退化中的长白山西坡灌木苔原优势种分布差异
2017-07-20靳英华许嘉巍王绍先王嫒林张英洁刘丽杰贺红士
靳英华,许嘉巍,*,王绍先,王嫒林,张英洁,金 慧,赵 莹,刘丽杰,尹 航,贺红士,3
1 东北师范大学地理科学学院, 长春 130024 2 长白山科学研究院,二道白河 133613 3 密苏里大学自然资源学院,哥伦比亚 65211
退化中的长白山西坡灌木苔原优势种分布差异
靳英华1,许嘉巍1,*,王绍先2,王嫒林1,张英洁1,金 慧2,赵 莹2,刘丽杰2,尹 航2,贺红士1,3
1 东北师范大学地理科学学院, 长春 130024 2 长白山科学研究院,二道白河 133613 3 密苏里大学自然资源学院,哥伦比亚 65211
长白山苔原带植被正在发生显著变化,灌木苔原中灌木植物分布范围萎缩,重要值下降。通过样方调查数据,分析灌木苔原中优势种的变化,灌木分布格局和灌木群落结构特征沿海拔的差异,旨在揭示长白山灌木苔原退化的区域差异,为明确其退化机理提供基础数据。研究表明:(1)长白山西坡灌木苔原退化严重,多种草本植物已经侵入,并成为优势种。目前7个优势种中灌木仅占2席,草本植物占据5席,与1979年的样方调查结果相比灌木优势种的数量和地位都明显下降。7个优势种均为聚集分布,各优势种分布呈现斑块化、分离化,统一的灌木苔原面临解体;大部分灌木苔原群落中,出现了草本层,苔原带下部灌木苔原中草本层高于灌木层,物种组成和群落形态接近草木苔原。(2)灌木在各海拔均仍有广泛分布,但其空间分布格局明显不同。在海拔2300m以下,灌木的分布产生较强的聚集现象,特别是在海拔2100m以下这种聚集分布现象更为突出;在海拔2300m以上灌木的聚集程度较弱。(3)长白山西坡灌木苔原退化的区域分异明显,在海拔2100m以下灌木苔原退化严重,成为草-灌苔原;在海拔2100—2300m之间,灌木苔原退化较严重,成为灌-草苔原;在海拔2300m以上,退化较轻,仍为灌木苔原。由此推断,长白山西坡灌木苔原的退化机理应包括两个方面:草本植物入侵,种间竞争导致灌木退化,以及环境变化导致灌木退化,二者皆可能是全球气候变化的结果。
长白山;灌木苔原;优势种;退化;区域分异
1 研究背景
长白山高山苔原是我国最为典型的山地苔原,位于长白山火山锥体上部(2000—2500m)[1、2]。根据前人调查,长白山高山苔原以灌木苔原为主要类型。灌木苔原植物种类较少,极地或高山种属约占80%,建群植物以小灌木为主,牛皮杜鹃(Rhododendronaureum)和笃斯越橘(Vacciniumuliginosum)是常见的建群种,群落简单,只有灌木层和苔藓、地衣两层,灌木层高约8—22cm[2]。长白山高山苔原还少量分布着草木苔原[3],草木苔原也被称为草本-灌木苔原[2],群落组成更为单一,以大白花地榆(Sanguisorbastipulata)和小白花地榆(Sanguisorbaparviflora)等草本植物为代表,具有湿生性[3]。
过去30年来,长白山灌木苔原在植被组成和分布格局方面都发生了显著的变化,灌木苔原中,一些草本植物如大白花地榆(Sanguisorbastipulata)、小白花地榆(Sanguisorbaparviflora)、小叶章(Deyeuxiaangustifolia)、单花橐吾(Ligulariajamesii)、高岭风毛菊(Saussureaalpicola)、长白老鹳草(Geraniumbaishanense)、尖被藜芦(Veratrumoxysepalum)、大苞柴胡(Bupleurumeuphorbioides)等大量侵入,多度增加,形成草本层,成为常见种甚至优势种。在部分地区通过种间竞争,成功取代灌木,形成草本植物斑块[4-8];与之相反,牛皮杜鹃、笃斯越橘、越桔(Vacciniumvitis-idaea)、松毛翠(Phyllodocecaerulea)、宽叶仙女木(Dryasoctopetalavar.asiatica)、毛毡杜鹃(Rhododendronconfertissimum)等灌木植物分布范围明显萎缩,多度和重要值下降[9],灌木苔原呈现退化趋势。
在全球气候变化背景下,苔原带植被敏感,变化显著,高山苔原植被的变化更为领先[10-11],所以高山苔原植被的变化对全球气候变化具有指示意义。
现已证实,欧洲山地苔原草本植物入侵和北极的苔原带灌木分布区向北扩展为对全球气候变化的响应[12-14]。目前有关长白山灌木苔原退化的研究很少,退化机理尚无定论。本研究通过调查长白山西坡灌木苔原优势种的分布,揭示灌木苔原退化的区域差异,分析长白山灌木苔原退化机理,为提出灌木苔原退化为全球气候变化响应的一般模式观点提供基础数据。
2 材料与方法
2.1 研究区概况
长白山(41°23′N—42°36′N,126°55′E—129°E)地处吉林省东南,为中国和朝鲜两国的界山,是中国东北最高山峰(2691m)。由于地势高峻,气候、植被和土壤成明显的垂直带分布。从下而上为山地针阔混交林带、山地针叶林带、亚高山岳桦林带和高山苔原带[15]。
长白山苔原带地表多为碱性粗面岩风化物和少量火山灰,地貌为流水改造的火山锥坡面,土壤为薄层山地苔原土,气候为寒带气候。
2.2 研究方法
2.2.1 样地调查
参照钱家驹等于1979年在长白山西坡苔原带对灌木苔原进行植物样方调查的位置,2014年8月在长白山西坡苔原带海拔2050—2550m布设了一个长2250 m、宽100 m的调查样地(图1),进行了系统样方调查,2015年8月做了补充调查。在样地内,沿着海拔升高的方向,每隔50m,横向设置100m的样条,样条两侧各留出5m,间隔30m均匀布设4个1m×1m的控制性样方,共计46×4=184个控制样方,记录经纬度、海拔(由GPS测得)、植物群落特征(植物种、株数、株高、盖度)。
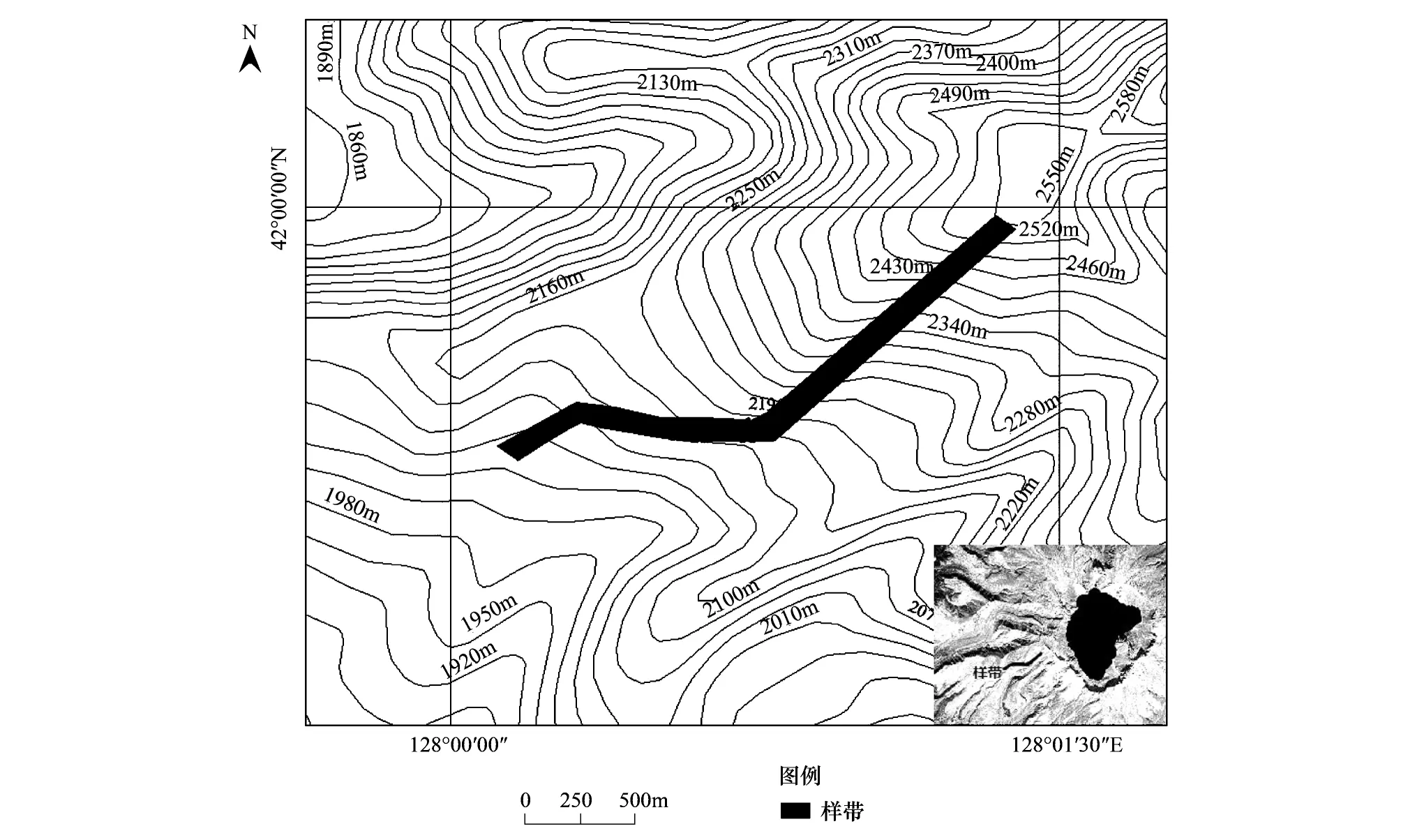
图1 样带位置图Fig.1 Transect location map
2.2.2 数据处理和分析
重要值=[相对密度+相对频度+相对盖度]/3。其中,相对密度=某种植物的个体数/全部植物个体数×100%;相对频度=某种植物的种群频度/所有种群频度之和×100%;相对盖度=某种植物的种群盖度/所有种群盖度之和×100%。
采用扩散系数(DI) 测定分布格局类型, 然后用t检验判断分布格局的显著性。
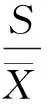
(1)
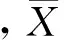
采用聚集强度负二项参数(K)判定聚集强度。
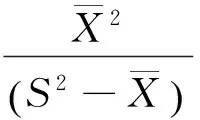
(2)
式中,K值愈小,聚集度愈大。如果K值较大(一般为8以上),则接近随机分布。
利用Excel软件对各物种的密度、频度和盖度进行统计。
3 结果与分析
3.1 调查样地优势种的变化
调查样地内种子植物种类共78种,隶属22科。鉴于钱家驹记载了灌木苔原有7个优势种,本研究也选取重要值前7位的优势种,进行优势种变化比较。7个优势种按重要值大小依次为牛皮杜鹃、大白花地榆、小白花地榆、小叶章、高岭风毛菊、单花橐吾、笃斯越橘(表1)。
钱家驹记载的灌木苔原7个优势种为牛皮杜鹃、笃斯越橘、越桔、松毛翠、宽叶仙女木、毛毡杜鹃、大白花地榆[4],其中灌木6种,草本1种,此次调查的7个优势种中灌木只有2种,草本5种,变化十分明显。虽然灌木牛皮杜鹃依然为最主要的优势种,但多种草本植物已经占据主导,例如小白花地榆、大白花地榆和小叶章等。灌木笃斯越橘、越桔和宽叶仙女木的重要值分别为第7、11和59位,其它灌木种类在样地中偶见,苔原带灌木的优势地位已经下降,灌木苔原向草木苔原转化。
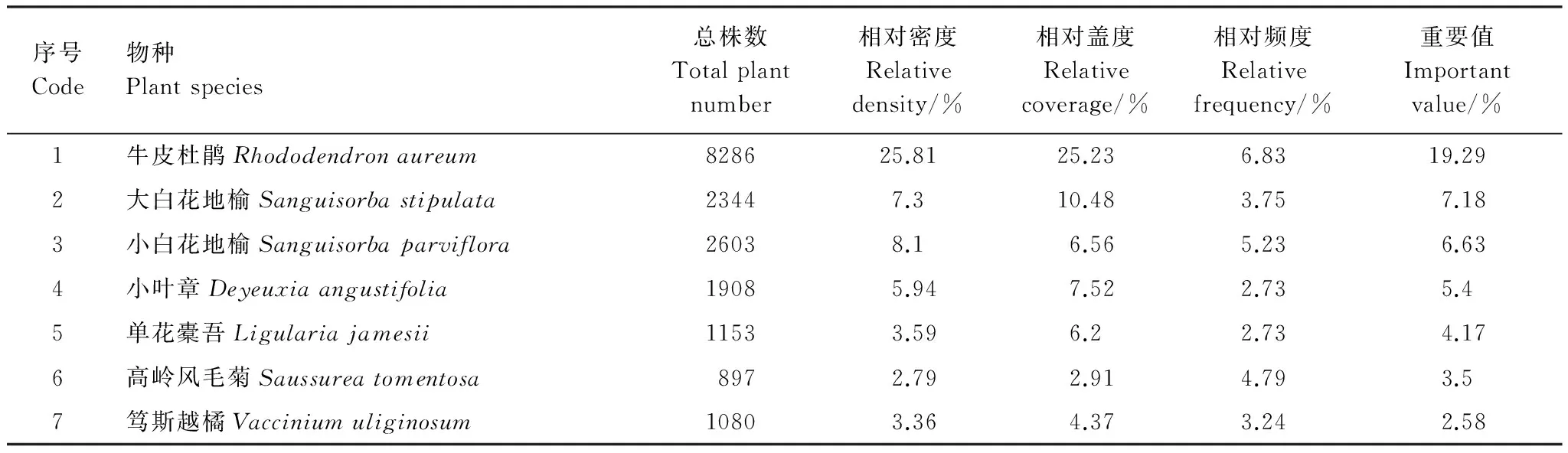
表1 长白山高山苔原带植物群落各种群重要值
3.2 调查样地优势植物的聚集特征
7种优势植物的扩散系数(DI)均大于1,且都通过T检验,可以判定7种优势植物的分布格局都属于聚集分布(表2)。牛皮杜鹃的K值最大,聚集强度最弱;笃斯越橘的K值最小,聚集强度最强。大多数草本植物的K值都小,聚集强度都较强(表2)。各优势种分布斑块化,统一的灌木苔原面临解体。
3.3 调查样地群落结构的分布特征
苔原带不同海拔高度上灌木和草本优势种的多度差异大,群落结构随海拔发生变化。在海拔2100m以下,灌木多度较少,以牛皮杜鹃和笃斯越橘为主,草本的多度最大,超过灌木,为草本-灌木苔原;在海拔2100—2300m之间,灌木以牛皮杜鹃和笃斯越橘为主,草本数量较多,但多度少于灌木,为灌木-草本苔原;在海拔2300—2500m之间,草本数量较少,为灌木苔原。其中2300—2400m灌木苔原以牛皮杜鹃和笃斯越橘为主,盖度较大,为典型的灌木苔原(图2)。在海拔2400m以上灌木的种类构成发生变化,牛皮杜鹃多度比例减少,而个体小、更低矮的松毛翠、宽叶仙女木等增多,灌木盖度较小,为稀疏的灌木苔原(图3)。

表2 优势植物空间格局类型
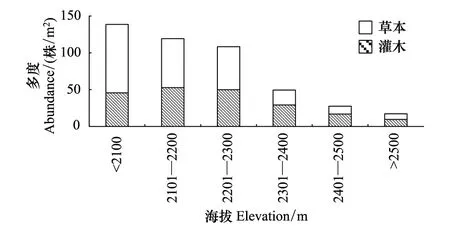
图2 群落中灌草多度对比随海拔变化图 Fig.2 Contrast on abundance between shrub and herb with elevation
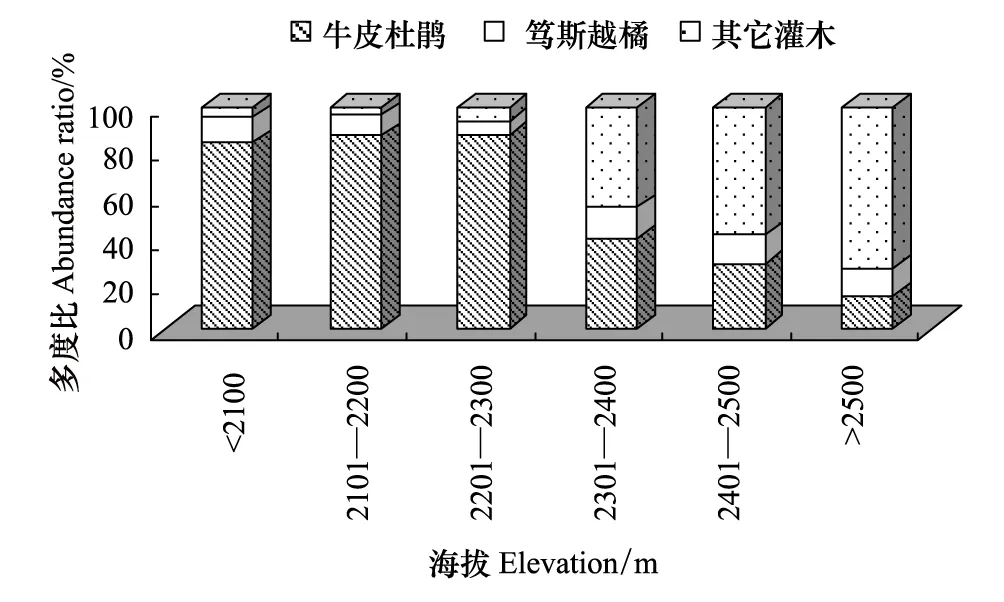
图3 群落中灌木组成随海拔变化图 Fig.3 Change of shrub community composition with elevation
在调查样地中灌木缺失的样方占到28.2%,海拔2300m以下灌木缺失的样方占到31.8%。海拔2300m以上灌木缺失的样方占到19.2%。
以灌木为主(灌木盖度超过50%)的样方比例在不同海拔有显著差异,灌木为主的样方比例在海拔2100m以下低于50%,随海拔升高灌木为主的样方比例在显著增加,特别是在海拔2300—2400m灌木为主的样方比例接近80%,在海拔2400m以上灌木为主的样方比例大幅度下降(高海拔区具有植被盖度小,甚至裸地的区域)(表3)。
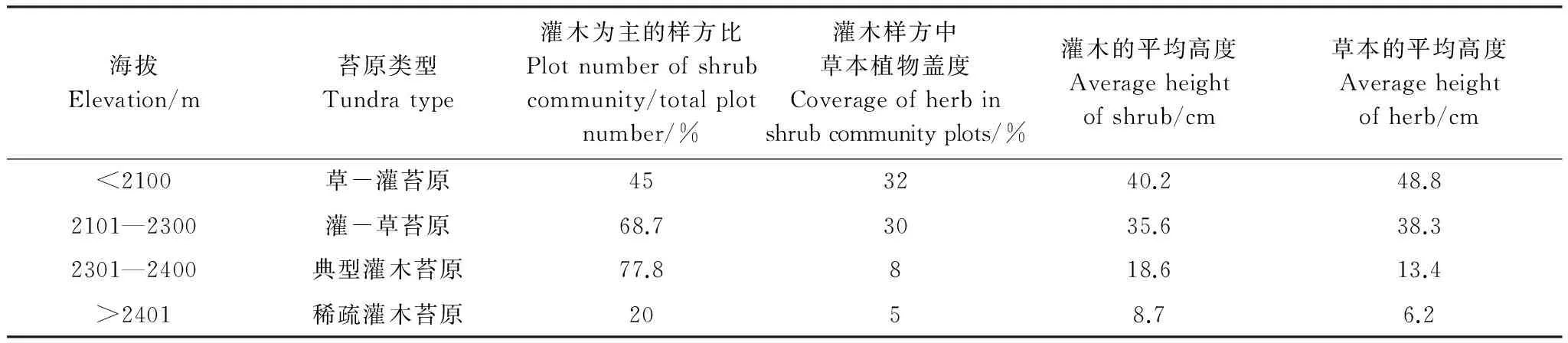
表3 调查样地灌木群落的结构
灌木为主的样方中在海拔2300m以下草本盖度超过30%,在海拔2300m以上草本盖度低于10%。以牛皮杜鹃和笃斯越橘为优势种的灌木群落虽仍占据苔原带最大面积,但群落内出现了草本层,随海拔的升高灌木和草本的高度都在降低,但在海拔2300m以下草本层的高度高于灌木层,在海拔2300m以上草本层的高度低于灌木层(表3)。
3.4 调查样地灌木和草本优势种聚集程度的空间差异
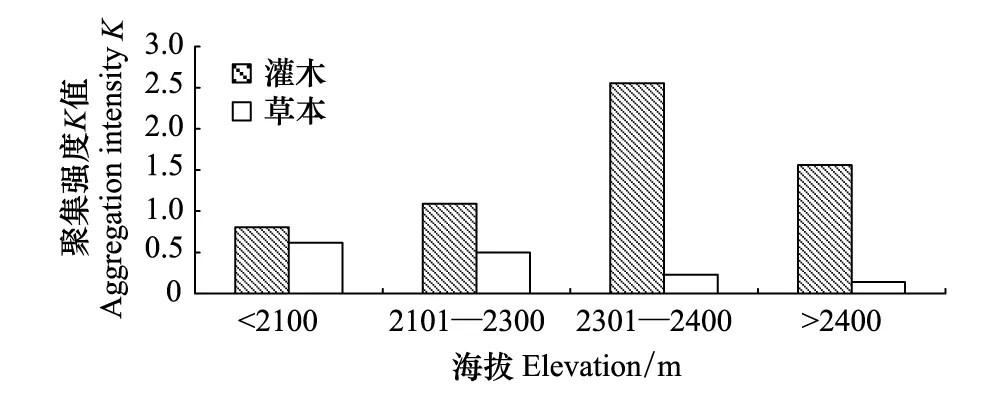
图4 灌木和草本优势种聚集程度随海拔的变化 Fig.4 Aggregation intensity of dominant shrubs and herbs plant species with elevation
灌木在各海拔均有广泛分布,但其空间分布格局明显不同(图4)。在海拔2300m以下,灌木和草本的聚集强度K值都小,且差异不大,说明在苔原带的下部灌木和草本都为较强的聚集分布。由于草本植物入侵,草本与灌木之间存在较强的种间竞争,草本植物占据了部分空间,迫使牛皮杜鹃等灌木的分布产生较强的聚集现象。特别是在海拔2100m以下,灌木的分布聚集现象更为突出。在海拔2300m以上,灌木的聚集强度K值大,草本的聚集强度K值很小,两者差异大,说明在苔原带的上部,灌木分布的聚集强度较弱,而草本是更为聚集分布的。在海拔2300m以上受到草本植物的影响小,灌木的聚集程度弱,但在海拔2400m以上,苔原植被覆盖度较低,有裸地和苔藓地衣斑块出现,灌木呈现一定程度的聚集。
4 讨论
长白山西坡灌木苔原植被发生显著变化,其退化机理应包括两个方面:草本植物入侵,种间竞争导致灌木退化,以及环境变化导致的灌木退化,二者皆可能是全球气候变化的结果。
长白山西坡苔原带研究样地30年前为灌木苔原,主要是灌木和苔藓地衣,很少有草本植物[4、5],目前草本植物已经占据全域,形成草本层片,草本植物主要为林线附近的林下喜温草本植物。在过去几十年里,很多高山、亚高山地区的低海拔物种已经向高海拔扩展[11,16-20],并且草本植物向上扩展的更明显[21- 23],这种植被变化的原因主要是对全球气候变暖的响应。
生长季温度明显升高,积温增加,生长期延长,引起的苔原气候状况改进,促使喜温的草本分布趋于更高的海拔,这种变化短时间内引起苔原带生物多样性增加[24];但随着时间的推移,温度升高后环境发生改变,导致本地物种竞争能力下降,在与外来植物竞争的过程中逐渐消失并被取代[25]。长白山灌木苔原植被以多年生小灌木为主,小灌木植株矮小、匍匐贴地、强烈分枝,交织成网,形成密集而松软的垫状植物,这种密集的垫状植物有利于保温,可以在气温低的情况下形成温度较高的近地面小气候环境且有防风的功能。小灌木为中位芽(地上芽)植物,第二年可迅速返青;而草本不具备这些形态特征,又为地面和地下芽植物,返青不快。因此,在长白山苔原带低温条件下,灌木和草本植物竞争中,灌木占据明显优势。目前长白山苔原带温度升高,生长季延长[26],使灌木优势明显下降,草本植物生长快,植株高的优势意义加强。温度升高,导致草本植物入侵,种间竞争导致灌木退化。
长白山苔原带的冬季气温升高显著,雪被厚度下降,春季升温快,积雪融化提前[26]。积雪变化对灌木植物影响强烈,雪被厚度下降,不利于中位芽的小灌木越冬,冻害加剧;积雪融化提前,也可导致灌木返青时土壤水分不足,使灌木生长不良,产生退化[14]。
已有研究表明,欧亚、北美北部的苔原带气温升高幅度大,南部的森林在向北部的苔原带扩张[27]。如果在欧亚、北美北部的苔原带上,未来出现我们在高山苔原发现的草甸化现象,森林下草本植物率先进入苔原带,引起草甸化,就可能抑制森林的扩张。引起碳平衡、雪被、反照率、土壤持水性等一系列的变化,产生不可估量的复合作用。
5 结论
(1)长白山西坡灌木苔原退化严重,多种草本植物已经侵入,并成为优势种。1979年的样方调查表明,样地的7个优势种中6种为灌木,但目前的7个优势种中,草本植物占据5席,灌木仅占2席;7个优势种均为聚集分布,各优势种分布斑块化、分离化,统一的灌木苔原面临解体;部分(苔原带下部)灌木苔原群落中,出现了草本层,严重退化的灌木苔原草本层的高度高于灌木层,物种组成和群落形态接近草木苔原。
(2)灌木在各海拔均仍有广泛分布,但其空间分布格局明显不同。在海拔2300m以下,草本植物与灌木之间存在较强的种间竞争,草本植物占据了部分空间,迫使灌木的分布产生较高的聚集现象。特别是在海拔2100m以下,灌木的分布聚集现象更为突出;海拔2300m以上受到草本植物的影响小,灌木的聚集程度小。
(3)长白山西坡灌木苔原退化的区域分异明显,在海拔2100m以下,草本的多度和盖度超过灌木的多度和盖度,灌木为主的样方比例低于50%,灌木苔原退化严重,成为草-灌苔原;在海拔2100—2300m之间,灌木苔原退化较严重,草本数量较多,但多度少于灌木,成为灌-草苔原;在海拔2300m以上,退化较轻,草本数量较少,仍为灌木苔原。
综上,长白山西坡灌木苔原带退化机理应包括两个方面:草本植物入侵,种间竞争导致灌木退化,以及环境变化导致的灌木退化,二者皆可能是全球气候变化的结果。
[1] 黄锡畴, 刘德生, 李祯. 长白山北侧的自然景观带. 地理学报, 1959, 25(6): 435- 446.
[2] 黄锡畴, 李崇皜. 长白山高山苔原的景观生态分析. 地理学报, 1984, 39(3): 285- 297.
[3] 郎惠卿, 李桢. 长白山的植物地理//长白山地理系统研究 (一). 长春: 东北师范大学出版社, 2010: 202- 208.
[4] 钱家驹, 张文仲. 长白山高山冻原植物的调查研究简报(I). 东北师大学报: 自然科学版, 1980, (1): 49- 65.
[5] 钱宏. 长白山高山冻原植物群落的数量分类和排序. 应用生态学报, 1990, 1(3): 254- 263.
[6] 许嘉巍, 张飞虎. 长白山自然地理研究的几个主要问题//长白山地理系统与生态安全(四). 长春:东北师范大学出版社, 2011: 266- 274.
[7] 靳英华, 许嘉巍, 梁宇, 宗盛伟. 火山干扰下的长白山植被分布规律. 地理科学, 2013, 33(2): 203- 208.
[8] 宗盛伟, 许嘉巍, 吴正方. 长白山西坡小叶章侵入苔原带调查与机理分析. 山地学报, 2013, 31(4): 448- 455.
[9] 宗盛伟, 许嘉巍, 吴正方, 乔琳琳, 王丹丹, 孟祥君, 杜海波, 王雷, 王丹, 王鹏, 敖小龙, 夏毓璘, 吴福梅. 长白山西坡小叶章侵入苔原带过程及影响. 生态学报, 2014, 24(23): 6837- 6846.
[10] Danby R K, Hik D S. Variability, contingency and rapid change in recent subarctic alpine tree line dynamics. Journal of Ecology, 2007, 95(2): 352- 363.
[11] Bjorkman A D, Elmendorf S C, Beamish A L, Vellend M, Henry G H R. Contrasting effects of warming and increased snowfall on Arctic tundra plant phenology over the past two decades. Global Change Biology, 2015, 21(12): 4651- 4661.
[12] Klanderud K, Birks H J B. Recent increases in species richness and shifts in altitudinal distributions of Norwegian mountain plants. The Holocene, 2003, 13(1): 1- 6.
[13] Wardle D A, Gundale M J, Jäderlund A, Nilsson M C. Decoupled long-term effects of nutrient enrichment on aboveground and belowground properties in subalpine tundra. Ecology, 2013, 94(4): 904- 919.
[14] Myers-Smith I H, Forbes B C, Wilmking M, Hallinger M, Lantz T, Blok D, Tape K D, Macias-Fauria M, Sass-Klaassen U, Lévesque E, Boudreau S, Ropars P, Hermanutz L, Trant A, Collier L S, Weijers S, Rozema J, Rayback S A, Schmidt N M, Schaepman-Strub G, Wipf S, Rixen C, Ménard C B, Venn S, Goetz S, Andreu-Hayles L, Elmendorf S, Ravolainen V, Welker J, Grogan P, Epstein H E, Hik D S. Shrub expansion in tundra ecosystems: dynamics, impacts and research priorities. Environmental Research Letters, 2011, 6(4): 45509- 45523.
[15] 赵大昌. 长白山的植被垂直分布带. 森林生态系统研究, 1980, (1): 65- 70.
[16] McDougall K L, Morgan J W, Walsh N G, Williams R J. Plant invasions in treeless vegetation of the Australian Alps. Perspectives in Plant Ecology, Evolution and Systematics, 2005, 7(3): 159- 171.
[17] Beckage B, Osborne B, Gavin D G, Pucko C, Siccama T, Perkins T. A rapid upward shift of a forest ecotone during 40 years of warming in the Green Mountains of Vermont. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(11): 4197- 4202.
[18] Kelly A E, Goulden M L. Rapid shifts in plant distribution with recent climate change. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(33): 11823- 11826.
[19] Parolo G, Rossi G. Upward migration of vascular plants following a climate warming trend in the Alps. Basic and Applied Ecology, 2008, 9(2): 100- 107.
[20] Odland A, Høitomt T, Olsen S L. Increasing vascular plant richness on 13 high mountain summits in southern Norway since the early 1970s. Arctic, Antarctic, and Alpine Research, 2010, 42(4): 458- 470.
[21] Pauli H, Gottfried M, Reiter K, Klettner C, Grabherr G. Signals of range expansions and contractions of vascular plants in the high Alps: observations (1994- 2004) at the GLORIA master site Schrankogel, Tyrol, Austria. Global Change Biology, 2007, 13(1): 147- 156.
[22] Walther G R, Beißner S, Burga C A. Trends in the upward shift of alpine plants. Journal of Vegetation Science, 2005, 16(5): 541- 548.
[23] Thuiller W, Richardson D M, Midgley G F. Will climate change promote alien plant invasions?//Nentwig W, ed. Biological Invasions. Berlin Heidelberg: Springer, 2006: 197- 211.
[24] Erschbamer B, Unterluggauer P, Winkler E, Mallaun M. Changes in plant species diversity revealed by long-term monitoring on mountain summits in the Dolomites (northern Italy). Preslia, 2011, 83(3): 387- 401.
[25] Halloy S R P, Mark A F. Climate-change effects on alpine plant biodiversity: a New Zealand perspective on quantifying the threat. Arctic, Antarctic, and Alpine Research, 2003, 35(2): 248- 254.
[26] 宗盛伟, 吴正方, 杜海波. 近52a长白山苔原带生长季气候变化特征. 干旱区研究, 2013, 30(1): 41- 49.
[27] Wipf S, Stoeckli V, Bebi P. Winter climate change in alpine tundra: plant responses to changes in snow depth and snowmelt timing. Climatic Change, 2009, 94: 105- 121.
Distribution variations of dominant plant species in degraded shrub tundra on the western slope of the Changbai Mountains
JIN Yinghua1, XU Jiawei1,*, WANG Shaoxian2, WANG Ailin1, ZHANG Yingjie1, JIN Hui2, ZHAO Ying2, LIU Lijie2, YIN Hang2, HE Hongshi1,3
1SchoolofGeographicalScience,NortheastNormalUniversity,Changchun130024,China2ChangbaiMountainAcademyofSciences,Erdaobaihe133613,China3SchoolofNaturalResources,UniversityofMissouri,Columbia,MO65211,USA
Tundra vegetation of the Changbai Mountains is undergoing significant changes. In the alpine tundra on the western slopes of the mountain, the area of distribution of shrubs is shrinking and the importance of shrubs as dominant plant species is reducing significantly. This study analyzed changes of dominant plant species in the shrub tundra, distribution pattern variations of shrubs and characteristics of shrub community structure at different elevations using plot survey data, revealed regional differentiation of degradation in shrub tundra and the degradation mechanisms of shrub communities. The results showed the following: (1) The shrub community in the alpine tundra on the western slopes has been severely degraded. Various herbs have invaded the area and became the dominant species. Five herbs and two shrub species were the dominant species of alpine tundra, which indicated a decrease in the number and importance value of dominant shrub species compared to the results of the survey conducted in 1979. The aggregated distribution was the dominant pattern of distribution of the seven dominant species, whereas the distribution of each dominant species occurred in separated patches; the shrub tundra faced fragmentation. In shrub communities of alpine tundra, herbs appeared and gradually became dominant. The herbs were taller than the shrubs in alpine tundra at lower elevations, and the species composition and community form were nearing those of the herb-shrub tundra. (2) Shrubs were widely distributed at different elevations, but their spatial distribution patterns were distinct. Below 2300 m a.s.l., the distribution of shrubs appeared aggregated, especially at elevations below 2100 m a.s.l. The shrubs did not aggregate above 2300 m a.s.l. (3) Regional differentiation of the degraded alpine tundra on the western slopes was significant. Below 2100 m a.s.l., the number of herbs was more abundant than that of shrubs, and the coverage and height of the shrubs were decreased; so the degradation is the most serious in this region, which developed into herb-shrub tundra. Between 2100 and 2300 m a.s.l., the degradation was significant, but the number of shrubs were more abundant than that of herbs; this region developed into shrub-herb tundra. Above 2300 m a.s.l., the degradation of shrubs was not significant and shrub tundra was preserved. It can be inferred that the degradation mechanism of the shrub tundra includes two aspects: herb invasion of the shrub tundra, resulting in interspecific competition, and environmental changes of tundra zone, which could be attributed to global climate change.
Changbai Mountains; shrub tundra; dominant plant species; degradation; regional differentiation
长白山科学研究院开放基金课题(201501);国家自然科学基金资助项目(41571078,41171072)
2016- 03- 19; 网络出版日期:2017- 02- 22
10.5846/stxb201603190489
*通讯作者Corresponding author.E-mail: xujw634@nenu.edu.cn
靳英华,许嘉巍,王绍先,王嫒林,张英洁,金慧,赵莹,刘丽杰,尹航,贺红士.退化中的长白山西坡灌木苔原优势种分布差异.生态学报,2017,37(11):3716- 3723.
Jin Y H, Xu J W, Wang S X, Wang A L, Zhang Y J, Jin H, Zhao Y, Liu L J, Yin H, He H S.Distribution variations of dominant plant species in degraded shrub tundra on the western slope of the Changbai Mountains.Acta Ecologica Sinica,2017,37(11):3716- 3723.
