上述数据分析结果与其各自种-多度分布图相符(图2)。
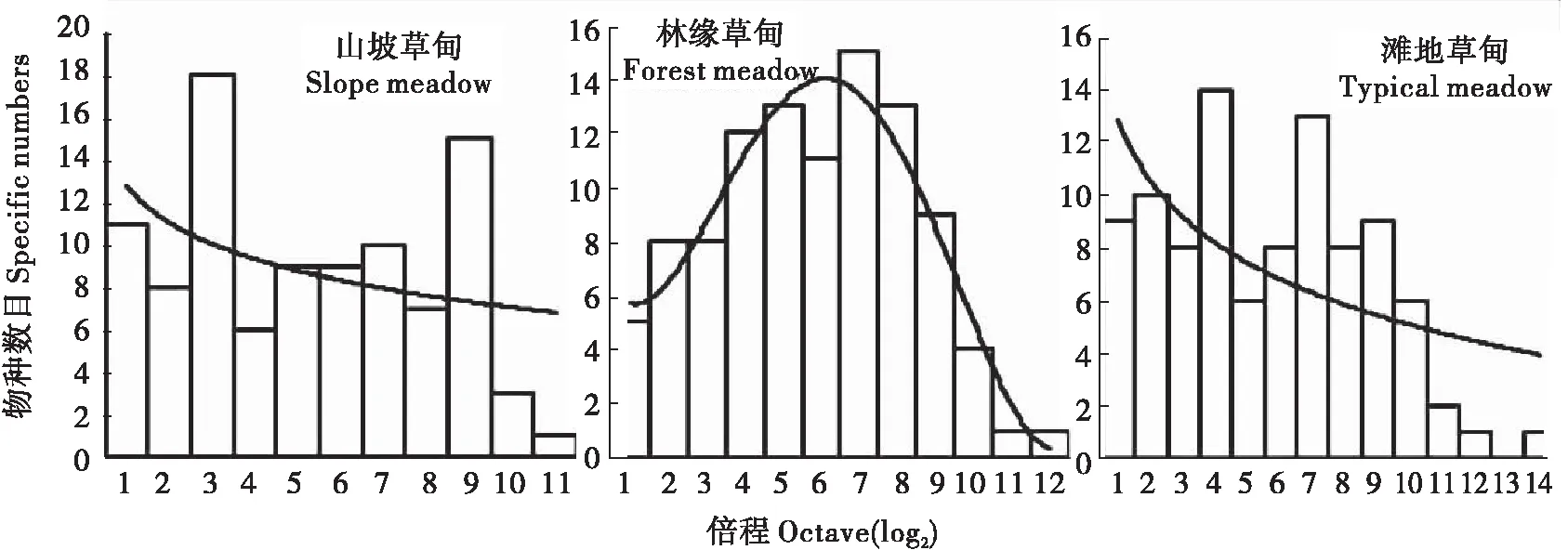
图2 色季拉山冬虫夏草不同生境中地表节肢动物群落种—多度曲线Fig.2 Species-abundance curves forground-dwelling arthropods in three habitats types
由图2可知,林缘草甸的中间物种较多,优势种和富积种较少;山坡草甸出现了两个明显的波峰,分别位于倍程轴的左右两端,可见其整体分布格局变化剧烈,稀疏种和富积种均占据了较大比例,但除此之外,其余倍程波动平缓,说明群落的数量分布较均匀;滩地草甸中,不但更多的类群集中在倍程轴的左端,而且在13倍程处出现了断点,说明稀疏物种在地表节肢动物群落中占据了较大比例,而少量富积种在个体数量上占有极大优势,群落结构极不稳定。
2.3 不同生境类型区域地表节肢动物多样性特征
由图3可知,地表节肢动物多样性丰富度指数、均匀度指数和多样性指数表现完全一致,均为在林缘草甸中最高,山坡草甸次之,滩地草甸最低;而滩地草甸的λ优势度指数明显较高,山坡草甸和林缘草甸较为接近但后者略高。
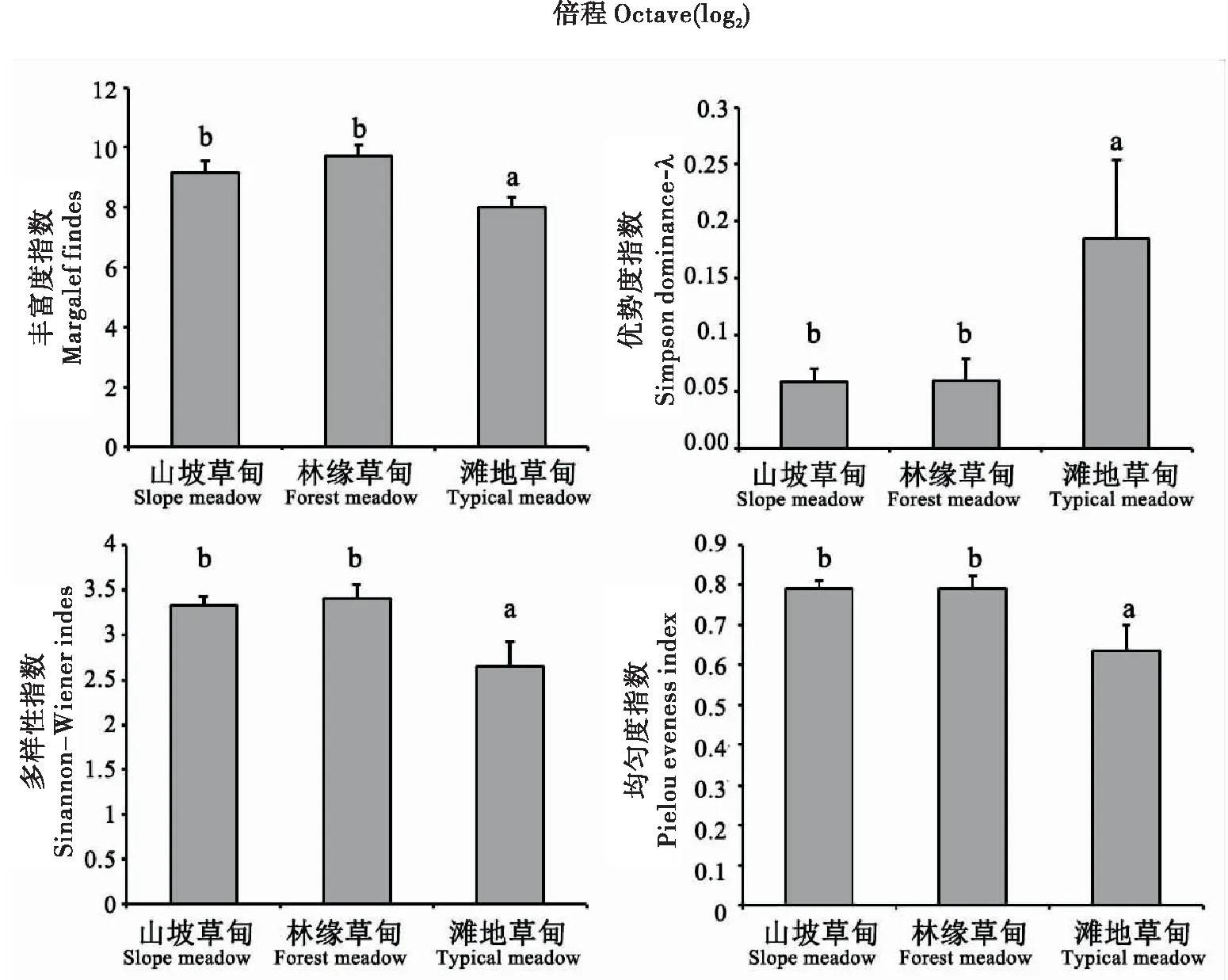
图3 色季拉山冬虫夏草不同生境草甸中地表节肢动物群落多样性特征Fig.3 Diversity indexes ofground-dwelling arthropods in three habitats types in Mt.Segrila注:柱形图上部的工字线表示标准差,标准差线上部的不同字母表示差异显著(P<0.05, Kruskal-Wallis test)。Note: Bars denote standard deviation; different letters above bars denote significant differences among means (P<0.05, Kruskal-Wallis test 5).
以生境为分组变量进行克罗斯考尔和瓦里斯秩方差分析,结果显示,不同生境下地表节肢动物丰富度(H2,24=17.4150,P=0.0002)、λ优势度(H2,24=14.4950,P=0.0007)、多样性(H2,24=16.3400,P=0.0003)和均匀度指数(H2,24=15.0950,P=0.0005)均差异显著。平均等级排序多重比较结果表明,四个多样性指数在山坡草甸和林缘草甸二者之间差异均不显著,在滩地草甸与上述两种生境之间差异显著。
3 结论与讨论
3.1 物种丰富度及抽样充分性
调查共计获得地表节肢动物标本54757号,经鉴定,隶属14目、62科、114种。通过物种累积曲线和种积丰度估值的直观反映,在3种生境中均表现出极好的抽样效果,山坡草甸、林缘草甸和滩地草甸的物种数量分别为97种、100种和95种。目前国内大部分研究直接以样本的物种数目S值来表现群落的物种丰欠状况,可能导致对实际物种数的过低估计(李巧等,2009),仅有少数学者尝试在多样性研究中运用物种累积曲线和种积丰度估值(郑国等,2009;陈又清等,2010),然而从他们的分析结果来看,抽样效果并不理想(李巧,2009)。
与上述研究相比,本研究查获的物种数量较少,采样效果却更为理想。抽样的充分性及物种数量较少的客观事实产生相悖,原因可能有如下几点:(1)高寒地区的群落的相对单一性,即物种的多样性随着海拔升高和温度降低有下降趋势,这也与诸多研究的结果吻合(Gaston and Chown, 1999;Addo-Bediakoetal., 2000;Sandersetal., 2007;Yangetal., 2013);(2)本研究的调查批次达到34次,跨越2个年度,在采样频率和时间跨度上都超过许多同类研究(Jungetal., 2008;Hsieh and Linsenmair, 2012),然而样地面积及样点数目却都少于上述研究,小范围内的长时间采集,对调查地点充分抽样,但是同时限制了物种数量。扩大抽样规模、使用种-面积累计曲线考察、对更多的异质性的样地进行采集,调查的结果可能更能反映高寒地区的节肢动物多样性,但是考虑到本研究仅针对冬虫夏草适生地,这样的抽样结果是完全可以接受的。
3.2 不同冬虫夏草生境的地表节肢动物种类组成和数量结构
3.3 不同冬虫夏草生境的地表节肢动物的物种-多度分布格局
物种-多度曲线反映了物种的数量分布特征。而本研究的结果显示:林缘草甸的地表节肢动物群落能较好的拟合对数正态分布模型,符合一般的昆虫群落的模式,表明群落生境的条件较好、物种丰富而分布又较为均匀;而山坡草甸和滩地草甸的分布格局则更接近对数级数分布,符合生态位优先占领假说。由于对数级数分布多出现在生境条件较为严酷、物种少而生境又不相互重叠的群落(郑国,2008),由此可见,山坡草甸和滩地草甸生态系统生境条件相对较差,这可能是放牧、砍伐行为对本身就十分脆弱的高寒生境造成的干扰所致。而在基本无管理的林缘草甸,植被更易到达稳定而健康的状态,因此较之另外两种生境,其物种数量分布更为均匀,优势类群的比例较低,这与Koivula(2002)及Niemela(2001)等人关于阔叶林中大型节肢动物物种-多度曲线随着植物群落多样性变化,植被群落越稳定,动物种-多度曲线越平缓、物种的数量分布更均匀的结论一致。
3.4 不同冬虫夏草生境的地表节肢动物的多样性比较
多样性、优势度和均匀性这三者在以往的研究中通常会同时使用。多样性较高的生境往往有更低的优势度,群落分布也更均匀(Jungetal., 2008; Hsieh and Linsenmair, 2012),本研究的结果也支持了这一点:滩地草甸的多样性最低,优势类群的优势度最高,群落最不均匀,但其他两种生境在各类多样性指数上无差异。关于多样性的研究大部分会选取干扰程度不同的几种生境加以比较,并且结果基本上都能支持中度干扰假说(Intermediate disturbance hypothesis,IDH)(侯沁文等,2015;杨立军和张丹丹,2015)。但由于本研究开展地点位于高寒地区,几种生境都基本处于无干扰状态,如果将放牧行为也视为一种干扰,那么如前所述,阴坡应该最接近自然状态的生境,但同时却表现出最高的多样性(虽然略高于阳坡却无显著差异),该结果与IDH并不完全相符,原因之一可能是高寒地区群落的抵抗干扰的能力远远低于温带及热带地区,即便是放牧这种程度的干扰也会引起多样性的降低。高寒草甸的过度放牧应引起注意。
综上所述,通过野外调查和后续的数据分析,可初步了解冬虫夏草适生地草甸节肢动物群落的物种多样性概况,以其作为生态指示探针,可在一定程度上反映冬虫夏草适生地的生态条件。总体来说,冬虫夏草微生境表现出简单、群落结构较为单一的特征,且易受到热量条件、人为干扰等因素的影响。控制温度条件和人为管理程度,有利于冬虫夏草生境中的生物多样性的维持,而地表节肢动物多样性水平和冬虫夏草产量、虫草寄主钩蝠蛾昆虫的种群密度之间的关系,将是下一步非常值得研究的问题。
References)
Addo-Bediako A, Chown SL, Gaston KJ.Thermal tolerance, climatic variability and latitude [J].ProceedingsoftheRoyalSocietyofLondon.SeriesB:BiologicalSciences, 2000, 267 (1445): 739-745.
Chen YQ, Li Q, Wang SM,etal.Diversity of Curculionoidea in subtropical monsoon evergreen broadleaved forest in Puer City, Yunnan [J].JournalofYunnanUniversity, 2010, 32 (1): 97-102, 107.[陈又清, 李巧, 王思铭, 等.普洱市亚热带季风常绿阔叶林区象甲总科昆虫多样性[J].云南大学学报, 2010, 32 (1): 97-102, 107]
Editorial Board of Fauna Sinica, the Chinese Academy of Sciences.Fauna Sinica.Insecta [M].Beijing: Science Press, 2003.[中国科学院中国动物志编辑委员会.中国动物志.昆虫纲[M].北京: 科学出版社, 2003]
Gaston KJ, Chown SL.Elevation and climatic tolerance: A test using dung beetles [J].Oikos, 1999, 86 (3): 584-590.
Hou QW, Tie J, Bai HY.Effects of human disturbance on moth diversity in Manghe National Nature Reserves, Shanxi, Northern China [J].JournalofEnvironmentalEntomology, 2015, 37 (1): 20-29.[候沁文, 铁军, 白海艳等.人为干扰对山西蟒河国家级自然保护区蛾类多样性的影响[J].环境昆虫学报, 2015, 37 (1): 20-29]
Hill T, Lewicki P.Statistics:Methods and applications [Z].Tulsa: StatSoft, 2007.
Hsieh YL, Linsenmair KE.Seasonal dynamics of arboreal spider diversity in a temperate forest [J].EcologyEvolution, 2012, 2 (4): 768-777.
Jung M, Kim S, Kim H,etal.Biodiversity and community structure of ground-dwelling spiders in four different field margin types of agricultural landscapes in Korea [J].AppliedSoilEcology, 2008, 38 (2): 185-195.
Koivula M, Kukkonen J, Niemelă J.Boreal Carabid-Beetle (Coleoptera, Carabidae) Assemblages along the clear-cut originated succession gradient [J].BiodiversityandConservation, 2002, 11 (7): 1269-1288.
León-Cortés JL, Soberón-Mainero J, Llorente-Bousquets J.Assessing completeness of Mexican sphinx moth inventories through species accumulation functions [J].DiversityandDistributions, 1998, 4: 37-44.
Li JF, Zou ZW, Liu X,etal.Biology ofThitarodespui(Lepidoptera, Hepialidae), a host species ofOphiocordycepssinensis [J].JournalofEnvironmentalEntomology, 2011, 33 (2): 195-202.[李峻锋, 邹志文, 刘昕等.冬虫夏草寄主蒲氏钩蝠蛾的生物学[J].环境昆虫学报, 2011, 33 (2): 195-202]
Li Q.Species accumulation curves and its application [J].ChineseJournalofAppliedEntomology, 2009, 48 (6): 1882-1888.[李巧.物种累积曲线及其应用[J].应用昆虫学报, 2009, 48 (6): 1882-1888]
Li Q, Chen YQ, Chen ZH.Research methods on ant community [J].ChineseJournalofEcology, 2009, 28 (9): 1862-1870.[李巧, 陈又清,徐正会.蚂蚁群落研究方法[J].生态学杂志, 2009, 28 (9): 1862-1870]
May RM.Patterns of Species Abundance and Diversity [M].Cambridge: Harvard University Press, 1975, 81-120.
Sanders NJ, Lessard J, Fitzpatrick MC,etal.Temperature, but not productivity or geometry, predicts elevational diversity gradients in ants across spatial grains [J].GlobalEcologyandBiogeography, 2007, 16 (5): 640-649.
Xin JL, Yang QS, Hu CY.InsectMorphological Taxonomy [M].Shanghai: Fudan University Press, 1985.[忻介六, 杨庆爽, 胡成业.昆虫形态分类学[M].上海: 复旦大学出版社, 1985]
Yang LJ, Zhang DD.Diversity of moth communities and variation along artificial disturbance gradient in Mount Jinggangshan National Nature Reserve, China [J].JournalofEnvironmentalEntomology, 2014, 36 (5): 679-686.[杨立军, 张丹丹.井冈山自然保护区蛾类多样性及人为干扰的影响[J].环境昆虫学报, 2014, 36 (5): 679-686]
Yang ZL, Guo H, Zhang J,etal.Stochastic and deterministic processes together determine alpine meadow plant community composition on the Tibetan Plateau [J].CommunityEcology, 2013, 171 (2): 495-504.
Yu H.Population Differentiation ofThitarodesand Ground-dwelling Arthropods Community in the Habitat ofThitarodesin Mt.Sejila of Tibet [D].Guang zhou: Sun Yat-Sen University, 2013.[喻浩.西藏钩蝠蛾种群分化及其适生地地表节肢动物多样性研究[D].广州: 中山大学, 2013]
Zhang GR, Yu JF, Wu GG,etal.Factors influencing the occurrence ofOphiocordycepssinensis[J].ActaEcologicaSinica, 2011, 31 (14): 4117-4125.[张古忍, 余俊锋, 吴光国等.冬虫夏草发生的影响因子[J].生态学报, 2011, 31 (14): 4117-4125]
Zheng G.Spider Diversity in Xishuangbanna Rainforest, Southwest of China [D].Beijing: Institute of Zoology, Chinese Academy of Sciences, 2008.[郑国.西双版纳热带雨林蜘蛛多样性研究[D].北京: 中国科学院动物学研究所, 2008]
Zheng G, Yang XD, Li SQ.Biodiversity of ground-dwelling spider in six forest types in Xishuangbanna, S.W.China [J].ActaEntomologicaSinica, 2009, 52 (8): 875-884.[郑国, 杨效东, 李枢强.西双版纳六种林型地表蜘蛛多样性比较研究[J].昆虫学报, 2009, 52 (8): 875-884]
Zheng LY, Gui H.Insect Classification [M].Nanjing: Nanjing Normal University Press, 1999 [郑乐怡, 归鸿.昆虫分类[M].南京:南京师范大学出版社, 1999]
Zhu YJ, Duan SQ, Ouzhu LJ.Advances in the study ofCordycepssinensis[J].JournalofMinzuUniversityofChina(Natural Sciences Edition), 2009, 18 (2): 27-33.[朱印酒, 段双全, 欧珠朗杰.冬虫夏草的研究进展[J].中央民族大学学报(自然科学版), 2009, (2): 27-33]
Zou ZW.On the Insects of the GenusThitarodesin Mt.Sejila of Tibet [D].Guangzhou: Sun Yat-sen University, 2009.[邹志文.西藏色季拉山钩蝠蛾属Thitarodes昆虫研究[D].广州: 中山大学, 2009]
Ground-dwelling arthropods communies in different habitat ofCordycepssinensisin Mt.Segrila of Tibet
YU Hao1,2, ZHANG Jian-Shuang3, WU Wen-Jing2,4, ZHANG Gu-Ren2,3*
(1.College of Chemistry and Life Sciences, Integrated Mountain Research Institute,Guizhou Education University, Guiyang 550018,China;2.State Key Laboratory for Biocontrol, Sun Yat-sen University, Guangzhou 510275, China; 3.Academe of Food and Health Engineering, Sun Yat-sen University, Guangzhou 510275; 4.Guangdong Key Laboratory of Integrated Pest Management in Agriculture, Guangzhou 510275)
In order to determine the diversity of ground-dwelling arthropods in the habitat ofCordycepssinensis,pitfall trap methods were used to collect arthropods in three different meadow types in Mt.Segrila of Tibet, from June 2009 to November 2010.Species accumulation curves are used and the results showed that sampling effort is sufficient.A total of 114 species belonging to 14 orders and 62 families were identified.There were obvious differences in individual number distribution of ground-dwelling arthropod assemblages among three meadow types; however, the similar taxa were obtained in different habitats.The most dominant family was Tenebrionidae, Formicidae and Tomoceridae in Slope meadow, Forest meadow and Typical meadow, respectively.All values of diversity indices (richness, diversity and equitability) were in sequence of Forest meadow > Slope meadow > Typical meadow.In conclusion, the community composition of ground-dwelling arthropods have a closely relationship with thermal condition and artificial disturbance in the microhabitat ofCordycepssinensis.
Mt.Segrila;Cordycepssinensis; alpine meadow; ground-dwelling arthropods; diversity
喻浩,张剑霜,吴文静,等.色季拉山冬虫夏草适生地草甸地表节肢动物群落多样性[J].环境昆虫学报,2017,39(3):556-565.
国家科技支撑计划课题(2011BAI13B06);贵州省科学技术基金项目(黔科合J字〔2014〕2146号);贵州师范学院博士基金(13BS018)
喻浩,男,1986年生,博士,副教授,从事昆虫纲、蛛形纲等节肢动物的多样性研究,E-mail: insect1986@126.com
*通讯作者Author for correspondence, E-mail: zhanggr@mail.sysu.edu.cn
Received: 2016-04-08; 接受日期Accepted: 2016-06-14
Q968.1
A
1674-0858(2017)03-556-09
