皱皮油丹 (樟科) 在云南东南部的发现及其生物地理学意义
2017-05-30莫月清李朗张金国李锡文李捷
莫月清 李朗 张金国 李锡文 李捷


摘要: 该文报道了在云南省马关县古林箐省级自然保护区发现的一个云南省新记录种——皱皮油丹 (Alseodaphne rugosa Merr. et Chun),并对其形态、生境及该新记录种的生物地理学意义进行了描述与讨论。此新记录种的发现对樟科油丹属植物的系统分类学、生物地理学和区域生物多样性等方面的研究具有重要意义,同时也为探讨海南岛的起源问题提供了具有价值的研究材料。
关键词: 皱皮油丹, 樟科, 新记录种, 生物地理学, 海南岛
中图分类号: Q948.15文献标识码: A文章编号: 10003142(2017)07091209
樟科 (Lauraceae) 隶属于被子植物门 (Angiospermae) 木兰亚纲 (Magnoliid) 樟目(Laurales) (Reveal & Chase, 2011),是一个较大的木本植物类群[除草本寄生无根藤属(Cassytha Linn.)外],全世界约50属,2 500~3 000种,广泛分布于世界热带至亚热带地区(Rohwer, 1993; Chanderbali et al, 2001; 李捷和李锡文,2004)。
油丹属 (Alseodaphne Nees) 隶属于樟科,全球50余种,90%的物种集中分布于亚洲的热带地区,具体分布范围自斯里兰卡,经印度、中南半岛及我国南部,至马来西亚、印度尼西亚及菲律宾。我国约有10种,产云南南部及广东、海南 (李锡文等,1982; Li et al, 2008)。
在我国分布的10种油丹属植物分别为毛叶油丹 [Alseodaphne andersonii (King ex Hook. f.) Kosterm.]、细梗油丹 (A. gracilis Kosterm.)、油丹 (A. hainanensis Merr.)、河口油丹 (A. hokouensis H. W. Li)、黄连山油丹 (A. huanglianshanensis H. W. Li & Y. M. Shui)、麻栗坡油丹 [A. marlipoensis (H. W. Li) H. W. Li]、长柄油丹 [A. petiolaris (Meissn.) Hook. f.]、皱皮油丹 (A. rugosa Merr. et Chun)、西畴油丹 (A. sichourensis H. W. Li) 和云南油丹 (A. yunnanensis Kosterm.),它们主要分布在广东、云南和海南 (李锡文等,1982; Li et al, 2008)。皱皮油丹是我国的特有油丹属物种之一,之前报道仅分布在海南,为海南特有种 (陈焕镛,1964)。关于海南岛的特有种有较多文献报道,大多学者希望借助海南岛的特有种来揭示其植物区系的起源和年龄(邢福武等,1995;王发国等,2006;秦新生等,2011),而且大多学者都认为海南岛的植物区系起源于华夏植物区系(缪汝槐,1986;王伯荪,1982;张超常和刘兰芳,1983)。
海南岛是我国仅次于台湾岛的第二大岛屿,关于海南岛的研究文献不在少数。在以往的研究文献中,不少学者研究发现海南岛与大陆有很多相似性(地质特征相似,植物区系关系密切),并推测海南岛可能是从大陆分离出去的,而关于海南岛从大陆分离的位置也存在不少争议,有的学者认为海南岛是从广东省分离出去的(林晓东和宗永强,1987;赵焕庭等,2007),而有的学者则认为海南岛是从北部湾分离出去的 (Zhu, 2016; 梁光河,2013)。
2016年5月至8月期间,我们在云南省马关县古林箐省级自然保护区采集到一种油丹属植物标本若干份,经鉴定为皱皮油丹,是该物种在云南省分布的新记录。此新记录种的发现对樟科油丹属植物的系统分类学、生物地理学和区域生物多样性等方面的研究具有重要的意义,同时也为探讨海南岛的起源问题提供了具有价值的研究材料。
1研究方法
1.1 形态特征比较鉴定
在野外采集生长良好,具有代表性的枝条,充分干燥制作成标本,对该物种初步鉴定为皱皮油丹。但由于皱皮油丹为海南特有種,尚未在云南发现,所以还通过查阅《中国植物志》第31卷樟科 (李锡文等,1982)、Flora of China(第7卷樟科) (Li et al, 2008)、《海南植物志》第1卷樟科 (陈焕镛等,1964)以及皱皮油丹发表时的原始文献 (Merrill & Chun, 1935)中皱皮油丹的形态描述,并和皱皮油丹模式标本以及以往在海南采集到的皱皮油丹标本进行形态比较研究,以进一步鉴定该物种。
1.2 分子系统学手段鉴定
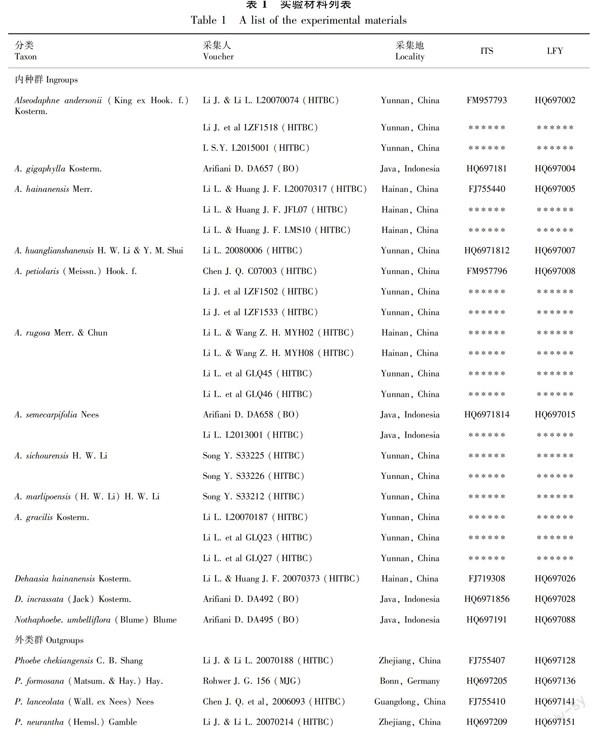
1.2.1 实验材料为进一步确定该种的分类学身份,本研究增加了分子系统学的研究方法。根据Li et al (2011)的研究方法,我们选取ITS (Internal Transcribed Spacer) 和LEAFY intronⅡ序列作为分子标记。共选取油丹属10种共23个个体(主要包括海南采集到的皱皮油丹MYH02、MYH08两个个体,以及云南采集到的皱皮油丹GLQ45、GLQ46两个个体,其余油丹属种类详见表1);此外,还包括莲桂属 (Dehaasia Bl.) 2种,赛楠属 (Nothaphoebe Bl.) 1种,共计13个种类作为内类群,选取楠属 (Phoebe Nees) 4种作为外类群。详见表1。
1.2.2 实验方案本研究使用的实验方法 (包括DNA提取,PCR扩增,测序等) 沿袭Li et al (2011)的研究方法来进行,形成ITS和LEAFY intronⅡ序列联合矩阵,并用贝叶斯分析 (Bayesian Inference, BI) 和最大简约法 (maximum parsimony, MP) 对联合矩阵进行系统发育分析。
贝叶斯分析使用 MrBayes 3.1.2(Huelsenbeck & Ronquist, 2001; Ronquist & Huelsenbeck, 2003) 软
件进行运算,对4个平行进化链进行200万代运算。马尔可夫链 (Makov chains) 每100代取样1次,得到的系统进化树去掉前25%作为 “burn-in”,然后对剩下的系统进化树进行一致性分析,得到系统进化树,支持率用后验率 (posterior probability value) 表示。
最大简约法分析使用PAUP 4.0b10 (Swofford, 2003)软件进行运算,分析采用启发式搜索 (Heuristic search),具体参数为random addition sequence with 1000 replicates, TBR branch-swapping, STEEPEST DESCENT, MULTREES in effect, all characters were unordered, equally weighted。得到的系统进化树使用靴带法进行检测 (100 Bootstrap replicates),支持率用靴带支持率 (Bootstrap value) 表示。
2研究结果
2.1 皱皮油丹形态学比较结果
通过对皱皮油丹模式标本的比对,以及对海南和云南两地该种的形态比较,我们发现云南所采集到的该油丹属物种与皱皮油丹,除在果实大小存在差异外,其它特征较为吻合。
常绿乔木,高达13 m,胸径达40 cm。老枝粗壮,灰白色,具光泽,有多数纵向开裂的突起的褐色椭圆形皮孔和近圆形大叶痕。叶互生,集生于枝梢,长圆状倒卵形或长圆状倒披针形,长12~32 cm,宽3.5~9 cm,先端短渐尖,基部楔形,革质,上面干时浅棕色,光亮,下面绿白色,中脉浅棕色,上面凹陷,下面明显凸起,浅棕色,细脉显著,网状;叶柄粗壮,长1~2.5 cm,腹面具槽,背面圆形。圆锥花序腋生于去年生枝条近顶端,长15~20 cm,多花,无毛,多分枝;总梗长4.5~10 cm;花梗纤细,长5~8 mm,无毛。花被片6,近等大,外面无毛,内面密被白色柔毛,外轮花被片宽卵形,长2 mm,宽1.5 mm,先端锐尖,内轮花被片宽卵形,长2.5 mm,宽2.2 mm,先端锐尖,果时均脱落。能育雄蕊9,第一轮雄蕊长约2 mm,花丝长约1.5 mm,被疏柔毛,花药椭圆形,先端截平,4室,上2室较小,下两室较大,药室均内向,第二轮雄蕊长约1.8 mm,花丝稍长于花药,被疏柔毛,花药椭圆形先端截平,4室,上2室较小,下两室较大,药室均内向,第三轮雄蕊长2.2 mm,花丝稍长于花药,被疏柔毛,近基部有成对圆状心形具短柄腺体,花药长圆形,4室,上2室较小,下两室较大,均外向。退化雄蕊明显,长1.8 mm,三角状箭头形,具柄。子房椭球形,长1.2 mm,无毛,花柱长0.8 mm,柱头盘状,不明显。果序近顶生,粗壮,长10~18 cm,无毛。果扁球形,长4 cm,直径4.5 cm;果梗粗壮,长1~2.5 cm,顶端直径0.5~1 cm,鲜时肉质,多疣。花期5—6月,果期7—10月。
以上形态描述依据云南新记录的标本做出 (图1, 图2: AB)。
凭证标本: 云南省马关县,古林箐省级自然保护区:李朗 (Li L),刘志芳 (Liu LZ),莫月清 (Mo YQ),张金国 (Zhang JG),GLQ26 (HITBC), GLQ33 (HITBC), GLQ45 (HITBC), GLQ46 (HITBC), 2016年5月16日。云南省马关县,古林箐省级自然保护区:李朗 (Li L),蔡超男(Cai CN),莫月清 (Mo YQ) ,张金国 (Zhang JG),20160032 (HITBC), 20160033 (HITBC), 20160036 (HITBC), 2016007 (HITBC), 20160038 (HITBC), 20160040 (HITBC),2016年8月28日。
海南省鸚哥岭国家级自然保护区:李朗 (Li L),黄建峰 (Huang JF),2013035 (HITBC), 2013026 (HITBC), 2013年4月18日。海南省鹦哥岭国家级自然保护区:李朗 (Li L),2007369 (HITBC), 2007370 (HITBC), 2007371 (HITBC), 2007年11月27日。海南省保亭县,兴隆:侯宽昭 (Hou KZ),73452 (IBK),1935年8月14日。海南省定安县,五指山:黄志 (Huang Z),35424 (IBK),1933年12月15日。海南省五指山:陈念驹 (Chen N J),44254 (IBK),1932年11月11日。
地理分布和生境:皱皮油丹分布于我国海南省鹦哥岭、吊罗山、黎母山、尖峰岭、五指山和霸王岭,和云南东南部,生长在海拔700~1 300 m的山谷密林中。与皱皮油丹亲缘关系密切 (Li et al, 2011) 的油丹同样分布在海南的鹦哥岭、吊罗山、黎母山、尖峰岭、五指山和霸王岭,另一分布地为越南北部老街省,与云南省马关县的地理距离极近。
应该指出的是云南产的皱皮油丹与海南产的存在果实大小差异,云南皱皮油丹果长3.5~4.5 cm,直径4~5 cm,海南皱皮油丹果长2~3 cm,直径2.5~3.5 cm,果实形态几无差异(图2:A,DF),仅依据较大果实这一差别特征另立一新种,显然是不合适的。鉴于该物种在云南与海南的分布存在地理距离和生境差异,且目前掌握的研究材料尚不是十分充分,特别是目前没有观察到海南皱皮油丹的花形态特征,故暂不考虑作种下分类处理。
2.2 系统发育分析结果
基于ITS+LEAFY intronⅡ联合矩阵,使用贝叶斯和最大简约法分析得到的系统发育树一致,因此在本文中只展示贝叶斯分析结果。在云南分布的皱皮油丹的两个个体 (GLQ45、GLQ46) 与在海南分布的皱皮油丹的两个个体 (MYH02、MYH08) 形成一单系分支 (93%的靴带支持率和100%的后验率,见图3中突出显示个体);此两地的个体又分别形成单系分支 (均有100%的靴带支持率和后验率),说明云南皱皮油丹鉴定可靠;其中皱皮油丹又与主要分布于海南的另一油丹属物种——油丹的系统关系非常密切 ,二者在系统发育树上形成一单系分支(100%的靴带支持率和后验率) (图3)。
综上所述,结合形态学比较和分子系统学的研究结果,于云南采集到的油丹属物种确认为皱皮油丹,此为皱皮油丹在云南的新记录。3新记录种发现的生物地理学意义
皱皮油丹是海南的特有物种,如今在云南新记录的发现,对于该物种的生物地理学以及区域生物多样性研究均有着非常重大的意义。皱皮油丹在云南和海南两地呈间断分布格局的现象,证实了海南岛与大陆之间存在联系。关于海南岛的起源问题已经有相当多文献的报道,其中影响较大的是梁光河 (2013)在前人研究的基础上提出的海南岛从大陆北部湾分离出去的8项证据:①东南亚地块逃逸构造与新特提斯演化在地质上得到了广泛认可;②地形地貌和山脊复原后能够得到极好的吻合;③地质层位、断裂带、岩浆岩复原后能够得到极好的吻合;④石油地震勘探和深海油气钻探发现了海南岛小齿轮的边界;⑤琼东南中央凹陷带油气钻探结果证明其中沉积物来源于红河冲积物;⑥GPS观测证实海南岛目前仍然在向东南漂移并伴随左旋;⑦海南岛大型非本土动物存在的历史说明其曾经与大陆相连;⑧人口分布特征的一致性。这些证据均说明海南岛起源于大陆。
此外,有关化石方面的报道也支持了此观点。刘裕生 (1990) 报道了发现于广西百色盆地更新统长蛇岭组的油丹近似种 (Alseodaphne cf. hainanensis Merr.) 的叶片化石;韦利杰等 (2005) 曾于云南腾冲上新统芒棒组采到油丹 (A. hainanensis Merr.) 的叶片化石;Hu et al(2007) 对浙江嵊州上新世地层中发现的油丹近似种 (Alseodaphne cf. hainanensis Merr.) 的叶表皮特征以及古气候进行了详细研究。李景照等 (2009) 在海南岛长昌盆地始新世长昌组中发现了樟科植物一新种长昌油丹 (Alseodaphne changchangensis) 的完整叶片化石标本,这是迄今发现的时代最早、纬度最低的油丹属化石,并发现其与现生油丹 (A. hainanensis Merr.) 最为相似。以上这些化石证据说明油丹曾在大陆和海南广泛分布,随着地质变迁与气候变化,油丹分布范围逐渐缩小,如今主要分布在越南北部及中国海南 (李锡文等,1982),呈现间断分布格局。而皱皮油丹与油丹的系统发育关系非常密切,在系统发育树上呈姊妹关系,如今皱皮油丹在云南新记录的发现,使其和油丹呈现类似的间断分布格局(云南东南部与海南、越南北部与中国海南;中国云南东南部与越南北部毗邻),暗示了海南岛与大陆之间的联系。
一个岛屿的特有植物能够在一定程度上揭示它的起源和年龄以及演化规律,很多学者研究发现海南岛的特有植物比例较低,推测其起源于大陸。Zhu (2016) 更是详细研究了海南岛种子植物共196科,1 283属和3 894种的分布式样,并将越南的植物区系以及中国的广西和广东等地的植物区系与中国海南的植物区系相比较,结果发现海南岛的植物区系在科属水平上以热带分布为主,亚洲热带分布占23.85%,而东亚分布仅占3.74%,且只有7属和约10%的物种是海南岛特有的,如此低比例的特有属种暗示了其起源于大陆。此外,Zhu (2016)的研究还发现中国海南岛的种子植物种类与越南地区以及中国的广西和云南地区在科属水平上有极大的相似性,故推测中国海南岛曾与越南北部和中国的广西接壤。
本研究发现海南的特有种——皱皮油丹在云南也有分布,恰恰支持了关于海南岛起源于大陆,并从大陆分离出去的观点。与皱皮油丹一同分布于中国海南岛的油丹,也已经被发现在越南北部有所分布 (李锡文等,1982)。这也支持和印证了中国海南岛曾与中国的广西以及越南北部接壤的观点。
致谢野外考察得到了云南省林业厅、古林箐省级自然保护区管理局的大力支持;采样工作得到了中国科学院西双版纳热带植物园植物系统发育与保护生物学实验室刘志芳同学、蔡超男同学的协助;该物种线条图由中国科学院昆明植物研究所王凌老师完成。在此一并表示感谢。
参考文献:
CHANDERBALI AS, VAN DER WERFF H, RENNER SS, 2001. Phylogeny and historical biogeography of Lauraceae: Evidence from the chloroplast and nuclear genomes [J]. Ann Mo Bot Gard, 88(1): 104-134. doi:10.2307/2666133.
CHEN HY, 1964. Alseodaphne N [M]//Flora of Hainan. Tomus 1. Beijing: Science Press: 265-266. [陈焕镛, 1964. 油丹属 [M]//海南植物志:第1卷. 北京: 科学出版社: 265-266.]
CHEN J Q, LI L, LI J, et al, 2009. Bayesian inference of nrDNA ITS sequences from Machilus (Lauraceae) and its systematic significance [J]. Acta Bot Yunnan, 31(2): 117-126. [陈俊秋, 李朗, 李捷, 等, 2009. 樟科润楠属植物ITS序列贝叶斯分析及其系统学意义 [J]. 云南植物研究, 31(2): 117-126.]
HUELSENBECK JP, RONQUIST F, 2001. MrBayes: Bayesian inference of phylogenetic trees [J]. Bioinformatics, 17(8): 754-755.
HU YQ, FERGUSON DK, LI CS, et al, 2007. Alseodaphne (Lauraceae) from the Pliocene of China and its paleoclimatic significance [J]. Rev Palaeobot Palynol, 146(1-4), 277-285. doi:10.1016/j.revpalbo.2007.06.002.
LIAO RH, 1986. A gymnospermous floristic analysis of Hainan Island [J]. Acta Sci Nat Univ Sunyatseni (Nat Sci Ed), 25(4): 59-65. [缪汝槐, 1986. 海南岛裸子植物区系 [J]. 中山大学学报(自然科学版), 25(4): 59-65.]
LIANG GH, 2013. Eight evidences about Hainan Island separated from Chinas Beibuwan Gulf with drifting and rotation [J]. Acta Geol Sin, 87(Z): 73-76. [梁光河, 2013. 海南岛从中国北部湾分离旋转漂移出去的8大证据 [J]. 地质学报, 87(Z): 73-76.]
LI HW, BAI PY, LI SG, et al, 1982. Alseodaphne N [M]//Flora Reipublicae Popularis Sinicae. Tomus 31. Beijing: Science Press: 68-79. [李锡文,白佩瑜,李树刚,等, 1982. 油丹属 [M]//中国植物志:第31卷. 北京: 科学出版社: 68-79.]
LI HW, LI J, HUANG PH, et al, 2008. Lauraceae [M]//WU ZY, RAVEN PH, HONG DY. Flora of China. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press, 7: 102-254.
LI J, LI HW, 2004. Advances in Lauraceae systematic research on the world scale [J]. Acta Bot Yunnan, 26(1): 1-11. [李捷, 李锡文, 2004. 世界樟科植物系统学研究进展 [J]. 云南植物研究, 26(1): 1-11.]
LI JZ, QIU J, LIAO WB, et al, 2009. Eocene fossil Alseodaphne from Hainan Island of China and its paleoclimatic implications [J]. Sci Chin (Ser D: Earch Sci), 39(12): 1753-1758. [李景照, 邱玨, 廖文波,等, 2009,. 海南岛始新世Alseodaphne化石及其古气候指示 [J]. 中国科学 (D辑:地球科学), 39(12): 1753-1758.]
LI L, LI J, ROHWER JG, et al, 2011. Molecular phylogenetic analysis of the Persea group (Lauraceae) and its biogeographic implications on the evolution of tropical and subtropical AmphiPacific disjunctions [J]. Am J Bot, 98(9): 1520-1536.
LIU XD, ZONG YQ, 1987. More on the origin of the Qiongzhou Strait [J]. Trop Geogr, 7(4): 338-345. [林晓东, 宗永强, 1987. 再论琼州海峡成因 [J]. 热带地理, 7(4): 338-345.]
LIU YS, 1990. Cuticular studies on two Pleistocene species of Lauraceae in Baise Basin, Guangxi [J]. Acta Bot Sin, 32(10): 805-808+829-830. [刘裕生, 1990. 广西百色盆地更新世樟科两种植物角质层研究 [J]. 植物学报, 32(10): 805-808+829-830.]
MERRILL, CHUN, 1935. Additions to our knowledge of the Hainan flora Ⅱ [J]. Sunyatsenia, 2(3/4): 244-245.
QIN XS, ZHONG YF, SONG XQ, et al, 2011. The wild endemic resources of ornamental plants and their utilization in Hainan Island [J]. Chin Landsc Architect, 27(7): 72-78. [秦新生, 钟云芳, 宋希强, 等, 2011. 海南特有野生花卉资源及其利用 [J]. 中国园林, 27(7): 72-78.]
REVEAL JL, CHASE MW, 2011. APG III: Bibliographical information and synonymy of Magnoliidae [J]. Phytotaxa, 19(1): 71-134.
ROHWER JG, 1993. Lauraceae [M]//KUBIZKI K, ROHWER JG, BITTRICH V. The families and genera of vascular plants; Berlin: Springer-Verlag, 2: 366-391.
ROHWER JG, LI J, RUDOLPH B, et al, 2009. Is Persea (Lauraceae) monophyletic? Evidence from nuclear ribosomal ITS sequences [J]. Taxon, 58(4): 1153-1167.
RONQUIST F, HUELSENBECK JP, 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models [J]. Bioinformatics, 19(12): 1572-1574. doi:10.1093/bioinformatics/btg180
SWOFFORD DL, 2003. PAUP*: Phylogenetic analysis using parsimony (*and other methods), version 4.0b10 [J]. Sunderland: Sinauer Associates.
WEI LJ, SUN BN, XIE SP, et al, 2005. Epidermal microstructural studies on plant Alseodaphne Hainanensis Merr. from the pliocene of Tengchong in Yunnan Province [J]. Acta Micropalaeontol Sin, 22(4): 392-399. [韦利杰, 孙柏年, 解三平, 等, 2005. 云南腾冲上新统植物油丹Alseodaphne hainanensis Merr.表皮微细构造研究 [J]. 微体古生物学报, 22(4): 392-399.]
WANG BS, 1982. The pteridophyte flora of Hainan [J]. Acta Sci Nat Nat Univ Sunyatseni (Nat Sci Ed), 21(1): 94-100. [王伯荪,1982. 海南岛蕨类植物区系 [J]. 中山大学学报(自然科学版), 21(1): 94-100.]
WANG FG, QIN XS, CHEN HF, et al, 2006. Endemic plants in limestone region on Hainan Island [J]. J Trop & Subtrop Bot, 14(1): 45-54. [王发国, 秦新生, 陈红锋,等, 2006. 海南岛石灰岩特有植物的初步研究 [J]. 热带亚热带植物学报, 14(1): 45-54.]
XING FW, WU DL, LI ZX, et al, 1995. Eedemic plants of Hainan Island [J]. J Trop & Subtrop Bot, 3(1): 1-12. [邢福武,吴德邻,李泽贤,等, 1995. 海南岛特有植物的研究 [J]. 热带亚热带植物学报, 3(1): 1-12.]
ZHANG CC, LIU LF, 1983. The angiospermous flora of Hainan [J]. Acta Sci Nat Univ Sunyatseni (Nat Sci Ed), 22(3): 69-76. [张超常, 刘兰芳, 1983. 海南岛被子植物区系 [J]. 中山大学学报(自然科学版), 22(3): 69-76.]
ZHAO HT, WANG LR, YUAN JY, 2007. Origin and time of Qiongzhou Strait [J]. Marin Geol Quat Geol, 27(2): 33-40. [趙焕庭, 王丽荣, 袁家义, 2007. 琼州海峡成因与时代 [J]. 海洋地质与第四纪地质, 27(2): 33-40.]
ZHU H, 2016. Biogeographical evidences help revealing the origin of Hainan Island [J]. PLoS ONE, 11(4), e0151941.
