改变碳输入对南亚热带海岸沙地典型天然次生林土壤呼吸的影响
2017-06-06高伟叶功富黄志群陈智勇余再鹏岳新建陈梦瑶
高伟,叶功富,黄志群,陈智勇,余再鹏,岳新建,陈梦瑶
(1.湿润亚热带山地生态国家重点实验室培育基地,350007,福州;2.福建师范大学地理研究所,350007 ,福州; 3.湿润亚热带生态地理过程教育部重点实验室,350007,福州;4.福建省林业科学研究院,350012,福州; 5.福建农林大学林学院,350001,福州)
改变碳输入对南亚热带海岸沙地典型天然次生林土壤呼吸的影响
高伟1,2,4,叶功富4,黄志群1,2,3†,陈智勇5,余再鹏1,2,3,岳新建5,陈梦瑶5
(1.湿润亚热带山地生态国家重点实验室培育基地,350007,福州;2.福建师范大学地理研究所,350007 ,福州; 3.湿润亚热带生态地理过程教育部重点实验室,350007,福州;4.福建省林业科学研究院,350012,福州; 5.福建农林大学林学院,350001,福州)
土壤呼吸是陆地生态系统碳循环的一个重要过程,开展环境因子和改变碳输入对土壤呼吸影响的研究具有重要意义。2015年3月—2016年2月,在南亚热带海岸沙地典型天然次生林中设置去除根系、去除凋落物、加倍凋落物和对照4种处理,采用LI-8100连续观测改变碳输入对土壤呼吸的影响。结果表明:改变碳输入没有显著影响10cm土壤温度和湿度(P>0.05);不同处理土壤呼吸速率存在明显的季节变化,表现为夏高冬低,最大值出现在5月或者6月,最小值出现在11月或12月;土壤呼吸速率的年均值为加倍凋落物>对照>去除根系>去除凋落物,不同改变碳输入方式均降低了土壤呼吸的Q10值;矿质土壤呼吸、凋落物呼吸和根系呼吸对土壤总呼吸的贡献分别为41.24%、43.29%和15.45%;不同处理土壤呼吸速率分别与土壤温度和湿度呈显著的指数和线性正相关(P<0.05),双因子模型能解释土壤呼吸变异的45%~69%;改变碳输入影响土壤可溶性有机碳和微生物生物量碳,不同处理土壤呼吸速率与可溶性有机碳和微生物生物量碳呈正相关。因此,改变碳输入引起土壤易变碳的变化进而影响土壤呼吸。
南亚热带; 沙质海岸; 天然次生林; 土壤呼吸; 碳输入;Q10值
大气中CO2浓度的升高及由其可能引发的全球气候变化已经引起世界性的关注,使陆地生态系统碳循环成为全球性的研究热点[1-3]。在全球2万5 000亿t的土壤碳库中,有1万5 500亿t为土壤有机碳(SOC),9 500亿t为土壤无机碳(SIC),土壤碳库是大气碳库(7 600亿t)的3.3倍和植物碳库(5 600亿t)的4.5倍[2]。土壤有机碳可在土壤中停留的时间超过1 000年,因此土壤可提供巨大的碳汇以抵消化石燃料燃烧而导致的CO2排放,造林是增加植被层和土壤层碳封存的有效措施[3]。作为陆地向大气输出CO2的主要途径,土壤呼吸是全球陆地生态系统碳循环的重要组成部分[4],由土壤呼吸引起的CO2释放被认为是全球碳循环的最大通量之一,土壤呼吸速率的微小波动都会显著影响大气中的CO2浓度[5]。
土壤呼吸主要包括根系呼吸、土壤微生物和土壤动物的呼吸以及含碳矿物质的化学氧化作用。凋落物是土壤呼吸地上部分的主要碳源[6],其数量和质量可以显著影响表层土壤碳底物的有效性,从而限制土壤呼吸[7]。有研究指出,植物地上部分的凋落物量与土壤呼吸密切相关[8],改变凋落物输入可以显著改变土壤的CO2通量[9]。凋落物输入增加可能引起“激发效应”从而加速土壤中固有有机碳的矿化,致使土壤碳释放增强[10]。此外,改变凋落物输入还可影响土壤表层细根的分布[11],进而改变根系分泌物以影响土壤呼吸速率。根系是将植物光合产物输送到地下土壤的主要途径,大量研究表明,去除根系可以显著降低土壤呼吸速率[6,12]。根呼吸是全球碳平衡的重要组分,其对土壤总呼吸的贡献率为5%~84%,大多数研究集中在30%~40%[13]。
南亚热带海岸沙地是我国沿海地区典型生态脆弱带,受台风、风暴潮以及旱涝交替等灾害性天气危害严重[14],以乡土树种潺槁木姜子(Litseaglutinosa(Lour.) C. B. Rob.)为建群种的天然次生林是南亚热带海岸沙地广泛分布的典型植被类型[15],其树种组成丰富,生物多样性高,具有重要的基因价值和碳汇功能,同时相对木麻黄(CasuarinaequisetifoliaForst.)等人工林,其生态系统稳定性强,对于应对区域上的生态系统波动具有重要意义。目前,关于南亚热带海岸沙地土壤呼吸的研究多集中在根系[16]和林龄[17]对木麻黄土壤呼吸的影响,对其他植被类型土壤呼吸特征的研究尚少见报道,海岸沙地上地带性植被的土壤呼吸对改变凋落物和根系等碳源输入的响应机制尚不明确。基于此,笔者研究改变土壤碳输入对海岸沙地天然次生林土壤呼吸的影响,探讨其土壤碳释放的变化规律及主要影响因子,以期为海岸沙地土壤碳库管理和科学评价其碳循环过程提供理论参考。
1 研究区概况
研究区位于福建省东山县赤山国有防护林场(E 117°24′22.17″,N 23°38′21.22″),该场位于福建省东南部,台湾海峡西岸,属于南亚热带海洋性季风气候,1月份平均气温13.1 ℃,7月份平均气温27.3 ℃,多年平均气温为20.8 ℃,冬无严寒,夏无酷暑,终年无霜冻,多年平均降水1 103.8 mm。土壤主要有砖红壤性红壤,水稻土,风沙土和盐碱土4类。海岸沙地现存植被大部分以木麻黄纯林为主,林下植被较少,植物组成简单,少部分为木麻黄与相思类、湿地松、桉树类树种的块状或带状混交林及以乡土常绿阔叶树种为建群种的天然次生林[18]。
2 材料与方法
2.1 样地设置
选择林场附近的一片天然次生林作为土壤呼吸的监测样地,该林分物种丰富,受干扰程度较小,样地内主要乔木优势树种为潺槁木姜子和朴树(CeltissinensisPers.),重要值分别为133.62和63.68,灌木种有毛果算盘子(GlochidioneriocarpumChamp. ex Benth.)、豺皮樟(LitsearotundifoliaHemsl. var. oblongifolia (Nees) Allen)、土蜜树(BrideliatomentosaBl.)、鳝藤(Anodendronaffine(Hook. et Arn.) Druce)等,主要草本为沿阶草(Ophiopogonbodinieri)和荩草(Arthraxonhispidus(Thunb.) Makino),郁闭度0.9以上。2014年12月选取林内保存较好的地段建立4个20 m×20 m的监测小区,每个小区间隔10 m缓冲带,土壤为风积沙土,土层厚度>100 cm,枯落物层3~6 cm,样地布设完毕后采集样地表层土壤进行理化性质测定(表1)。
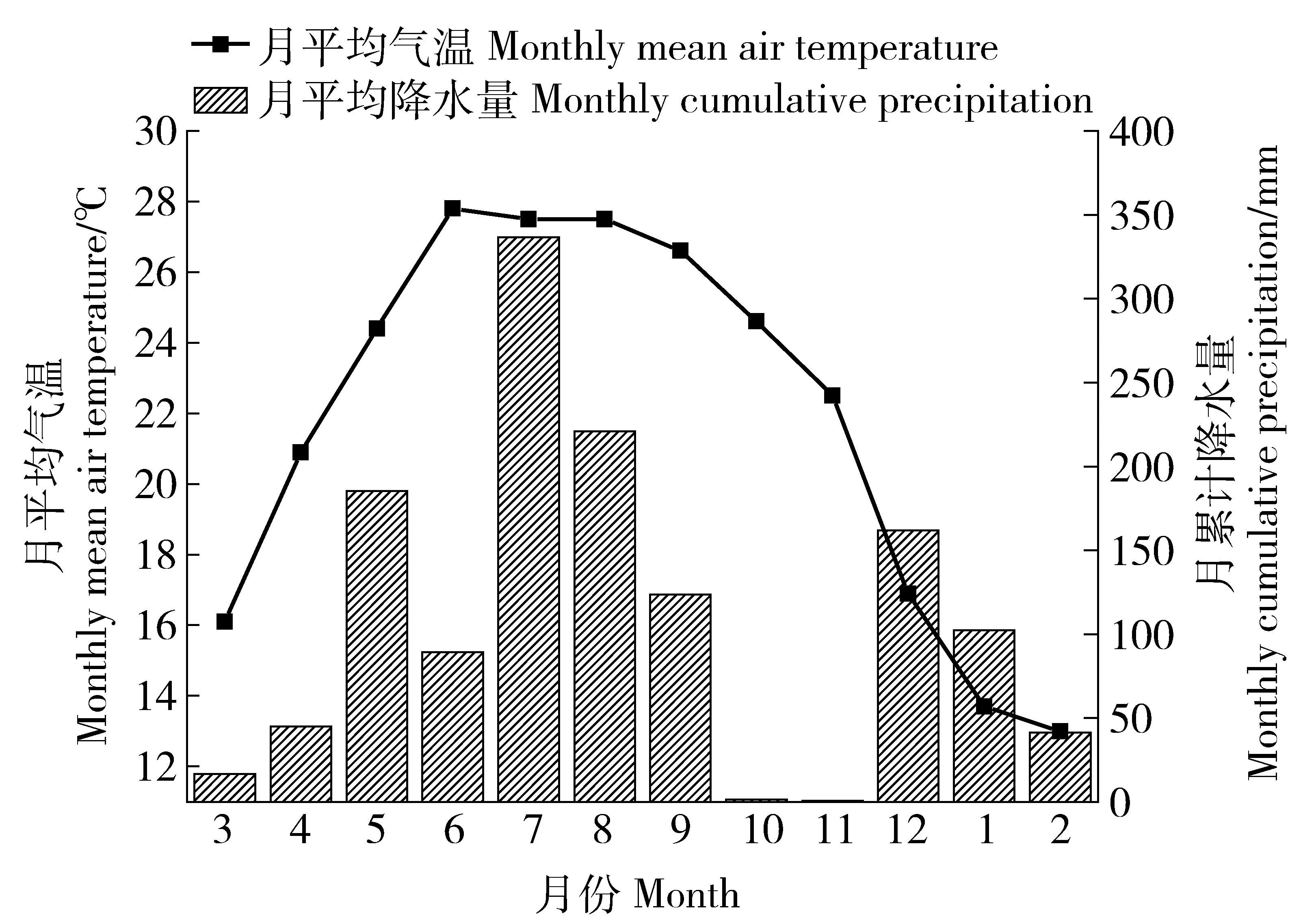
图1 研究期间平均温度和平均降水量Fig.1 Monthly mean air temperature and cumulative precipitation and during the experimental period
在每个小区中各设置4个2 m×2 m的次小区进行4种实验处理,处理1(去除根系):采用挖壕沟法切断根系,壕沟深度≥1 m,并在壕沟内垂直铺设2层100目的尼龙网阻断外围根系向次小区内生长;处理2(去除凋落物):将次小区内的地上凋落物全部移走,并在次小区上方30~50 cm处铺设一张面积为2 m×2 m,孔径为1 mm的尼龙网,阻隔凋落物;处理3(加倍凋落物):将处理2中移走的凋落物均匀的撒在本次小区内,之后每月将处理2中尼龙网的凋落物均匀撒在本次小区内;处理4(对照):不做处理。在每个次小区中间位置埋设1个内径20.0 cm、高6 cm的PVC土壤环,平放压入土中3 cm,整个测量期内土壤环位置保持不变。
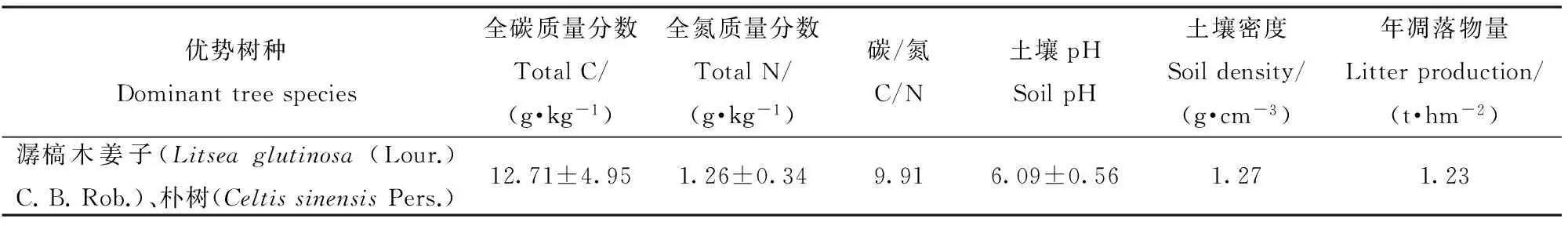
表1 天然次生林林分特征及表层(0~10 cm)土壤性质
注:表中数字为平均值±标准差。Note: Data in table are mean ± standard deviation.
2.2 土壤呼吸速率及温湿度测定
从2015年3月开始(切断根系3个月后),采用LI-8100土壤碳通量测量系统(Li-Cor Inc.,Lincoln,NE,USA)进行土壤呼吸速率的监测,每个月下旬选择晴朗无雨的天气测量土壤呼吸,测定时间为08:30—11:30[19],每个土壤环测量时间为2 min,在测定土壤呼吸的同时,采用数字瞬时温度计和时域反射仪(TDR)测定土壤环附近10 cm土壤温度和0~10 cm土壤体积含水量,并用烘干法对土壤含水率进行校正,实验于2016年2月结束,共测量12次。
2.3 土壤可溶性有机碳测定
土壤呼吸测定结束后,分别在每种处理的次小区中沿对角线选择10个点采集0~10 cm土壤,混合后带回实验室测定土壤可溶性有机碳DOC以及微生物生物量碳MBC。DOC采用冷水浸提,MBC采用氯仿熏蒸—硫酸钾浸提,不同浸提液采用总有机碳分析仪(TOC-VCPH/CPN analyzer,日本)测定有机碳含量。
2.4 数据统计分析
在SPSS 18.0中进行单因素方差分析(One-way ANOVA),在方差分析的基础上采用Duncan法多重比较检验不同处理间土壤呼吸速率、土壤温度、土壤湿度等的差异性。
Rr=R4-R1。RL=R4-R2;Rm=R1+R2-R4。
式中:Rr为根系呼吸速率;RL为凋落物呼吸速率;Rm为矿质土壤呼吸速率(包括微生物呼吸和土壤有机质分解);R1为去除根系处理的土壤呼吸速率;R2为去除凋落物处理的土壤呼吸速率;R4为对照处理土壤呼吸速率。单位均为μmol·m-2·s-1。
分别采用线性和非线性回归分析建立土壤呼吸与土壤温度、土壤湿度之间的关系模型:
Rs=aW+b;Rs=aebθ;Rs=a(θ×W)+b;
Rs=a+bθ+cW;Rs=aθbWc;Rs=aebθWc。
式中:θ为10 cm土壤温度,℃;W为10 cm土壤湿度,%;a、b、c为待定参数;土壤呼吸温度敏感系数Q10值=e10b,b为模型Rs=aebθ中的温度系数。
3 结果与分析
3.1 改变碳输入对土壤温度、湿度的影响
观测期内,不同处理间表层土壤温度均为夏高冬低,存在显著的季节变化,高温期为5—9月,9月之后,土壤温度逐渐降低,于翌年2月降到最低,之后逐渐回升。一年中,不同处理间10 cm土壤温度的平均值均为21.3 ℃左右,不同处理间土壤温度无显著差异(P>0.05,表2和图2a)。
不同处理间表层土壤湿度也存在显著的季节变化,干湿季明显。一年中,不同处理土壤含水率峰期为4—6月,最高值均出现在5月,7—11月土壤含水率较低。不同处理土壤含水率的年均值均在5%左右,不同处理间无显著差异(P>0.05,表2和图2b)。
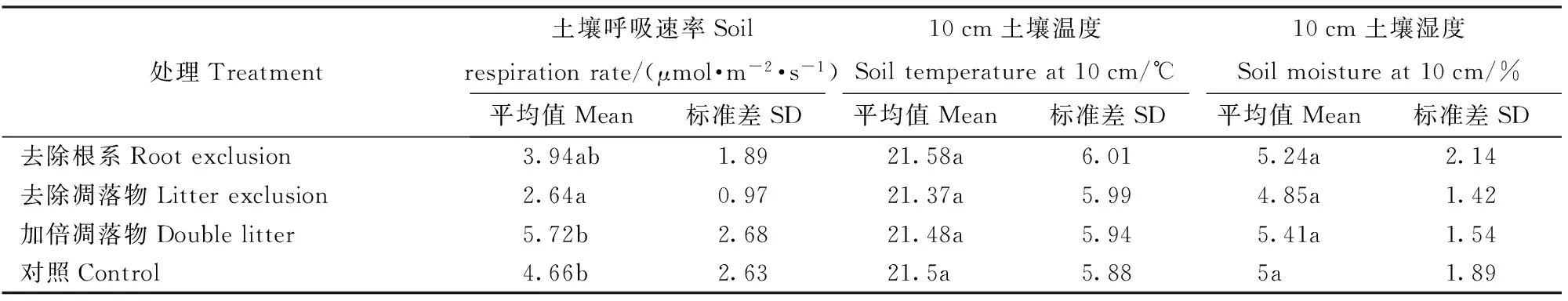
表2 不同处理10 cm土壤温度、10 cm土壤湿度及土壤呼吸速率年均值多重比较
注:表中不同字母表示在α=0.05水平差异显著。
Note: Different letters indicate significant difference among treatments(α=0.05)
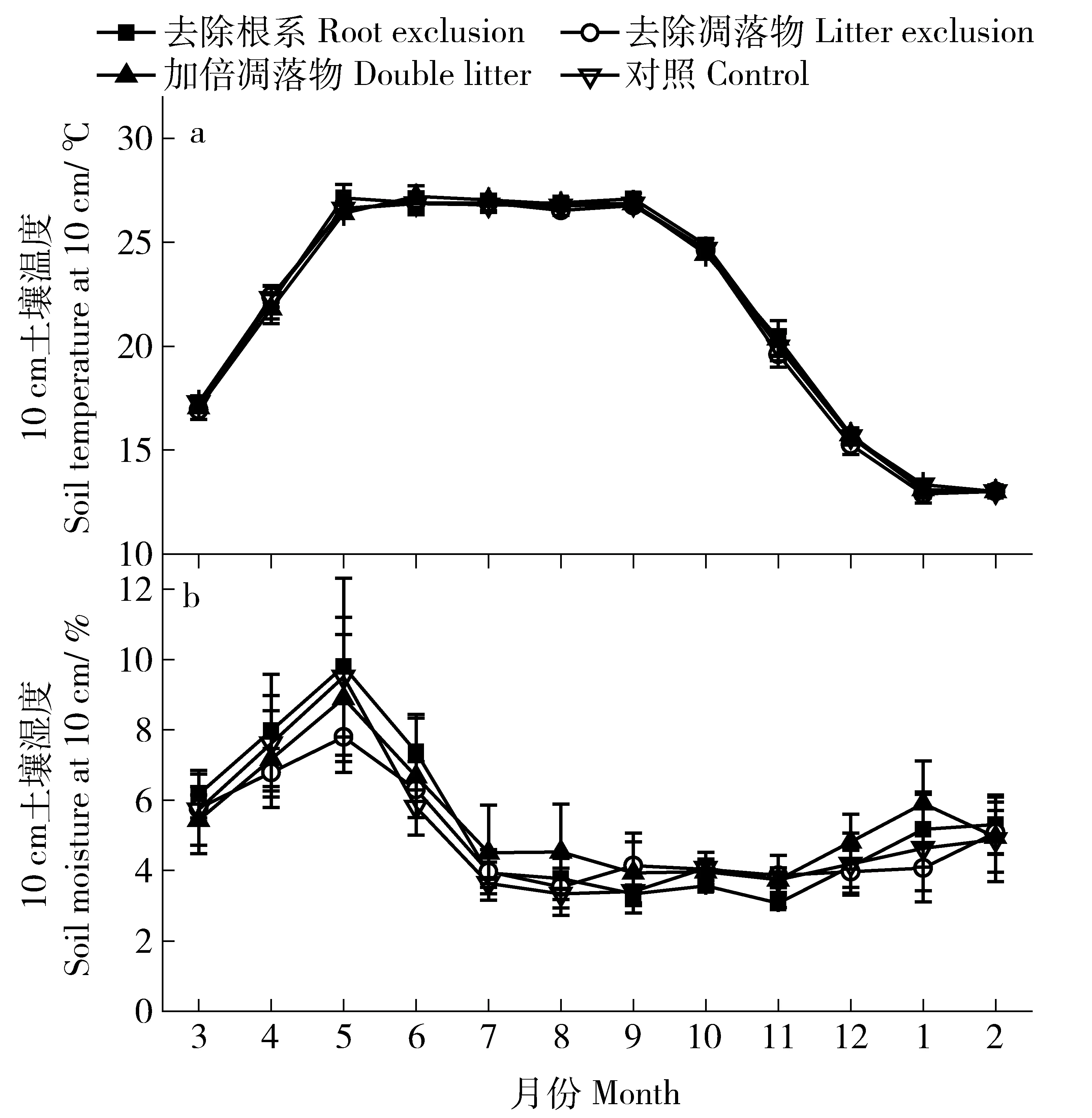
图2 不同处理10 cm土壤温度和0~10 cm土壤湿度变化动态Fig.2 Dynamics of soil temperature and soil moisture at 10 cm depth with different treatments
3.2 改变碳输入对土壤呼吸速率的影响
不同处理的土壤呼吸速率存在显著的季节变化,基本规律为夏季高、冬季低。一年中,土壤呼吸的峰值出现在5月或者6月,谷值出现在11月或12月。不同处理土壤呼吸速率的年均值为加倍凋落物(5.72±2.68 μmol·m-2·s-1)>对照(4.66±2.63 μmol·m-2·s-1)>去除根系(3.94±1.89 μmol·m-2·s-1)>去除凋落物(2.64±0.97 μmol·m-2·s-1),去除凋落物的土壤呼吸速率显著小于其他处理(P<0.05,表2和图3)。
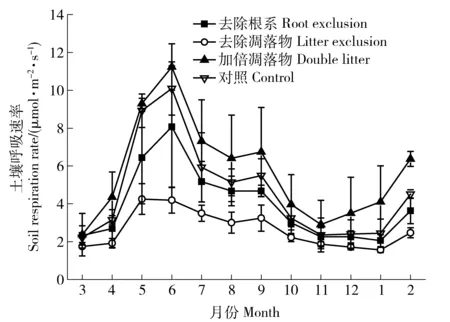
图3 天然次生林不同处理土壤呼吸速率变化动态Fig.3 Dynamic changes of soil respiration rate with differenttreatments in natural secondary forest
3.3 不同处理土壤呼吸速率变化幅度
改变碳输入显著影响土壤呼吸速率(图4)。去除根系处理的土壤呼吸速率比对照年平均降低12.27%,除首次测量土壤呼吸高于对照外,其他月份土壤呼吸速率均低于对照,变化幅度最大值出现在5月,比对照降低27.79%,最小值出现在11月,比对照降低4.24%;去除凋落物处理的土壤呼吸速率比对照年平均降低38.05%,最大值出现在6月,比对照降低58.47%,最小值出现在11月,比对照降低20.79%;加倍凋落物处理的土壤呼吸速率比对照年平均增加27.64%,最大值出现在翌年1月,比对照增加67.62%,最小值出现在5月,比对照增加4.22%。
监测期内,天然次生林矿质土壤呼吸速率、凋落物呼吸速率和根系呼吸速率的平均值分别为1.92±0.47、2.02±1.7和0.72±0.78 μmol·m-2·s-1,三者对土壤总呼吸的贡献率分别为41.24%、43.29%和15.45%。
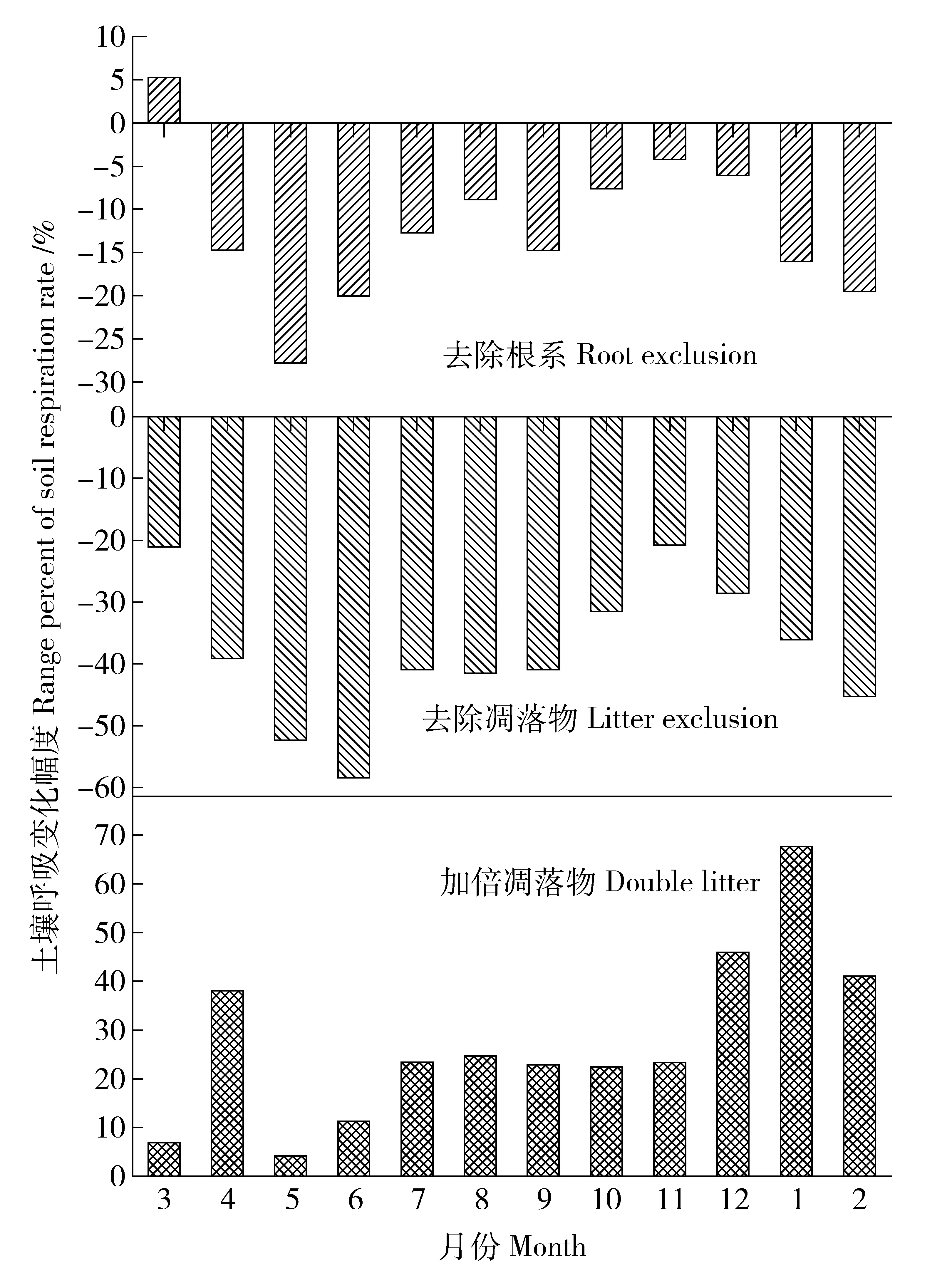
图4 天然次生林不同处理土壤呼吸速率变化幅度Fig.4 Range percent of soil respiration rate with different treatment in natural secondary forest
3.4 土壤呼吸速率与温湿度的关系
回归分析表明,不同处理的土壤呼吸速率与10 cm土壤温度之间存在显著的指数相关关系,且与土壤湿度存在显著的正相关(P<0.01,表3)。10 cm土壤湿度可解释土壤呼吸变异的19%~33%,10 cm土壤温度可解释土壤呼吸变异的20%~52%,不同处理均降低了土壤呼吸的温度敏感性(Q10),去除根系、去除凋落物、加倍凋落物和对照的Q10值分别为1.58、1.55、1.52和1.77。
采用多元线性和非线性回归方程建立土壤呼吸与土壤温度、土壤湿度之间的复合关系模型。可见,不同模型与单因子模型相比均不同程度的提高了拟合精度,双因子复合模型可解释土壤呼吸变异的45%~69%。
3.5 改变碳输入对土壤可溶性有机碳的影响
不同处理土壤可溶性有机碳(DOC)和微生物生物量碳(MBC)浓度为加倍凋落物>对照>去除根系>去除凋落物,改变碳输入均不同程度的提高或降低了DOC和MBC,不同处理间DOC浓度无显著差异(P>0.05),而加倍凋落物的MBC浓度显著高于其他处理(P<0.05)。由回归分析可见,不同处理的土壤呼吸速率与可溶性有机碳和微生物生物量碳均呈显著的正相关。
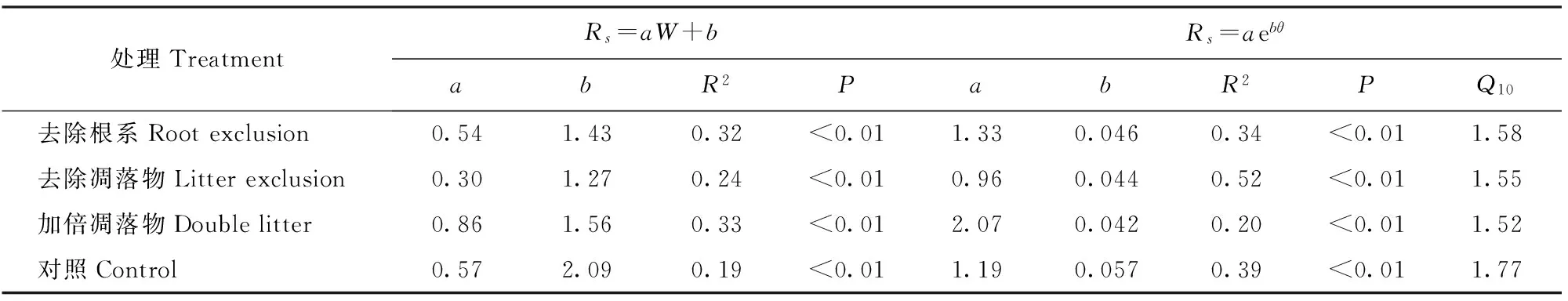
表3 土壤呼吸与10 cm土壤温度、土壤湿度的一元回归模型
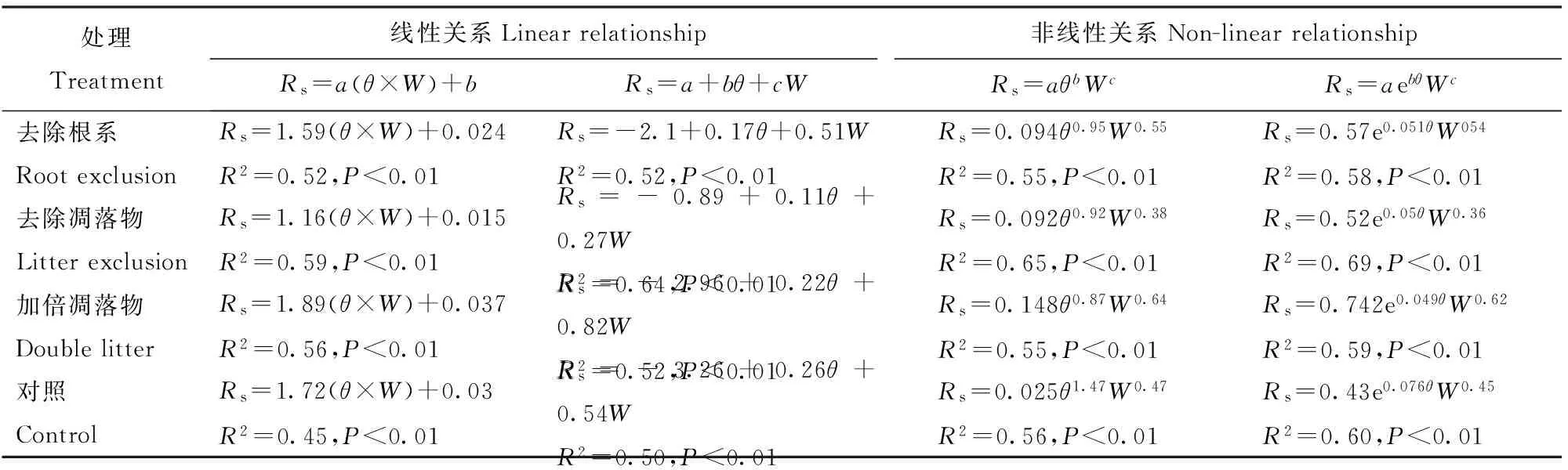
表4 土壤呼吸与10 cm土壤温度(θ)、10 cm土壤湿度(W)的复合关系模型
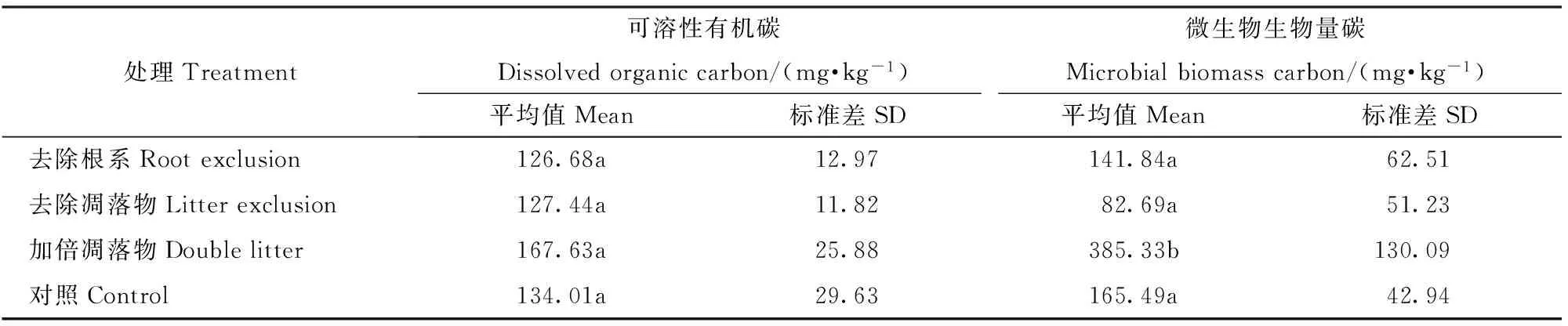
表5 不同处理土壤可溶性有机碳和微生物生物量碳
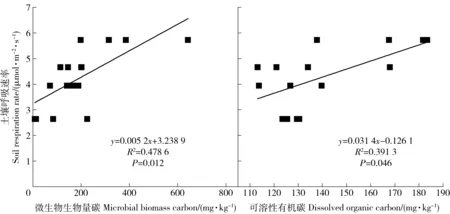
图5 不同处理平均土壤呼吸速率与微生物生物量碳和可溶性有机碳的关系Fig.5 Relationships between mean soil respiration rate and dissolved organic carbon or microbial biomass carbon with different treatments
4 讨论
4.1 凋落物与土壤呼吸的关系
在森林生态系统中,添加或去除凋落物可以改变根系和微生物生长所需要的有机碳供应,从而显著增加或降低土壤呼吸[23-29]。大量研究认为,由于激发效应的存在,加倍凋落物所引起的土壤呼吸增加幅度大于去除凋落物而引起的土壤呼吸减小幅度[20];但也有研究显示,去除和添加凋落物对土壤呼吸速率的影响并不是等同或成比例的[6],新有机碳的添加可以补偿由于激发效应而引起的固有有机碳的损耗[21]。汪金松等[6]研究发现,去除凋落物使油松人工林土壤呼吸速率降低的幅度为36.03%,而添加凋落物使土壤呼吸速率增加的幅度为21.01%。高强等[22]研究发现,加倍凋落物使木荷林土壤呼吸速率提高17.86%,而去除凋落物使土壤呼吸速率降低25.32%。本研究中,去除凋落物使土壤呼吸速率降低38.05%,加倍凋落物使土壤呼吸速率增加27.64%,即去除凋落物土壤呼吸的降低幅度大于添加凋落物土壤呼吸的增加幅度,除补偿作用外,这也可能与新添加的凋落物尚处于分解初期,土壤呼吸的激发效应尚未充分体现有关。
土壤的异氧呼吸和有机质的分解与土壤微生物的活性有重要关系,土壤微生物生长除受环境因子影响外,还受到碳源有效性的制约[23]。作为微生物生长的重要底物,凋落物数量决定了土壤微生物生物量[24],凋落物分解释放的易变碳使土壤微生物活性提高[25],去除凋落物将减少微生物生长所需要的可溶性有机碳和养分,并改变土壤微环境,引起微生物生物量和活性的降低。Hooker等[26]研究发现,添加凋落物使土壤微生物生物量碳增加13%。Feng等[27]研究发现,去除凋落物使土壤微生物量碳减少19%。本研究中,添加或去除凋落物分别使土壤DOC和MBC发生不同程度的升高或降低,且添加凋落物处理的MBC显著高于其他处理,说明改变凋落物输入影响可溶性有机碳向土壤的淋溶和土壤微生物的生长,进而影响土壤呼吸。
4.2 根系与土壤呼吸的关系
作为土壤呼吸的自养呼吸组分,根系呼吸在森林生态系统中占有重要地位,去除根系将导致土壤呼吸降低。左强等[12]研究发现,去除根系使红松阔叶混交林土壤呼吸降低36.49%。汪金松等[6]研究显示,去除根系使油松人工林土壤呼吸降低10.76%。在南亚热带海岸沙地上,去除根系使木麻黄幼龄林、中龄林和成熟林土壤呼吸分别降低28.11%、28.98%和26.47%[16]。本研究中,去除根系使海岸沙地天然次生林土壤呼吸速率降低12.27%,其降低幅度低于木麻黄人工林,这种差异可能主要与森林类型的不同有关。
根系是植物光合产物向地下输送的唯一途径,切断根系使植物地上部分光合产物向地下部分分配并通过根系分泌向土壤的碳输入受阻,可能导致根系活性和分泌物降低,从而影响土壤微生物量,进而影响土壤呼吸。Hart等[28]研究发现,采用挖壕沟去除根系13a后,土壤微生物量碳降低25%~75%。Li等[29]研究发现,去除根系7a后,土壤微生物量碳降低45.4%。也有研究认为,根系去除对土壤微生物的影响存在树种与区域差异,去除根系不一定影响土壤的微生物量[30]。本研究中,去除根系分别使土壤DOC和MBC降低5.47%和14.29%,但与对照尚无显著差异,这可能与林分类型和研究区域的不同或与上述研究相比本研究去根处理时间尚较短有关。
凋落物和根系对土壤呼吸的影响是一个复杂的生物学过程,其贡献存在明显的空间变异,此外,还随植被类型的不同而变化。陈光水等[31]报道的中国森林土壤呼吸模式中矿质土壤呼吸占土壤呼吸的比例变化范围为25.1%~69.7%,枯枝落叶层呼吸占土壤呼吸的比例变化范围为1.7%~49%,根系呼吸占土壤呼吸的比例变化范围为10%~65%。本研究中,矿质土壤呼吸速率、凋落物呼吸速率和根系呼吸速率对土壤总呼吸的贡献率分别为41.24%、43.29%和15.45%,研究结果符合中国森林土壤呼吸的一般模式。
4.3 土壤环境因子对土壤呼吸的影响
土壤温度和湿度是影响土壤呼吸的重要环境因子,大量研究认为,土壤呼吸与表层土壤温度具有良好的相关性[6],但与土壤湿度的相关性却因树种和区域的不同而差异较大,表现为正相关(促进)[16]、负相关(抑制)[32]或无显著相关性[19]。肖胜生等[16-17]研究指出,在南亚热带海岸沙地上,土壤温度变异不大,但水分亏缺严重,土壤水分是土壤呼吸的重要限制因子。本研究中,因为林分郁闭度较高,不同处理的土壤温度和湿度并没有显著的差异,不同处理土壤呼吸速率与表层土壤温度均呈显著的指数相关关系,与土壤湿度呈显著的正相关。随着土壤温度和含水量的增加,微生物活性和根系代谢增强,土壤呼吸的峰值出现在一年中温度和水分均较高的季节,土壤温度和湿度的升高对土壤呼吸的促进具有明显的交互作用,采用温度和湿度2因子进行模型拟合均比单因子模型提高拟合精度,双因子复合模型可解释土壤呼吸变异的45%~69%。
Q10值反映土壤呼吸速率对温度的敏感性,其受土壤生物和底物质量等多种因素的调控。改变碳输入对Q10值的影响也存在明显树种和区域差异,表现为升高[33]或降低[32]。Boone等[34]研究发现,在Harvad森林中进行加倍凋落物、去除凋落物和去除根系处理后,其土壤呼吸的Q10值分别为3.4、3.1和2.5,均低于对照处理的Q10值3.5。本研究中,去除根系、去除凋落物和加倍凋落物的Q10值分别为1.58、1.55和1.52,低于对照的Q10值1.77,与Boone等研究结果一致,即不同改变碳输入方式均降低了土壤呼吸的Q10值。已有研究显示,在不同森林生态系统或者气候条件下,土壤呼吸的Q10值差异很大[35],陈光水等[31]报道的中国森林土壤呼吸Q10值变化范围为1.33~5.33,本研究结果在此范围之内。
[1] CHEN Youchao, LUO Ji, LI Wei, et al. Comparison of soil respiration among three different subalpine ecosystems on eastern Tibetan Plateau, China[J]. Soil Science and Plant Nutrition, 2014, 60(2): 231.
[2] LAL R. Soil carbon sequestration impacts on global climate change and food security[J]. Science, 2004, 304: 1623.
[3] HUANG Yuhui, LI Yuelin, XIAO Yin, et al. Controls of litter quality on the carbon sink in soils through partitioning the products of decomposing litter in a forest succession series in South China[J]. Forest Ecology and Management, 2011, 261(7): 1170.
[4] ZENG Xinhua, ZHANG Wanjun, SHEN Huitao, et al. Soil respiration response in different vegetation types at Mount Taihang, China[J]. Catena, 2014, 116: 78.
[5] SCHLESINGER W H, ANDREWS J A. Soil respiration and the global carbon cycle[J]. Biogeochemistry, 2000, 48(1): 7.
[6] 汪金松, 赵秀海, 张春雨, 等. 改变C源输入对油松人工林土壤呼吸的影响[J]. 生态学报, 2012, 32(9): 2768. WANG Jinsong,ZHAO Xiuhai,ZHANG Chunyu,et al. Changes of carbon input influence soil respiration in aPinustabulaeformisplantation[J]. Acta Ecologica Sinica, 2012, 32(9): 2768.
[7] DEFOREST J L, CHEN J, MCNULTY S G. Leaf litter is an important mediator of soil respiration in an oak-dominated forest[J]. International Journal of Biometeorology, 2009, 53(2): 127.
[8] RAICH J W, SCHLESINGER W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate[J]. Tellus, 1992, 44(2): 81.
[9] SAYER E J, POWERS J S, TANNER E V J. Increased litterfall in tropical forests boosts the transfer of soil CO2to the atmosphere[J]. Plos One, 2007, 2(12): e1299.
[10] FONTAINE S, BAROT S, BARRÉ P, et al.Stability of organic carbon in deep soil layers controlled by fresh carbon supply[J]. Nature, 2007, 450(7167): 277.
[11] SAYER E J, TANNER E V J, CHEESMAN A W. Increased litterfall changes fine root distribution in a moist tropical forest[J]. Plant and Soil, 2006, 281(1): 5.
[12] 左强,何怀江,赵秀海. 改变碳源输入对蛟河红松阔叶混交林土壤呼吸的影响[J]. 应用与环境生物学报, 2015, 21(6): 1136. ZUO Qiang,HE Huaijiang,ZHAO XiuHai. Influence of carbon input changes on the soil respiration in a mixed broadleaved-Korean pine forest in western foothill of Changbai Mountain[J]. Chin J Appl Environ Biol, 2015, 21(6): 1136.
[13] HANSON P J, EDWARDS N T, GARTEN C T, et al. Separating root and soil microbial contributions to soil respiration: A review of methods and observations[J]. Biogeochemistry, 2000, 48(1): 115.
[14] 高伟,叶功富,游水生,等. 沙质海岸带木麻黄林与天然林主要种群的生态位特征比较[J]. 生态与农村环境学报, 2011, 27(2): 35. GAO Wei,YE Gongfu,YOU Shuisheng,et al. Comparison betweenCasuarinaequisetifoliaforest and natural forest on sandy coast in niche characteristics[J]. Journal of Ecology and Rural Environment,2011, 27(2): 35.
[15] 游水生,叶功富,徐俊森,等. 福建东山岛海岸带潺槁树种群生命表分析[J]. 广西植物, 2009, 29(1): 96. YOU Shuisheng,YE Gongfu,XU Junsen,et al. Analysis of life table ofLitseaglutinosapopulation on the coast in Dongshan Island[J].Guihaia,2009, 29(1): 96.
[16] XIAO S S, YE G F, ZHANG L H, et al. Soil heterotrophic respiration inCasuarinaequisetifoliaplantation at different stand ages[J]. Journal of Forestry Research, 2009, 20(4): 301.
[17] 肖胜生,叶功富,董云社,等. 木麻黄沿海防护林土壤呼吸动态及其关键影响因子[J]. 中国环境科学, 2009, 29(5): 531. XIAO Shengsheng,YE Gongfu,DONG Yunshe,et al. Soil respiration dynamics and major controlling factors in a coastal protective forest[J]. China Environmental Science, 2009, 29(5): 531.
[18] 高伟,叶功富,卢昌义,等. 福建东山岛海岸带风水林数量分类与排序[J]. 中南林业科技大学学报, 2013, 33(8): 117. GAO Wei,YE Gongfu,LU Changyi,et al. Quantitative classification and sequence of Feng-shui forest in coastal zone of Dongshan island in Fujian[J]. Journal of Central South University of Forestry and Technology, 2013, 33(8): 117.
[19] 肖胜生,熊永,段剑,等. 基于组分区分的南方红壤丘陵土壤呼吸对植被类型转换的响应[J]. 农业工程学报, 2015, 31(14): 123. XIAO Shengsheng,XIONG Yong,DUAN Jian,et al. Responses of soil respiration to vegetation type conversion in south hilly red soil based on main components[J]. Transactions of the Chinese Society of Agricultural Engineering, 2015, 31(14): 123.
[20] SULZMAN E W, BRANT J B, BOWDEN R D, et al. Contribution of aboveground litter, belowground litter, and rhizosphere respiration to total soil CO2efflux in an old growth coniferous forest[J]. Biogeochemistry, 2005, 73(1): 231.
[21] LEFF J W, WIEDER W R, TAYLOR P G, et al. Experimental litterfall manipulation drives large and rapid changes in soil carbon cycling in a wet tropical forest[J]. Global Change Biology, 2012, 18(9): 2969.
[22] 高强,马明睿,韩华,等. 去除和添加凋落物对木荷林土壤呼吸的短期影响[J]. 生态学杂志, 2015, 34(5): 1189. GAO Qiang,MA Mingrui,HAN Hua,et al.Short-term effects of aboveground litter exclusion and addition on soil respiration in aSchimasuperbaforest in Zhejiang Province,Eastern China[J]. Chinese Journal of Ecology, 2015, 34(5): 1189.
[23] WARDLE D A. A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil[J]. Biological Reviews, 2008, 67(3): 321.
[24] ZAK D R, TILMAN D, PARMENTER R P, et al.Plant production and soil microorganisms in late-successional ecosystems: a continental-scale study[J]. Ecology, 1994, 75(8): 2333.
[26] HOOKER T D, STARK J M. Soil C and N cycling in three semiarid vegetation types: response to an in situ pulse of plant detritus[J]. Soil Biology and Biochemistry, 2008, 40(10): 2678.
[27] FENG Wenting, ZOU Xiaoming, SCHAEFER D. Above- and belowground carbon inputs affect seasonal variations of soil microbial biomass in a subtropical monsoon forest of southwest China[J]. Soil Biology and Biochemistry, 2009, 41(5): 978.
[28] HART S C, SOLLINS P. Soil carbon and nitrogen pools and processes in an old-growth conifer forest 13 years after trenching[J]. Canadian Journal of Forest Research, 1998, 28(8): 1261.
[29] LI Yiqing, XU Ming, SUN O J, et al. Effects of root and litter exclusion on soil CO2efflux and microbial biomass in wet tropical forests[J]. Soil Biology and Biochemistry, 2004, 36(12): 2111.
[30] BRANT J B, MYROLD D D, SULZMAN E W. Root controls on soil microbial community structure in forest soils[J]. Oecologia, 2006, 148(4): 650.
[31] 陈光水,杨玉盛,吕萍萍,等. 中国森林土壤呼吸模式[J]. 生态学报, 2008, 28(4): 1748. CHEN Guangshui,YANG Yusheng,LYU Pingping,et al.Reginal patterns of soil respiration in China’s forest[J]. Acta Ecologica Sinica, 2008, 28(4): 1748.
[32] 余再鹏,万晓华,胡振宏,等. 亚热带杉木和米老排人工林土壤呼吸对凋落物去除和交换的响应[J]. 生态学报, 2014, 34(10): 2529. YU Zaipeng,WAN Xiaohua,HU Zhenhong,et al. Contrasting responses of soil respiration to litter manipulation in subtropicalMytilarialaosensisandCunninghamialanceolataplantations[J]. Acta Ecologica Sinica, 2014, 34(10): 2529.
[33] 王光军,田大伦,闫文德,等. 改变凋落物输入对杉木人工林土壤呼吸的短期影响[J]. 植物生态学报, 2009, 33(4): 739. WANG Guangjun,TIAN Dalun,YAN Wende, et al. Effects of aboveground litter exclusion and addition on soil respiration in aCunninghamialanceolataplantation in China[J].Chinese Journal of Plant Ecology, 2009, 33(4): 739.
[34] BOONE R D, NADELHOFFER K J, CANARY J D, et al. Root exert a strong influence on the temperature sensitivity of soil respiration. Nature[J]. Nature, 1998, 396(6711): 570.
[35] PENG Shushi, PIAO Shilong, WANG Tao, et al.Temperature sensitivity of soil respiration in different ecosystems in China[J]. Soil Biology and Biochemistry, 2009, 41(5): 1008.
Changes of carbon input influence soil respiration in a typical natural secondary forest on southern subtropical sandy coast
GAO Wei1,2,4, YE Gongfu4, HUANG Zhiqun1,2,3, CHEN Zhiyong5,
YU Zaipeng1,2,3, YUE Xinjian5, CHEN Mengyao5
(1.Cultivation Base of State Key Laboratory of Humid Subtropical Mountain Ecology, 350007, Fuzhou, China; 2.Institute of Geography, Fujian Normal University, 350007, Fuzhou, China; 3.Ministry of Education Key Laboratory of Humid Subtropical Eco Geographical Process, 350007, Fuzhou, China; 4.Fujian Academy of Forestry Science, 350012, Fuzhou, China; 5.College of Forestry, Fujian Agriculture and Forestry University, 350002, Fuzhou, China)
[Background] Globally, the pool of carbon in soils is greater than the pool contained in vegetation or the atmosphere, and changes in soil carbon content can therefore greatly affect the global carbon budget. Soil respiration is an essential process of carbon cycling in terrestrial ecosystems, thus the knowledge about influence of environmental factors and carbon input on soil respiration is critical for ecosystem management. [Methods] Four 20 m×20 m permanent fixed plots were established in a natural secondary forest in December 2014 in southeastern coast of Fujian Province, China, and four carbon input manipulations of root exclusion, litter exclusion, double litter and control(normal carbon inputs)were carried out in each plot. Monthly soil respiration (Rs) under different carbon input manipulations were measured by Li-8100 from March 2015 to February 2016. In addition, the dissolved organic carbon and microbial biomass carbon were determined afterRsmeasurement. One-way analysis of variance (ANOVA) with Duncan’s multiple comparison method was used to test the differences ofRs, soil temperature and soil moisture between different carbon input manipulations, besides the linear and nonlinear regression models were used to describe the relationship between seasonalRs, soil temperature and soil moisture. [Results] The results showed that no significant difference was found for soil temperature and soil moisture at 10 cm depth between different treatments (P>0.05). The seasonal pattern ofRswas obviously, which was higher in summer and lower in winter during the study period, and the highest fluxes observed in May 2015 or June 2015 and the lowest fluxes occurred in November 2015 or December 2015. The range of annual meanRsfor different treatments was double litter > control >root exclusion > litter exclusion, and allQ10values in different carbon input manipulations were lower than that in control. The contribution of mineral soil, litter layers and roots to total soil respiration was 41.24%, 43.29% and 15.45%, respectively.Rswas significantly and exponentially correlated with soil temperature and linearly and positively correlated with soil moisture at 10 cm depth (P<0.05). The bi-factor models integrated W10with T10explained 45%~69% of the variations forRs. The soil dissolved organic carbon and microbial biomass carbon were also influenced by carbon input changes, and the meanRssignificantly correlated with soil dissolved organic carbon and microbial biomass carbon concentration.[Conclusions] Our findings suggested that carbon input manipulations induced changes in soil labile carbon, and subsequently caused significant effects on soil respiration.
southern subtropical; sandy coast; natural secondary forest; soil respiration; carbon input;Q10values
2016-07-16
2017-03-28
项目名称: 国家自然科学基金“亚热带人工林树种对土壤氮吸收和转化的影响”(41371269);“亚热带森林土壤氮梯度、植物养分与土壤有机质来源变化耦合关系”(31570604);福建省属公益类科研院所基本科研专项“滨海沙地造林树种转换对土壤碳循环的影响”(2014R1011-7);福建省森林培育与林产品加工利用重点实验室、福建省木麻黄工程技术研究中心资助
高伟(1985—),男,博士研究生。主要研究方向:沿海防护林生态构建及海岸带生态系统恢复。E-mail:gao01271@163.com
†通信作者简介: 黄志群(1973—),男,博士,教授。主要研究方向:亚热带人工林经营及森林生态系统碳、氮循环。E-mail:zhiqunhuang@hotmail.com
S154.1;X833
A
2096-2673(2017)02-0009-09
10.16843/j.sswc.2017.02.002
