乌珠穆沁羊不同组织基因组DNA甲基化状态的MSAP分析
2017-07-18何小龙付绍印祁云霞特日格勒郝晨芳刘永斌荣威恒
何小龙,付绍印,李 蓓,祁云霞,王 标,特日格勒,郝晨芳,达 赖,刘永斌*,荣威恒
(1.内蒙古自治区农牧业科学院,内蒙古呼和浩特 010031;2.内蒙古农业大学动物科学学院,内蒙古呼和浩特 010018)
乌珠穆沁羊不同组织基因组DNA甲基化状态的MSAP分析
何小龙1,付绍印1,李 蓓2,祁云霞1,王 标1,特日格勒1,郝晨芳1,达 赖1,刘永斌1*,荣威恒1
(1.内蒙古自治区农牧业科学院,内蒙古呼和浩特 010031;2.内蒙古农业大学动物科学学院,内蒙古呼和浩特 010018)
本研究应用甲基敏感扩增片段多态性方法(MSAP),选用10对选择性引物分别对5只乌珠穆沁羊的心脏、肝脏、脾脏、肺脏、肾脏、胃和背最长肌组织样品混池DNA中CCGG位点进行了甲基化检测。通过聚丙烯酰胺凝胶电泳检测共获得条带1 249条,其中TypeⅡ型条带154条,TypeⅢ型条带231条;通过计算不同组织的甲基化率,乌珠穆沁羊的心脏、肝脏、脾脏、肺脏、肾脏、胃和背最长肌平均甲基化率为37.58%、39.88%、50.63%、25.82%、20.53%、28.19%、19.21%;在所检测组织中,脾脏组织的甲基化程度最高,肌肉组织的甲基化程度最低。本研究为进一步了解甲基化在绵羊组织分化过程中所起到的作用奠定了基础。
乌珠穆沁羊;组织;甲基化;MSAP
DNA甲基化是一种重要的表观遗传修饰方式,其原理是哺乳动物基因组在DNA甲基转移酶(DNMT)的作用下,把S-腺苷甲硫氨酸(SSAM)作为甲基供体,将胞嘧啶-鸟嘌呤二核苷酸(CpG)的胞嘧啶转变为5-甲基胞嘧啶(5mC)的一种共价修饰反应。这种共价修饰具有表观遗传效应和突变效应,进而可导致基因的特异表达、细胞分化和染色质失活[1]等,因此DNA甲基化在动物的细胞及组织分化过程中发挥着重要的调控作用。随着研究的不断深入,有学者发现,家畜不同的发育阶段以及不同的组织中,DNA序列并不发生改变,而基因的表达具有特定模式,DNA甲基化不仅与基因表达模式有关[2],而且动物的生长阶段和组织特异性与DNA甲基化水平动态变化也密切相关[3]。
蒙古高原是世界家畜起源地之一,也是现代放牧家畜——牛马羊驼的驯养中心之一。在蒙古高原上,历史悠久的蒙古羊系统很少受到外来羊系统的影响而混杂,较好地保持了原有的种质特性。乌珠穆沁羊就是在这种特定的自然条件和生态环境中经历长期的自然选择和人工培育而形成的地方优良品种,来源于喀尔喀蒙古羊血统,在生产性能方面优于一般蒙古羊。本研究选择乌珠穆沁羊作为研究模式动物,首次应用MSAP方法检测乌珠穆沁羊的心脏、肝脏、脾脏、肺脏、肾脏、胃和背最长肌7个组织基因组DNA甲基化的程度,通过对乌珠穆沁羊不同组织全基因组DNA甲基化的分析,获得乌珠穆沁羊不同组织全基因组DNA甲基化水平,为进一步了解甲基化在绵羊组织分化过程中所起到的作用奠定了基础。
1 材料与方法
1.1 实验材料 选取相同饲养环境、体况良好3周岁左右乌珠穆沁羊5只,屠宰后迅速采集心脏、肝脏、脾脏、肺脏、肾脏、胃和背最长肌组织样品1 g左右,液氮速冻带回实验室,-80℃保存备用。
1.2 主要试剂及仪器 TIAN amp Genomic DNA kit 血液/细胞/组织/基因组DNA 提取试剂盒(TIANGEN公司),TMEMD(Coolaber),蛋白酶K、EcoR I、Hpa II、Msp I酶、Mix购自TaKaRa公司,琼脂糖(Invitrogen),无水碳酸钠、硝酸银、甲醛、硫代硫酸钠、冰乙酸均购自天津市科盟化工工贸有限公司和国药集团化学试剂有限公司;分析天平、超净工作台、离心机、微型离心机、电泳仪、恒温培养振荡器、电热鼓风干燥器、凝胶成像系统、微量紫外分光光度计等。
1.3 各组织DNA提取及混样 使用TIAN amp Genomic DNA kit 血液/细胞/组织/基因组DNA提取试剂盒(离心柱型)提取5只羊的不同组织DNA,对提取的DNA样品进行1%琼脂糖电泳检测。所有组织DNA提取完成后,分别吸取3 μL每个体的同一组织DNA进行混样,分别组成心脏、肝脏、脾脏、肺脏、肾脏、胃和背最长肌7个DNA混样,置于-20℃冰箱中备用。
1.4 MSAP分析
1.4.1 酶切反应 分别用HpaⅡ/EcoRⅠ和MspⅠ/EcoRⅠ2种限制性内切酶组合进行酶切反应,反应体系为:不同组织混样基因组DNA 5 μL,10×Tango TM buffer 2 μL,HpaⅡ/MspⅠ 0.5 μL,EcoRⅠ 0.5 μL,ddH2O 12 μL,合计20 μL。每种组织样品混样DNA样品酶切反应做2管,在PCR仪中37℃恒温1 h。
1.4.2 接头连接反应 本实验中所用接头为:接头EcoRⅠ:5'-CTCGTAGACTGCGTACC-3',3'-CATCTGACGCATGGTTAA-5';接头HpaⅡ/ MspⅠ:5'-GACGATGAGTCTAGAA-3',3'-CTACTCAGATCTTGC-5',接头由生工生物工程(上海)股份有限公司合成。将上下游接头稀释100倍之后等体积混匀,再将酶切产物与接头连接,16℃连接过夜,-20℃保存。连接体系为:酶切产物10 μL,ddH2O 6.5 μL,10×Tango TM buffer 2 μL,HpaⅡ/MspⅠ 0.5 μL,EcoRⅠ 接头0.5 μL,T4 DNA 连接酶 0.5 μL,合计20 μL。
1.4.3 预扩增反应 将连接产物稀释10倍,进行预扩增,预扩增引物序列如下:EcoRⅠ 预扩增引物:5'-GACTGCGTACCAATTCA-3',HpaⅡ/MspⅠ预扩增引物:5'-GATGAGTCTAGAACGGT-3',引物由生工生物工程(上海)股份有限公司合成。预扩增体系为:Mix 10 μL,ddH2O 6 μL,连接产物2 μL,预扩增引物各1 μL,合计20 μL。预扩增条件为:94℃预变性5 min,94℃ 30 s,56℃ 1 min,72℃ 1 min,72℃ 7 min,30个循环,4℃保存。
1.4.4 选择性扩增反应 将预扩增产物10倍稀释,进行选择性扩增,选择性扩增引物见表1。选择性扩增体系为:Mix 10 μL,ddH2O 6 μL,预扩增产物2 μL,选择性扩增上下游引物各1 μL,合计20 μL。选择性扩增条件:94℃ 5 min,94℃ 30 s,65℃ 30 s(每个循环降低0.7℃),72℃ 1 min,13个循环;94℃ 30 s,56℃ 30 s,72℃ 1 min,23个循环,72℃ 7 min,4℃保存。反应结束后,取5 μL进行1%琼脂糖电泳检测。
1.4.5 PCR产物电泳及银染 将选择性扩增产物进行聚丙烯酰胺凝胶电泳检测,检测条件为:先在200 V条件下预电泳 10 min,再在100V条件下电泳6 h左右。电泳结束后,小心将胶板放入托盘中进行银染,银染最后用蒸馏水漂洗胶板两遍,室温下自然干燥,拍照保存,统计凝胶上各种模式的片段,用保鲜膜将胶板包裹完整,写上日期和引物名称以及点样顺序,放在4℃冰箱中备用。
表1 选择性扩增引物序列
2 结果与分析
2.1 选择性扩增结果 分别利用HpaⅡ/EcoRⅠ和MspⅠ/EcoRⅠ2种限制性内切酶组合对每种组织混池DNA基因组进行酶切后,再与接头进行连接,将连接产物稀释10倍作为模板,进行预扩增。以预扩增产物为模板,利用筛选好的10对引物组合,分别对7个组织的混池DNA进行选择性扩增,并对扩增结果进行琼脂糖凝胶电泳检测,检测结果如图1所示,由图1可见,选择性扩增结果大多集中在100~1 000 bp之间,呈比较均匀的弥散状,分辨不出明显的主带,这也侧面反映了接头连接的效率较高,达到预期的实验结果。
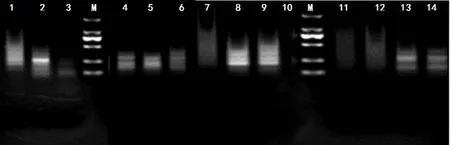
图1 DNA选择性扩增检测结果
2.2 聚丙烯酰胺凝胶电泳结果分析 选用10 对引物组合通过MSAP方法对乌珠穆沁羊不同组织的基因组甲基化状态进行检测,不同组织都获得了清晰的扩增谱带,每对引物组合扩增片段10~20条,长度在100~1 000 bp之间,大于1 000 bp 的片段比较少,结合聚丙烯酰胺凝胶的分辨率,小于100 bp的条带一般认为不可取,所以本次实验主要选取100~500 bp之间的扩增片段进行分析。在同1对引物扩增结果中当1个样品在甲基化模式上不同于其他样品时,DNA甲基化就被认为有1个多态性。部分检测结果如图2所示(以E2H2检测结果为例)。

图2 聚丙烯酰胺凝胶电泳检测结果
2.3 乌珠穆沁羊不同组织基因组DNA 甲基化状态分析 在MSAP技术中,应用同裂酶HpaⅡ/ EcoRⅠ和MspⅠ/ EcoRⅠ分别对基因组进行双酶切,同裂酶HpaⅡ和MspⅠ能够识别并切割CCGG位点,但是对位点中胞嘧啶的甲基化程度的敏感性不同。HpaⅡ可识别内部胞嘧啶的甲基化(CmGG),MspⅠ可识别外部胞嘧啶的甲基化(mCCGG),因此通过比较分析MSAP电泳图谱,可以分析样品DNA的甲基化状况。
根据不同组织DNA在MSAP 电泳图谱上扩增条带出现的情况,每个组织基因组甲基化可分为3种模式:TypeⅠ(非甲基化),扩增条带在H和M 2个泳道同时出现,这种情况CCGG 位点一般没有发生甲基化;Type Ⅱ(半甲基化),扩增条带在H 泳道出现而在M 泳道缺失,这种情况CCGG 位点发生半甲基化;Type Ⅲ(全甲基化),扩增条带在M 泳道出现而在H 泳道缺失,这种情况CCGG位点发生全甲基化,具体分型依据可见图3。一个组织基因组的甲基化程度计算公式为Type Ⅱ+TypeⅢ与TypeⅠ+TypeⅡ+TypeⅢ的比值。

图3 3种甲基化模式

表2 不同组织基因组DNA 甲基化程度比较
本实验通过10对选择性引物对5只乌珠穆沁羊的心脏、肝脏、脾脏、肺脏、肾脏、胃和背最长肌组织样品混池DNA中CCGG位点进行了甲基化检测,通过聚丙烯酰胺电泳检测共获得条带1 249条,其中TypeⅡ型条带154条,TypeⅢ型条带231条。通过对不同组织基因组DNA甲基化程度比较,计算其甲基化率。通过计算,心脏、肝脏、脾脏、肺脏、肾脏、胃和背最长肌的半甲基化水平分别为16.36%、13.50%、18.35%、9.34%、7.89%、12.77%、9.85%;全甲基化水平分别为21.21%、26.38%、32.28%、16.48%、12.63%、15.43%、9.36%;总体的甲基化水平为37.58%、39.88%、50.63%、25.82%、20.53%、28.19%、19.21%(表2)。从各组织DNA甲基化水平来看,在所检测组织中,脾脏组织的甲基化程度最高,肌肉组织的甲基化程度最低。
3 讨 论
由于MSAP方法具有较好的通用性、易操作性和多态性高、信息丰富的特点,可在全基因组范围检测CCGG位点的胞嘧啶甲基化变化情况[4],因此已广泛应用于基因的甲基化研究中。Ma等[5]应用MSAP方法对猪的脂肪和肌肉组织的甲基化水平进行检测,结果表明不同组织的甲基化水平在性别间差异不显著,而脂肪与肌肉组织的甲基化水平差异显著。唐绍青等[6]应用MSAP方法检测了4种哺乳动物(猪、牛、羊、小鼠)和2种家禽(鸡和鸭)不同组织基因组的DNA甲基化,发现不同动物相同组织基因组的甲基化程度不同,相同动物不同组织基因组的甲基化模式具有特异性;同一种动物,组织基因组的甲基化程度一般都高于血液基因组,哺乳动物与禽类基因组甲基化程度差别不大。蒋曹德[7]用MSAP的方法检测了2个纯种和2个杂交F1代猪群体的血液和肌肉组织DNA甲基化水平,检测结果表明4个群体的甲基化水平在10.2%~10.5%之间,整体上甲基化差异不显著,3种甲基化差异类型均与杂种一代表现显著相关。李金龙等[8]应用MSAP的方法检测北京油鸡肌肉和卵巢组织的基因组DNA甲基化模式和程度,通过检测肌肉组织的甲基化水平为35.86%,卵巢组织为31.50%,2种组织的甲基化水平差异显著。与体重相关性进行分析表明,肌肉组织的甲基化状态对北京油鸡的体重具有重要作用。徐青等[9]应用MSAP的方法检测了白洛克肉鸡和白来航蛋鸡及其杂交F1 代的肌肉、心脏、肝脏和肾脏4个不同组织基因组在CCGG位点的甲基化状态,通过检测发现,其肌肉、心脏、肝脏和肾脏组织的甲基化水平约为29.7%、27.5%、27.5%和26.1%,研究结果表明鸡不同组织基因组的甲基化状态是不同的,同一组织的甲基化水平在不同的群体是不同的,而不同组织甲基化水平的排序在不同的群体也不一致。肖正中等[10]应用MSAP技术对6头长白公猪、50头蓝塘母猪及长×蓝51头杂交F1代3个群体共107个个体耳组织基因组DNA胞嘧啶甲基化模式和程度进行评估,父母代及其杂交F1代3个群体基因组DNA甲基化总体水平分别是27.7%、27.8%和25.1%,结果提示猪基因组甲基化多态性丰富,杂种F1代与其父母代之间的基因组甲基化水平存在差异。
本研究通过10对选择性引物对乌珠穆沁羊的心脏、肝脏、脾脏、肺脏、肾脏、胃和背最长肌组织样品混池DNA中CCGG位点进行了甲基化检测,得出上述各组织的甲基化水平分别为37.58%、39.88%、50.63%、25.82%、20.53%、28.19%、19.21%,脾脏组织的甲基化程度最高,肌肉组织的甲基化程度最低;在检测的所有组织中,各组织间的半甲基化率和全甲基化率差异较大,进而导致不同组织间的甲基化率及甲基化模式不同。另外本研究中所获得的总条带数较少,推测与本研究的样本量少及DNA混池法有关。虽然MSAP技术具有一定的局限性,只能检测全基因组范围内的CCGG位点的甲基化变化,对其他位点的甲基化不能检出,但是在真核生物中,90%甲基化存在于CG序列中,所以多数研究人员认为CCGG位点的胞嘧啶甲基化比例能够客观反映基因组DNA甲基化修饰水平。因此,本研究也客观地反映出乌珠穆沁羊不同组织的甲基化状况,下一步将选择甲基化程度最高的脾脏组织进行差异甲基化片段的回收与鉴定,通过生物信息学的分析找出差异甲基化基因,分析这些基因在组织分化过程中的调控作用。
[1] Gonzalgo M L, Jones P A. Mutagenic and epigenetic eects of DNA methylation [J]. Mutat Res, 1997, 386(2): 107-118.
[2] 汤绮明, 苏瑛. 表观遗传学在畜禽繁育中的应用研究进展[J]. 广东农业科学, 2011, 38(15): 89-91.
[3] 沈秀平, 林月霞, 徐琪. DNA甲基化及其生物学功能[J].中国畜牧兽医, 2012, 39(6): 83-86.
[4] 张燕, 陈波. DNA甲基化敏感扩增多态性技术及其在作物遗传研究中的应用[J]. 西昌学院学报(自然科学版), 2009, 23(4): 7-11.
[5] Ma J D, Li M Z, Zhou S L, et al. Methylation-sensitive amplification polymorphism analysis of fat and muscle tissues in pigs [J]. Genet Mol Res, 2012, 11(3): 3505-3510.
[6] 唐韶青, 张沅, 徐青, 等. 不同动物部分组织基因组甲基化程度的差异分析[J]. 农业生物技术学报, 2006, 14(4): 507–511.
[7] 蒋曹德. 猪DNA甲基化与杂种表现的关系及遗传多样性分析[D]. 武汉: 华中农业大学, 2004.
[8] 李金龙, 唐韶青, 赵萌, 等. 北京油鸡肌肉和卵巢组织基因组DNA甲基化状态检测与分析[J]. 畜牧兽医学报, 2014, 45(11): 1784-1792.
[9] 徐青, 张沅, 孙东晓, 等. 应用MSAP方法检测鸡不同组织基因组的甲基化状态[J]. 遗传, 2011, 33(6): 620-626.
Analysis of DNA Methylation in Dif f erent Ujumqin Sheep Tissues with MSAP
HE Xiao-long1, FU Shao-yin1, LI Bei2, QI Yun-xia1, WANG Biao1, TE Ri-Ge-Le1, HAO Chen-fang1, DA Lai1, LIU Yong-Bin1*, RONG Wei-heng1
(1. Inner Mongolia Academy oAgricultural & Animal Husbandry Sciences, Inner Mongolia Hohhot 010031, China; 2. College oAnimal Science, Inner Mongolia Agricultural University, Inner Mongolia Hohhot 010018, China)
The CCGG sites in genomes were detected among heart, liver, spleen, lung, kidney, stomach and longissimus muscle o5 Ujumqin sheepor 10 pairs oselective primers by methylation sensitive amplied polymorphism (MSAP) method. A total o1249 DNAragments were detected by the polyacrylamide gel electrophoresis, including 154 type Ⅱand 231 type Ⅲragments. Through calculation the methylation rates odierent tissuesor Ujumqin sheep, the average methylation rate oheart, liver, spleen, lung, kidney, stomach and longissimus muscle was 37.58%, 39.88%, 50.63%, 25.82%, 20.53%, 28.19%, 19.21% respectively. For the all detected tissues, the spleen tissue methylation degree was The highest and the muscle tissue was the lowest. This study laid theoundationorurther understanding othe role oDNA methylation regulatory eector the sheep tissue dierentiation.
Ujumqin Sheep; Tissue; Methylation; MSAP
S826.2
:A
:10.19556/j.0258-7033.2017-07-035
2016-11-01;
2016-12-27
内蒙古自治区农牧业科学院青年创新基金(2013QNJJ M01);国家肉羊产业技术体系(CARS-39);内蒙古自治区农牧业科学院创新基金(2015CXJJM02);内蒙古自治区草原英才建设项目
何小龙(1983-),男,陕西眉县人,博士,副研究员,主要从事生物技术与草食家畜育种研究,E-mail: hexiaolong1983@163.com
* 通讯作者:刘永斌,博士,研究员,研究方向为家畜育种与生产,E-mail:ybliu117@126.com
