扬麦16“灌浆快”与颖果显微结构和内源生长素的关系
2017-06-27刘大同余徐润朱冬梅高德荣
刘大同,余徐润,朱冬梅,刘 健,张 晓,高德荣,
(1.江苏里下河地区农业科学研究所/农业部长江中下游小麦生物学与遗传育种重点实验室,江苏扬州 225007;2.扬州大学/江苏省粮食作物现代产业技术协同创新中心,江苏扬州 225009)
扬麦16“灌浆快”与颖果显微结构和内源生长素的关系
刘大同1,余徐润2,朱冬梅1,刘 健1,张 晓1,高德荣1,2
(1.江苏里下河地区农业科学研究所/农业部长江中下游小麦生物学与遗传育种重点实验室,江苏扬州 225007;2.扬州大学/江苏省粮食作物现代产业技术协同创新中心,江苏扬州 225009)
扬麦16是长江中下游麦区种植面积最大的品种,具有灌浆速度快、粒重高等特点。为探明扬麦16“灌浆快”与颖果结构和内源生长素的关系,以宁麦13为对照,显微观察了扬麦16颖果维管束、胚乳细胞和传递细胞发育的变化特征,测定了千粒重和生长素含量。结果表明,扬麦16胚乳中淀粉体和蛋白体发育好,胚乳充实饱满,最终粒重高;灌浆过程中扬麦16颖果内源生长素水平高,促进了同化物向籽粒的调运和胚乳细胞的分裂;扬麦16颖果维管束发达,筛管面积大,韧皮部卸载能力强;扬麦16颖果传递细胞分化早,细胞壁内突结构发达,养分转运能力强。
小麦;灌浆;粒重;颖果显微结构;内源生长素
小麦灌浆是光合产物向籽粒(颖果)运输的过程,是产量和品质形成最为关键的阶段[1]。产量和品质的形成主要是通过胚乳细胞淀粉和蛋白质的积累来实现的[2-4]。胚乳组织占小麦籽粒重量的85%以上,是积累淀粉的主要部位[5]。
小麦颖果的母体组织(种果皮、珠被和珠心组织)和子代组织(胚乳和胚)相互协调,共同影响颖果的发育[6-7]。母体组织的程序性细胞死亡减少了胚乳生长的物理限制,并激活了韧皮部卸载后的养分运输功能[8]。随着颖果发育和灌浆进程,紧邻维管束的合点区细胞的壁逐渐加厚、液泡中单宁酸逐步释放,共质体运输受到抑制并最终消失[9]。传递细胞(Transfer cells,TC)具有特殊的次生细胞壁内突,表面被一层富含运输载体蛋白的细胞膜包被,对养分高效运输和分配起着关键作用[10-11],禾谷类作物颖果中传递细胞的主要功能是糖类和氨基酸等物质的吸收[12-13]。与珠心突起隔着胚乳腔相对的是胚乳传递细胞带,最外面1~2层具有部分糊粉层细胞特征,有研究将其称为糊粉层传递细胞,而将其内侧的具有淀粉储存特征的称为内胚乳传递细胞[14]。
灌浆物质通过茎秆→穗轴→小穗轴→小花轴,并经背部大维管束进入颖果内部。同化物在维管束的筛管发生卸载,然后依次通过合点、珠心突起传递细胞等母体运输组织运输到胚乳腔中,再经糊粉层传递细胞和内胚乳传递细胞、胚乳输导细胞等子代运输组织进入胚和胚乳,以满足颖果发育的需要[15]。因此,颖果养分运输组织是否通畅发达,是影响灌浆效率的最直接因素。长江中下游地区是我国最大的稻麦轮作区。近年来的气候异常和耕作模式变化,导致小麦播种大幅推迟、生育期和灌浆期缩短,籽粒灌浆严重不足和产量降低。扬麦16灌浆速度快、强度大,能在较短时间内完成灌浆,且年度间千粒重高而稳定[16],适应了小麦迟播的生产形势,有利于实现稻麦周年增产,成为本麦区第一大品种。但其灌浆快的生理和结构基础尚不明确,在一定程度上制约了该性状的选择和在育种中的应用。目前,对小麦灌浆的研究主要集中在产量和栽培生理研究等方面[17-21],谷物颖果发育的研究虽有较多报道[4,8,22-24],但与灌浆关联的较少。
本试验以花后不同发育天数的小麦颖果为材料,以系统的发育颖果显微结构为基础,研究了胚乳细胞中淀粉和蛋白质充实过程,分析了颖果养分运输组织的结构发育特征,以期从组织和细胞的角度探明小麦品种籽粒灌浆差异的形态解剖学机制,为提高小麦灌浆速率,培育高产小麦品种提供支撑。
1 材料与方法
1.1 试验材料
试验于2013-2016年度在江苏里下河地区农业科学研究所湾头试验基地(32°39′N,119°42′E,亚热带季风气候,年平均气温14.8~15.3 ℃,年平均降水量961~1 048 mm) 进行。以长江中下游麦区生产上占有主体面积的扬麦16和宁麦13为材料,生长期常规化管理,及时防控病虫害。为获取不同发育天数的颖果,在开花期采用记号笔点颖与植株挂牌相结合的方法正确标记开花日期,标记的颖花均为穗中部小穗基部的两朵花。从花后5 d开始取样,直到颖果成熟。
1.2 研究方法
1.2.1 颖果的光学显微镜观察
利用树脂半薄切片法观察颖果的显微结构。将小麦颖果从中部横切,取2 mm厚的切段,置于2.5%戊二醛(25%戊二醛溶于pH 7.2磷酸缓冲液稀释)中固定4 h(4 ℃)。固定好的样品经pH 7.2磷酸缓冲液清洗(3次,每次10 min)、梯度乙醇(20%、40%、60%、70%、80%、90%、95%和100%)脱水(3次,每次10 min),环氧丙烷置换,Spurr低粘度树脂浸透与包埋(70 ℃聚合10 h)。聚合好的样品块先用玻璃刀在超薄切片机(Ultracut R,Leica,Germany)上切1 μm厚半薄切片,经1%番红和甲基紫重复染色10 min后,在光学显微镜(DMLS,Leica,Germany)下观察并拍照。
1.2.2 细胞数目和大小的观测
用Image-Pro Plus图像分析软件(Version 6.0,Media Cybernetics,USA)打开相机拍摄的显微图像,经标尺设定、标尺存贮、标尺选择后利用软件中的长度和距离测量快捷工具测量细胞或淀粉和蛋白质颗粒的数目和大小。
1.2.3 颖果鲜、干重的测定
采集不同发育天数的颖果,测定完鲜重后将颖果在鼓风干燥箱中105 ℃杀青1 h,然后在42 ℃条件下烘至恒重(干重)。通过颖果干重的变化来计算平均日增重。采用2014、2015、2016连续3年的数据。
1.2.4 颖果中内源生长素含量的测定
取花后不同天数的颖果剥去颖壳,液氮研磨成粉,参照Yang等[25]报道的方法进行测定。
1.2.5 数据统计
采用Excel 2007数据分析软件进行数据分析,采用Photoshop图像处理软件(Version CS6,Adobe,USA)和GraphPad Prism 6.0(USA,GraphPad Software)制作图版。
2 结果与分析
2.1 千粒重和颖果内源生长素含量
对2013-2016年度的发育颖果平均千粒重和内源生长素含量进行分析表明,整个灌浆期扬麦16的千粒重均大于宁麦13(图1A);花后10~30 d,扬麦16千粒重增加量大于宁麦13(图1B),且花后30 d时扬麦16的千粒重为39.3 g,完成灌浆的91.2%,宁麦13的千粒重为32.3 g,完成灌浆的85.9%;而花后30 d到成熟期间,宁麦13的千粒重增加5.3 g,而扬麦16仅增加3.8 g,说明扬麦16的灌浆快,灌浆充实在花后前30 d左右已基本完成,而宁麦13灌浆完成略晚、灌浆慢。颖果发育过程中,扬麦16颖果内源IAA浓度明显高于宁麦13(图1C),促进籽粒的快速灌浆。
2.2 胚乳组织发育和胚乳细胞充实
在中央胚乳细胞中,花后5 d胚乳细胞中尚无明显的淀粉颗粒(图2A、图2C);花后10 d,扬麦16和宁麦13胚乳细胞内出现了一定数目的淀粉体,淀粉体形状有较明显的差异,前者多呈纺锤形或不规则形状,后者多呈球形(图2B、图2D);花后15 d,淀粉体继续充实,体积变大、数量增加,扬麦16胚乳细胞中淀粉体数明显多于宁麦13,而且蛋白体数目明显比宁麦13多、体积大(图2E、图2H);花后20 d,两品种细胞核被淀粉体和蛋白体挤压变形,开始发生降解;花后25 d,两个品种的中央胚乳细胞基本被淀粉体和蛋白体填充,且扬麦16充实更加紧密(图2G、图2J)。此外,从花后15、20和25 d的切片可以看出,扬麦16 胚乳细胞内小淀粉体出现时间早,而宁麦13小淀粉体出现时间晚(图2F、图2G、图2I、图2J)。
近糊粉层胚乳细胞显微观察结果与中央胚乳细胞一致。对花后15和25 d的胚乳细胞淀粉体和蛋白体进行统计分析后发现,扬麦16胚乳细胞中淀粉和蛋白质的积累量显著高于宁麦13,而且扬麦16单个淀粉粒的平均面积和长度均大于宁麦13(表1)。综上所述,扬麦16胚乳细胞中淀粉体和蛋白体数量多、体积增长快,反映出淀粉和蛋白质充实速率高,籽粒灌浆速度快。

M:成熟期 Maturity.
2.3 颖果养分运输组织的发育
2.3.1 母体养分运输组织的发育
花后15 d,颖果背部维x管束发育成熟,具有较强的养分输送能力。此时的灌浆速率和千粒重增加值也最大(图1A、图1B)。对花后15 d的颖果背部维管束切片,显微结构观察结果表明,扬麦16木质部维管薄壁细胞和中间细胞的原生质较浓厚,中央大液泡不明显,导管与周围细胞间的空隙较大,说明质外体运输空间较丰富(图3);而韧皮部维管薄壁细胞被挤压,形状发生改变,原生质不明显,细胞核开始发生形变解体;最显著的差异是木质部导管和韧皮部筛管的周围细胞内有较多的淀粉粒(图3A、图3C)。宁麦13木质部维管薄壁细胞的液泡明显,原生质向四周压缩,细胞内淀粉粒数量少;韧皮部细胞也开始发生形变,细胞核开始解体,淀粉粒的大小和数量都小于扬麦16(图3B、图3D)。统计数据显示,扬麦16导管和筛管无论是单个导/筛管细胞面积还是总横截面积均明显大于宁麦13(表2),从结构上保证了水和灌浆物质的运输通道更发达,有较为明显的优势。

A~B及E~G分别为扬麦16花后5、10、15、20、25 d的中央胚乳细胞;C~D及H~J分别为宁麦13花后5、10、15、20、25 d的中央胚乳细胞;Nu:细胞核;PB:蛋白体;SG:淀粉粒;标尺为20 μm。
A-B and E-G represent the center endosperm cells of Yangmai 16 at 5,10,15,20,25 days after anthesis,respectively;C-D and H-J represent the center endosperm cells of Ningmai 13 at 5,10,15,20,25 days after anthesis,respectively;Nu:Cell nucleus;PB:Protein body;SG:Starch granule;Bar=20 μm.
图2 小麦颖果中央胚乳细胞的显微结构
Fig.2 Microstructure of center endosperm cells in wheat caryopsis
合点位于维管束和珠心突起之间,在养分向胚乳的运输过程中起重要作用。研究发现,扬麦16和宁麦13的合点发育状况有较明显差异(图4)。花后5 d,两者合点细胞的细胞核和细胞质还未消亡;花后10 d,两者细胞核数量均减少;花后15 d,扬麦16细胞核明显消亡,细胞质变得不明显,养分运输的阻力降低。宁麦13合点液泡单宁酸积累早、数量多,花后10 d即可观察到因积累单宁酸而边缘染成深色的液泡(图4D),这些物质会阻碍灌浆物质运输;而扬麦16花后10 d未见单宁酸积累。花后20和25 d的切片显示,宁麦13合点细胞中积累单宁酸的液泡数量和大小均远高于扬麦16。
珠心突起细胞从紧邻合点区域一直延伸至胚乳腔中,在灌浆期会分化出珠心突起传递细胞,以便于将养分从合点运输卸载至胚乳腔中。本文中将靠近合点的部分称为上部珠心突起,靠近胚乳腔的称为下部珠心突起。研究表明,扬麦16和宁麦13珠心突起传递细胞差异非常明显(图5)。花后5 d,大约有一层下部珠心突起细胞的壁加厚,分化成珠心突起传递细胞,可观察到扬麦16的珠心突起传递细胞形状较规则,排列更为紧密(图5A、图5C);花后10 d,珠心突起传递细胞壁进一步加厚,扬麦16的壁加厚更明显(图5B、图5D);花后15 d,下部珠心突起的多层细胞分化成传递细胞,细胞壁明显增厚且向内突起生长形成壁内突。而且从下部到上部,珠心突起传递细胞的壁逐渐变薄;扬麦16的细胞质和细胞核大量减少,宁麦13仍有较多的细胞核尚未消失(图5E、图5H);花后20 d,绝大部分珠心突起细胞分化成传递细胞,细胞壁明显增厚,细胞质和细胞核大量减少,扬麦16的细胞壁厚度显著大于宁麦13(图5F、图5I);花后25 d,细胞壁变得非常厚,扬麦16大部分细胞核和细胞质几乎消失,珠心突起传递细胞的层数、壁厚和壁内突的数量均显著大于宁麦13(图5G、图5J)。上述现象说明扬麦16珠心突起传递细胞更为发达,运输养分的能力更强。

表1 发育颖果胚乳细胞中淀粉体和蛋白体密度和大小Table 1 Density and size of starch granule and protein body in endosperm cells of development caryopsis
开花后15 d数据采集自光学显微镜下200倍放大图像,开花后25 d数据采集自光学显微镜下1 000倍放大图像,因此出现同一视野内密度降低的现象。
The data of 15 days after anthesis was acquired by 200 times magnification under optical microscope,and the data of 25 days after anthesis was acquired by 1 000 times magnification,and thus the density reduced in this table which showed in the same vision.
A:扬麦16的木质部和导管; B:宁麦13的木质部和导管; C:扬麦16的韧皮部和筛管; D:宁麦13的韧皮部和筛管;标尺为20 μm; ST:筛管; VE:导管; VP:维管薄壁细胞。
A:The xylem and vessel of Yangmai 16; B:The xylem and vessel of Ningmai 13; C:The phloem and sieve tube of Yangmai 16; D:The phloem and sieve tube of Ningmai 13; Bar=20 μm; ST:Sieve tube; VE:Vessel; VP:Vascular parenchyma cell.
图3 小麦颖果背部维管束的显微结构
Fig.3 Microstructure of dorsal vascular bundle in wheat caryopsis

表2 花后15 d扬麦16与宁麦13导管和筛管的面积及内径Table 2 Area and internal diameter of the vessels and sieve tubes of Yangmai 16 and Ningmai 13 at 15 days after anthesis
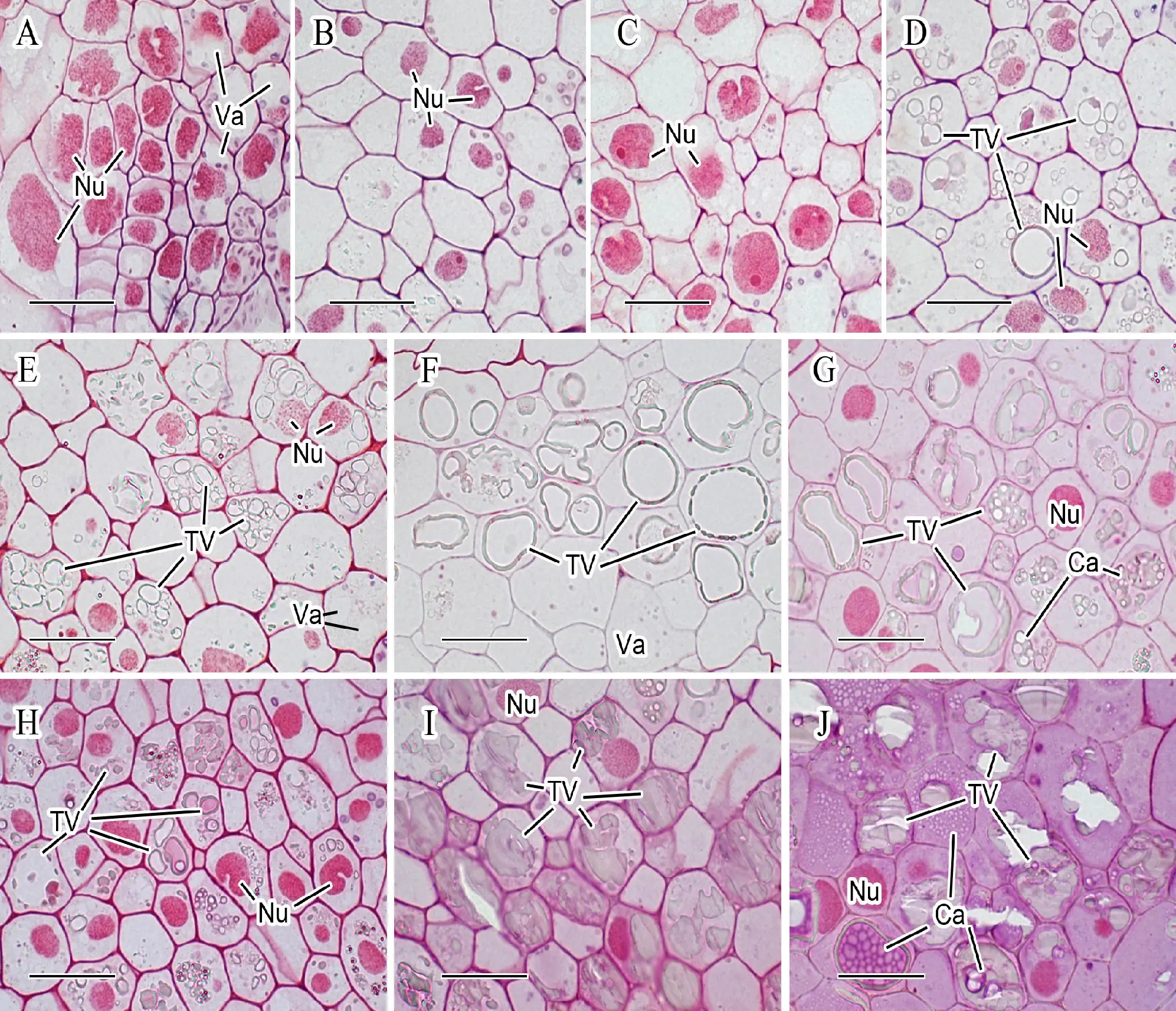
A~B及E~G分别为扬麦16花后5、10、15、20和25 d的合点区细胞;C~D及H~J分别为宁麦13花后5、10、15、20和25 d的合点区细胞;标尺为20 μm;Ca:胼胝质;Nu:细胞核;TV:含单宁酸的液泡;Va:液泡。
A-B and E-G represent the chalazal cell of Yangmai 16 at 5,10,15,20,25 days after anthesis,respectively;C-D and H-J represent the chalazal cells of Ningmai 13 at 5,10,15,20,25 days after anthesis,respectively;Bar=20 μm;Ca:Callose;Nu:Cell nucleus;TV:Tannin vacuole;Va:Vacuole.
图4 小麦颖果合点区细胞的显微结构
Fig.4 Microstructure of chalazal cells in wheat caryopsis

A~B及E~G分别为扬麦16花后5、10、15、20和25 d的珠心突起传递细胞;C~D及H~J分别为宁麦13花后5、10、15、20和25 d的珠心突起传递细胞;标尺为20 μm;EC:胚乳腔;NP:珠心突起;NPTC:珠心突起传递细胞;Nu:细胞核;TW:加厚的细胞壁。
A-B and E-G represent the nucellar projection transfer cells of Yangmai 16 at 5,10,15,20,25 days after anthesis,respectively;C-D and H-J represent the nucellar projection transfer cells of Ningmai 13 at 5,10,15,20,25 days after anthesis,respectively;Bar=20 μm;EC:Endosperm cavity;NP:Nucellar projection;NPTC:Nucellar projection transfer cells;Nu:Cell nucleus;TW:Thickened cell wall.
图5 小麦颖果珠心突起传递细胞的显微结构
Fig.5 Microstructure of nucellar projection transfer cells in wheat caryopsis
2.3.2 子代养分运输组织的发育
养分由珠心突起传递细胞卸载到胚乳腔后,转而通过属于子代养分运输组织的糊粉层传递细胞和内胚乳传递细胞输入内胚乳组织进行贮藏或满足发育需要。花后5 d,沿胚乳腔的胚乳表层细胞胞质较浓稠,开始向糊粉层传递细胞分化(图6A、图6C);花后10 d,某些相邻的糊粉层传递细胞在紧邻胚乳腔处开始分开,出现胞间隙(图6B、图6D),并随着颖果发育逐渐增大;花后15 d,扬麦16的细胞壁开始明显加厚,细胞壁厚度大、壁内突更明显(图6E至6J)。小麦内胚乳传递细胞中会有小淀粉体形成。这些淀粉体明显比胚乳中的淀粉体小,而且形状较一致,多为圆球形或椭圆形(图7)。从内胚乳传递细胞的发育可以看出,扬麦16细胞壁厚度也明显大于宁麦13(图7E至7J),细胞内淀粉粒的积累量也多于宁麦13(图7G、图7J),说明扬麦16灌浆快,细胞发育进程短。
糊粉层传递细胞由1~2层细胞组成,与之紧邻的1~2层细胞分化为内胚乳传递细胞,这两部分共同构成了小麦的胚乳传递细胞带,其在颖果发育早期就分化形成(图6A、图6C、图7B、图7D),为后来的灌浆物质大量运输入胚乳做好了结构保障,其发育的状况和特点也从而影响着灌浆速度。与宁麦13相比,扬麦16还形成了数量更多的细胞壁褶皱和壁内突;加厚的细胞壁与壁内突之间还形成了较大的质外体空间(图6E至6G、图7E至7F),扩大了养分的质外体运输渠道。

A~B及E~G分别为扬麦16花后5、10、15、20和25 d的糊粉层传递细胞;C~D及H~J分别为宁麦13花后5、10、15、20和25 d的糊粉层传递细胞;标尺为20 μm;箭头指示细胞壁内突;ATC:糊粉层传递细胞;ETC:胚乳传递细胞;Nu:细胞核。
A-B and E-G represent the aleurone transfer cells of Yangmai 16 at 5,10,15,20,25 days after anthesis,respectively;C-D and H-J represent the aleurone transfer cells of Ningmai 13 at 5,10,15,20,25 days after anthesis,respectively;Bar=20 μm;The wall ingrowths are indicated by arrow heads;ATC:Aleurone transfer cells;ETC:Endosperm transfer cells;Nu:Cell nucleus.
图6 小麦颖果糊粉层传递细胞的显微结构
Fig.6 Microstructure of aleurone transfer cells in wheat caryopsis
3 讨 论
3.1 扬麦16快速灌浆特性与颖果发育特征
本研究表明,花后15~25 d是扬麦16粒重增加最快的时期,该时期胚乳细胞内的淀粉体、蛋白体数量和体积的增加最快,胚乳细胞生长是颖果体积和粒重增大的主要原因。郭文善等[22,26]研究认为,胚乳细胞的增殖开始于花后8 d,花后15 d数目基本稳定;荆彦平等[4,27]研究表明,胚乳细胞从建成后就不断生长,在花后16~24 d生长速度最快。这与本文结论一致。
前人研究表明,IAA对有机物质向籽粒的转运有一定的促进作用,IAA 含量与蔗糖转化酶和蔗糖合成酶的活性呈极显著正相关,IAA 含量较高的基因型籽粒的充实速率也高[28];适量浓度的IAA能提高淀粉分枝酶活性,从而促进支链淀粉的合成[29];李春燕等[30]研究也表明,IAA对淀粉积累速率有极显著的促进作用。灌浆期扬麦16颖果的内源生长素IAA的含量较高,促进光合产物向颖果中的运输和积累,这是扬麦16灌浆快的原因之一。

A~B及E~G分别为扬麦16花后5、10、15、20和25 d的内胚乳传递细胞;C~D及H~J分别为宁麦13花后5、10、15、20和25 d的内胚乳传递细胞;标尺为20 μm;图E中箭头指示的为胞间连丝孔道;图F中指示的为细胞壁内突;ATC:糊粉层传递细胞;ETC:胚乳传递细胞;Nu:细胞核;SG:淀粉粒;Va:液泡。
A-B and E-G represent the starchy endosperm transfer cells of Yangmai 16 at 5,10,15,20,25 days after anthesis,respectively;C-D and H-J represent the starchy endosperm transfer cells of Ningmai 13 at 5,10,15,20,25 days after anthesis,respectively;The pore passage suspected to be plasmodesmata is indicated by the arrow head in E; The wall ingrowths is indicated by arrow head in F;Bar=20 μm;ATC:Aleurone transfer cells;ETC:Endosperm transfer cells;Nu:Cell nucleus;SG:Starch granule;Va:Vacuole.
图7 小麦颖果内胚乳传递细胞的显微结构
Fig.7 Microstructure of starchy endosperm transfer cells in wheat caryopsis
扬麦16的千粒重始终高于宁麦13,单个籽粒体积大、库容大。显微照片显示,灌浆期扬麦16的筛管管径大、传递细胞发达,养分运输组织发育与灌浆物质的“流”相适应。徐云姬等[31]研究表明,在相同施氮量下,扬麦16的SPAD值和光合速率均高于宁麦13,说明扬麦16的光合作用能力更强,提供了充足的同化物来源,“源强”则有利于同化物向籽粒中转运。综上所述,扬麦16的具有源强、库大、流畅的特点,我们推测其“源-库-流”较协调[32],因此灌浆快、粒重高。
3.2 扬麦16快速灌浆与颖果养分运输组织结构特征的关系
灌浆速率指单位时间内籽粒的干物质增加量,主要受遗传控制[17]。灌浆持续期恒定的情况下灌浆速率越大越有利于获得高产。作为灌浆物质进入颖果的唯一通道,背部维管束对籽粒灌浆和发育具有重要作用[33]。扬麦16颖果背部维管束发达,韧皮部分布范围广,观察到的筛管数量多、管径大,这使得同化物向周围组织卸载的路径和方向更多、通过共质体运输的接触面更广,避免了局部蔗糖为主的碳水化合物浓度过高而降低卸载速率。
合点是维管束和胚乳之间养分运输的桥梁,在颖果养分运输过程中具有十分重要的意义[23]。花后颖果发育过程中宁麦13的千粒重始终小于扬麦16,这一方面是其籽粒较小,另一方面可能是单宁酸等物质阻碍了灌浆物质的流入和运输[23]。扬麦16颖果发育后期合点细胞液泡较完整,推测其释放出的单宁酸少,对物质运输的阻力较小,可能是其“灌浆快”的主要原因之一;另外细胞内含物的消失也能减少养分运输的阻力[8,24]。
由韧皮部筛管卸载的灌浆物质以共质体途径或质外体途径经过合点细胞后,便运到珠心突起。紧邻胚乳腔的珠心突起表层细胞在灌浆物质中糖信号刺激下,逐渐分化成珠心突起传递细胞。Wang等[34]证明珠心突起传递细胞质膜表面积的增加与颖果干物质积累正相关。我们观察到越靠近胚乳腔,珠心突起传递细胞的壁内突越发达,且细胞质等内含物逐渐消失。扬麦16珠心突起传递细胞的细胞壁增厚早于宁麦13,壁内突更发达、数量更多,细胞排列紧密规则,细胞内含物少。这种结构特点促使由维管束卸载的绝大部分养分都被高效率的转运到胚乳腔中[13,35]。与珠心突起传递细胞隔着胚乳腔正对的是胚乳组织,紧邻胚乳腔的2~3层胚乳表层细胞依次分化为糊粉层传递细胞和内胚乳传递细胞,是胚乳腔中的养分快速输入胚乳贮藏细胞的关键通道[12,15,36]。研究表明,扬麦16的3种传递细胞都有壁内突数量更多、密度更大的特点,使跨膜的共质体运输速度得到提升,而且糊粉层传递细胞间的空隙使质外体空间深入到内层胚乳传递细胞,又增大了养分从质外体进入传递细胞内部的接触面和通道。因此,扬麦16颖果内养分运输组织的结构特征与其养分贮藏组织的快速发育相适应,是灌浆速度快、粒重高的结构学基础。
4 结 论
扬麦16花后千粒重增加快,胚乳细胞内淀粉体和蛋白体较同时期宁麦13发育好,淀粉和蛋白质等充实快;养分运输组织发达适应其“灌浆快”的特点:维管束发达、韧皮部卸载同化物能力强;合点细胞单宁酸等阻滞物质生成时间晚、含量低,细胞内含物消失早,有利于充分发挥运输通道功能;珠心突起传递细胞、糊粉层传递细胞和内胚乳传递细胞等分化早、发育状况好,细胞壁内突等养分运输的特异性结构发达,养分转运能力强;灌浆过程中颖果内源生长素水平高,促进了植株同化物向籽粒的调运。
[1] 李世清,邵明安,李紫燕,等.小麦籽粒灌浆特征及影响因素的研究进展[J].西北植物学报,2002,23(11):2031.LI S Q,SHAO M A,LI Z Y,etal.Review of characteristics of wheat grain fill and factors to influence it [J].ActaBotanicaBoreali-OccidentaliaSinica,2002,23(11):2031.
[2]SABELLI P A,LARKINS B A.The development of endosperm in grasses [J].PlantPhysiology,2009,149(1):14.
[3]BECRAFT P W.Cell fate specification in the cereal endosperm [J].SeminarsinCell&DevelopmentalBiology,2001,12(5):387.
[4] 荆彦平,刘大同,李栋梁,等.小麦内胚乳细胞及其淀粉体的发育[J].麦类作物学报,2013,33(4):818.JING Y P,LIU D T,LI D L,etal.Development of starch endosperm cells and amyloplasts in wheat [J].JournalofTriticeaeCrops,2013,33(4):818.
[5]BECRAFT P W,YI G.Regulation of aleurone development in cereal grains [J].JournalofExperimentalBotany,2011,62(5):1669.
[6]GARCIA D,GERALD J N F,BERGER F.Maternal control of integument cell elongation and zygotic control of endosperm growth are coordinated to determine seed size inArabidopsis[J].ThePlantCell,2005,17(1):52.
[7]ZHENG Y,WANG Z,GU Y.Development and function of caryopsis transport tissues in maize,sorghum and wheat [J].PlantCellReports,2014,33(7):1023.
[8]RADCHUK V,WEIER D,RADCHUK R,etal.Development of maternal seed tissue in barley is mediated by regulated cell expansion and cell disintegration and coordinated with endosperm growth [J].JournalofExperimentalBotany,2011,62(3):1217.
[9]COCHRANE M P,PATERSON L,GOULD E.Changes in chalazal cell walls and in the peroxidase enzymes of the crease region during grain development in barley [J].JournalofExperimentalBotany,2000,51(344):507.
[10]GUNNING B E S,PATE J S.Transfer cells:Plant cells with wall ingrowths,specialized in relation to short distance transport of solutes-their occurrence,structure,and development [J].Protoplasma,1969,68(1-2):107.
[11]OFFLER C E,MCCURDY D W,PATRICK J W,etal.Transfer cells:Cells specialized for a special purpose [J].AnnualReviewofPlantBiology,2003,54(1):431.
[12]THOMPSON R D,HUEROS G,BECKER H A,etal.Development and function of seed transfer cells [J].PlantScience,2001,160(5):775.
[13]ZHENG Y,WANG Z.Contrast observation and investigation of wheat endosperm transfer cells and nucellar projection transfer cells [J].PlantCellReports,2011,30(7):1281.
[14] 王慧慧,王 峰,刘大同,等.小麦胚乳传递细胞发育的结构观察[J].麦类作物学报,2011,31(5):944.WANG H H,WANG F,LIU D T,etal.Structure and development of endosperm transfer cells in wheat [J].JournalofTriticeaeCrops,2011,31:944.
[15] 郑彦坤.关于玉米、高粱和小麦的颖果养分运输组织发育的结构观察[D].扬州:扬州大学,2012:90-91.ZHENG Y K.Structure observation of caryopsis nutrient transport tissues in maize,sorghum and wheat [D].Yangzhou:Yangzhou University,2012:90-91.
[16] 王 慧,朱冬梅,王君婵,等.扬麦16耐迟播早熟特性研究[J].麦类作物学报,2016,36(12):1657.WANG H,ZHU D M,WANG J C,etal.Study on the early maturing characteristics of Yangmai 16 under late planting [J].JournalofTriticeaeCrops,2016,36(12):1657.
[17] 任明全,徐向阳.不同小麦品种籽粒灌浆特性的研究[J].华北农学报,1993,8(3):28.REN M Q,XU X Y.Studies on the grain filling characters of wheat cultivars [J].ActaAgriculturaeBorealiSinica,1993,8(3):28.
[18] 蔡庆生,吴兆苏.小麦籽粒生长各阶段干物质积累量与粒重的关系[J].南京农业大学学报,1993,16(1):27.CAI Q S,WU Z S.The relations of dry matter accumulation of grain growth stages to grain weight in wheat [J].JournalofNanjingAgriculturalUniversity,1993,16(1):27.
[19] 周竹青,朱旭彤.不同粒重小麦品种(系)籽粒灌浆特性分析[J].华中农业大学学报,1999,18(2):107.ZHOU Z Q,ZHU X T.Analysis on the grain filling characteristics of wheat varieties with different kernel weight [J].JournalofHuazhongAgriculturalUniversity,1999,18(2):107.
[20] 吴少辉,高海涛,张学品,等.播期对不同习性小麦品种籽粒灌浆特性的影响[J].麦类作物学报,2004,24(4):105.WU S H,GAO H T,ZHANG X P,etal.Effect of sowing date on grain filling characters in different wheat varieties [J].JournalofTriticeaeCrops,2004,24(4):105.
[21] 冯素伟,胡铁柱,李 淦,等.不同小麦品种籽粒灌浆特性分析[J].麦类作物学报,2009,29(4):643.FENG S W,HU T Z,LI G,etal.Analysis on grain filling characteristics of different wheat varieties [J].JournalofTriticeaeCrops,2009,29(4):643.
[22] 郭文善,方明奎,王蔚华,等.小麦籽粒胚乳细胞发育的生理特性[J].扬州大学学报(自然科学版),1998,1(2):41.GUO W S,FANG M K,WANG W H,etal.Physiological characteristics of grain endosperm cells in wheat [J].JournalofYangzhouUniversity(NaturalScienceEdition),1998,1(2):41.
[23]COCHRANE M P.Morphology of the crease region in relation to assimilate uptake and water loss during caryopsis development in barley and wheat [J].AustralianJournalofPlantPhysiology,1983,10(6):473.
[24]KLADNIK A,CHAMUSCO K,DERMASTIA M,etal.Evidence of programmed cell death in post-phloem transport cells of the maternal pedicel tissue in developing caryopsis fomazie [J].PlantPhysiology,2004,136(3):3572.
[25]YANG J,ZHANG J,WANG Z,etal.Hormonal changes in the grains of rice subjected to water stress during grain filling [J].PlantPhysiology,2001,127(1):315.
[26] 王蔚华,郭文善,方明奎,等.小麦籽粒胚乳细胞增殖及物质充实动态[J].作物学报,2003,29(5):779.WANG W H,GUO W S,FANG M K,etal.Endosperm cell proliferating and grain filling dynamics in wheat [J].ActaAgronomicaSinica,2003,29(5):779.
[27]JING Y P,LIU D T,YU X R,etal.Development of endosperm cells and starch granules in common wheat [J].CerealResearchCommunications,2014,42(3):514.
[28] 刘仲齐,吴兆苏,俞世蓉.吲哚乙酸和脱落酸对小麦籽粒淀粉积累的影响[J].南京农业大学学报,1992,15(1):7.LIU Z Q,WU Z S,YU S R.Effects of IAA and ABA on starch accumulation in wheat grain [J].JournalofNanjingAgriculturalUniversity,1992,15(1):7.
[29] 潘晓华,李木英,熊 伟,等.蔗糖和谷氨酰胺及植物激素对水稻离体培养穗淀粉积累的影响[J].江西农业大学学报,2000,22(1):1.PAN X H,LI M Y,XIONG W,etal.Effects of sucrose,glutamine and plant hormones in the culture medium on starch accumulation of detached rice panicles [J].ActaAgriculturaeUniversitatisJiangxiensis,2000,22(1):1.
[30] 李春燕,封超年,张 容,等.宁麦9号花后内源激素和蔗糖含量变化及其与籽粒淀粉合成的关系[J].麦类作物学报,2007,27(1):138.LI C Y,FENG C N,ZHANG R,etal.Relationship of grain starch synthesis with changes of endogenous hormone and sucrose content after anthesis in wheat variety Ningmai 9 [J].JournalofTriticeaeCrops,2007,27(1):138.
[31] 徐云姬,张伟杨,钱希旸,等.施氮量对小麦籽粒灌浆的影响及其生理机制[J].麦类作物学报,2015,35(8):1119.XU Y J,ZHANG W Y,QIAN X Y,etal.Effect of nitrogen on grain filling of wheat and its physiological mechanism [J].JournalofTriticeaeCrops,2015,35(8):1119.
[32] 王 忠.植物生理学(第2版) [M].北京:中国农业出版社,2009:238-272.WANG Z.Plant Physiology(2nd Edition) [M].Beijing:China Agriculture Press,2009:238-272.
[33] 王 忠,顾蕴洁,李卫芳,等.小麦胚乳发育及其养分输人的途径[J].作物学报,1998,24(5):536.WANG Z,GU Y J,LI W F,etal.Development of wheat endosperm and the pathway of nutrients entering the endosperm [J].ActaAgronomicaSinica,1998,24(5):536.
[34]WANG H L,OFFLER C E,PATRICK J W.The cellular pathway of photosynthate transfer in the developing wheat grain.II.A structural analysis and histochemical studies of the pathway from the crease phloem to the endosperm cavity [J].Plant,Cell&Environment,1995,18(4):373.
[35]WANG H L,OFFLER C E,PATRICK J W.Nucellar projection transfer cells in the developing wheat grain [J].Protoplasma,1994,182(1-2):39.
[36]ZHENG Y,WANG Z,YANG J,etal.Observation and comparison of structure changes in wheat caryopsis maternal tissues and endosperm [J].BrazilianJournalofBotany,2015,38(2):417.
Relationship between Caryopsis Microstructure and Endogenous Auxin Level and the Fast Grain Filling of Yangmai 16
LIU Datong1,YU Xurun2,ZHU Dongmei1,LIU Jian1,ZHANG Xiao1,GAO Derong1,2
(1.Lixiahe Regional Institute of Agricultural Sciences of Jiangsu/Key Laboratory of Wheat Biology and Genetic Breeding in the Middle and Lower Yangtze River,Ministry of Agriculture,Yangzhou,Jiangsu 225007,China; 2.Co-Innovation Center for Modern Production Technology of Grain Crops,Yangzhou University,Yangzhou,Jiangsu 225009,China)
Yangmai 16,with the largest planting area in the middle and lower Yangtze river,has features such as high speed of grain filling and high grain weight. In order to elucidate the relationship between rapid grain filling and the microstructure of caryopsis and the endogenous auxin,with Ningmai 13 as control,the systematic development and microstructural changes of the vascular cells,endosperm cells and transfer cells of Yangmai 16 caryopsis were observed. The thousand kernel weight and endogenous auxin(IAA) contents were studied. The main results indicated that,the starch granule and protein body were better developed in Yangmai 16,thus the endosperm enriched well and the final grain weight was higher. The endogenous auxin level in caryopsis of Yangmai 16 was higher during grain filling,which promoted the allocation and transportation of assimilates to caryopsis and the division of the endosperm cells. In Yangmai 16,the vascular bundles,especially the sieve tube,were significantly developed,and thus the phloem unloading ability of assimilate was stronger. The differentiation of transfer cells in Yangmai 16 was earlier,and the abundant cell wall ingrowth were conducive to give full play to transport function.
Wheat; Grain filling; Grain weight; Microstructure of caryopsis; Endogenous auxin
10.7606/j.issn.1009-1041.2017.06.04
时间:2017-06-07
2017-01-24
2017-04-18
国家现代农业产业技术体系建设专项(CARS-3-2-11);江苏里下河地区农业科学研究所科研基金项目;江苏省科技支撑计划项目(BE2014351);江苏省农业科技自主创新资金项目(CX14-2002)
E-mail:15062799250@163.com
高德荣(E-mail:gdr@wheat.org.cn)
S512.1;S311
A
1009-1041(2017)06-0739-11
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170607.1004.008.html
