山西太岳山南部针阔混交林群落特征及空间分布格局
2017-05-30王燕毕润成许强
王燕 毕润成 许强


摘要: 为探究针阔混交林群落的空间分布特性、种间关系及演替规律,该研究通过点格局分析方法中的RipleyL函数对山西太岳山南部1 hm2针阔混交林中主要乔木种群的空间分布格局及空间关联性进行了研究。结果表明:样地内共调查木本植物41科71属76种,以蔷薇科、毛茛科、唇形科、豆科、菊科、百合科为主,科的分布区类型主要以世界广布为主,属的分区类型以北温带为主;辽东栎(Quercus wutaishanica)、槲树(Quercus dentata)、油松(Pinus tabuliformis)和山杨(Populus davidiana)是该群落的主要树种,样地内的林木总径级结构与主要乔木种群的径级结构相似,呈近似正态分布。辽东栎和槲树的空间分布相似,随尺度增大种群的聚集性减弱并逐渐表现出随机分布的格局特征;油松和山杨在一定尺度范围内呈现聚集分布;主要树种均在一定尺度上表现为两两正相关,且槲树与油松、山杨在整个尺度内都表现为显著正相关。山西太岳山南部针阔混交林中各个乔木种群处于相对稳定的状态。
关键词: 针阔混交林, 群落结构, 空间分布格局, 空间关联性
中图分类号: Q948文献标识码: A文章编号: 10003142(2017)07090111
群落的组成与结构是植物群落生态学研究的基础,它不仅能反映群落中物种之间的关系,也能反映环境对物种生存的影响(李博,2000)。植物种群空间分布格局是种群个体在水平空间的配置状况或分布状态,它取决于自身和群落环境(Zhang & Meng,2004)。因此研究群落组成与空间格局有助于认识种群特征、种群间以及种群与环境之间相互作用的基本规律(John et al,2007;He & Duncan,2000),从而推断时空格局形成的潜在机制,一直是生态学研究的热点(GreigSmith,1983;Wiegand & Moloney,2004)。植物种群的空间分布格局为聚集分布(clumped distribution)、均匀分布(regular distribution)和随机分布(randonm distribution)。Ripley(1977)提出了点格局分析(point pattern analysis)的理论,该方法之所以具有较强的检验能力,是因为充分利用了点和点之间的距离。它不仅可以分析任意尺度下的种群格局和种间关系,同时还可以给出空间格局最大聚集强度下所对应的尺度,为种群空间格局研究提供方便(Druckenbrod et al,2005)。
太岳山南部自然植被多以辽东栎纯林和油松林纯林为主,是落叶阔叶林地带性植被类型,还有一些以山杨林、白皮松林和槭、椴、榆等组成的杂木林(闫明等,2003)。以往对太岳山的研究多以油松林、辽东栎林为对象,主要集中在种群径级结构(张婕等,2014)、群落学特征及空间格局(伊力塔等,2008;张笑菁等,2010;霍萌萌等,2014),光合特性(蔺琛等,2002),以及生态响应(郭东罡等,2011;金冠一等,2013;肖迪等,2016)等,这些都集中于对纯林生态学特性的研究,然而对针阔混交林的研究却少见报道。本研究以山西太岳山南部针阔混交林为例,通过野外调查,分析其群落特征,并采用点格局分析方法中的RipleyL函数对主要乔木种群的空间分布格局及空间关联性进行分析,旨在从空间格局的角度认识针阔混交林的空间分布特性、种间关系及演替的生态学过程和规律,以期为进一步促进地域森林资源的可持续利用及其保育措施提供科学依据。
1研究区自然概况
太岳林区是山西省八大林区之一,属太岳山主体部分。该区地处111°45′~112°33′ E,36°18′~37°05′ N,海拔为598~2 566.6 m。本区气候属暖温带大陆性季风气候,年平均气温为9.3~12.3 ℃,无霜期为160~180 d,年降水量为500~700 mm(曹静等,2015)。研究区地带性土壤为褐土,由于海拔差异大和土壤垂直分布明显,从低到高土壤垂直带谱为石灰性褐土带、褐土性土带、淋溶褐土带、棕色森林土、山地草甸土。植被垂直带沿海拔上升依次为灌草丛及农田带(800~1 000 m)、低中山针叶林带(1 000~1 300 m)、针叶阔叶混交林带(1 300~2 000 m)、落叶阔叶林带(1 400~2 150 m)和山地矮林和草甸带(2 000~2 345 m)(王祎玲等,2012)。调查区植被资源的乔木主要由辽东栎(Quercus wutaishanica)、槲树(Q. dentata)、油松(Pinus tabuliformis)、山杨(Populus davidiana)、野核桃(Juglans cathayensis)、千金榆(Carpinus cordata)等組成的杂木林。林下植被以土庄绣线菊(Spiraea pubescen)、水栒子(Cotoneaster multiflorus)、毛榛(Corylus mandshurica)、连翘(Forsythia suspensa)、忍冬(Lonicera japonica)、披针叶苔草(Carex lanceolata)、野青茅(Deyeuxia arundinacea)、茜草(Rubia cordifolia)为主。
2研究方法
2.1 样地设置
2015年8月,在大南坪林场附近选择人文干扰较轻的针阔混交林设置一块100 m × 100 m的典型样地,采用相邻网格法,将样地划分成100个10 m × 10 m的样方,在每个10 m × 10 m 的乔木样方内做2个 5 m × 5 m 的灌木样方和4个1 m × 1 m 的草本样方。运用坐标定位法对样地内木本植物(DBH≥1 cm)进行每木检尺。记录树种的种名、胸径(DBH)、树高和冠幅、生长状况及其样地中乔木的坐标(X,Y),同时记录各样方的海拔、坡度等环境因子。共记录乔木树种11种,共1 460株。样地概况见表1。
2.2 数据分析
2.2.1 乔木优势度的测度优势度=[相对多度(RAD%)+相对胸径(RDBH%)+相对高度(RHG)+相对频度(RFE%)]/4 (张金屯,2004)。
式中,RAD=Ai/∑Ai,RDBH=Di/∑Di,RHG=Hi/∑Hi,RFE=Fi/∑Fi。式中,Ai、Di、Hi、Fi分别代表某种植物的多度、胸径(cm)、高度(m)、频度;∑Ai、∑Di、∑Hi、∑Fi依次为所有植物的总多度、总胸径(cm)、总高度(m)、总频度。
2.2.2 径级结构分析由于树木的年龄检测比较困难,所以本研究以树木的胸径(DBH)作为表征树木年龄的指标,将胸径在2.5 cm以下的林木幼苗定义为DBH第Ⅰ级,其后以5 cm为步长增加一级,即2.5 cm≤ DBH<7.5 cm为第Ⅱ级,依次类推,总共将样地中的所有林木划分为8个径级,统计样地内各径级的株数。依据胸径径级,将林木划分为3个年龄段,分别为幼龄期(Ⅰ~Ⅱ级),中龄期(Ⅲ-Ⅴ级)和老龄期(Ⅵ-Ⅶ级)。
2.2.3 点格局(1)种群分布格局分析:RipleysK分析法中使用K作为统计函数(Ripley,1976)。K(r)被定义为以某一任意点为圆心,距离r以内的其余点数的期望值与样方内点密度的比值。其估计值如下:
K(r)=An2∑ni=1∑njw-1ijIi(uij)(1)
式中,A为样地面积,n为总点数(植物个体数),uij为点i和点j之间的距离,Wij为以点i为圆心,uij为半径的圆落在面积A中的比例。其为一个点(植株)可被观察到的概率,在这里引入它作为权重,以便消除边界效应(Edge effect)。通常用L(r)代替K(r)函数。其表达式如下:
L(r)=K(r)π-r(2)
若L(r)=0,呈随机分布;若L(r)>0,呈集群分布, 若 L(r)<0,呈均匀分布。L(r)值位于置信区间之上,种群呈聚集分布;L(r)值位于置信区间之下,种群呈均匀分布;L(r)值位于置信区间之内,种群呈随机分布。
(2)种间关系分析:两个物种之间的关系分析实际上是两个种的点格局分析,也叫多远点格局分析(Multivariate point pattern anylais)。
种间空间关系的公式如下:
K12r=An1n2∑ni=1∑njwiijIruij(3)
式中,n1和n2分别为种1和种2的个体数(点数),通常用L12(r)取代K12(r)。其公式如下:
L12r=K12rπ-r(4)
当L12(r)=0时, 表明两个种在t尺度下无关
联,当L12(r)>0时表明两个种为正关联, 当 L12(r)<0时, 表明两个种为负关联。当L12(r)值位于置信区间之上时,2个变量显著正相关;当L12(r) 值位于置信区间之下时,2个变量显著负相关;L12(r)值位于置信区间之内,2个变量相互独立。
数据分析使用Programita软件,对样地内主要树种的空间格局及其种间关系进行分析。采用空间尺度0~50 m,根据相应的零假设检验模型,进行99次Mentokalo (蒙特卡罗)模拟,得到上下包迹线构成的99%置信区间。利用 Excel 2010进行数据统计分析,利用Origin 9.0软件完成成图。
3结果与分析
3.1 群落特征
样地内共有木本植物76种41科71属,其中乔木11种,隶属于9科10属,灌木13种,隶属于6科12属,草本52种,隶属于28科42属。草本种类最多,灌木次之,乔木最少。属和种占优势的科主要有蔷薇科(6属6种)、毛茛科(5属6种)、唇形科(4属6种)、豆科(3属5种)、菊科(4属5种)和百合科(3属4种)。样地内木本植物科可划分为4个分布区类型,其组成以世界广布成分为主,同时混有泛热带、北温带和南温带间断分布及欧亚和南美洲温带间断分布等成分。属可划分为9个分布区类型,其中以北温带分布(30属)成分为主,旧世界温带分布(8属)、世界分布(7属)、泛热带分布(4属),东亚和北美洲间断分布(4属)、北温带和南温带间断分布(4属)等一些其他分布类型。
从表2可以看出,优势度排在前4位的辽东栎、槲树、油松和山杨是该群落的主要组成树种。其中辽东栎以483株的数量居于首位在该群落中占具优势;槲树的平均胸径低于总林木,说明槲树进入群落较晚,但个体数量排在第二,说明该区的气候环境条件适合槲树生长;油松的平均胸径和相对频度排在第二,但由个体数少于辽东栎和槲树,因此優势度排在第三;山杨的个体数量少于其他三个树种,但相对高度却大于其他三个树种,说明山杨的竞争能力更强。白桦的平均胸径、胸高断面积和相对高度都高于辽东栎,说明该样地中的白桦处于发育成熟期,但由于白桦个体数量较少,因此不占优势。五角枫和野山楂的个体数分别为4和5株,属于该林分的偶见种。其他树种的存在对稳定群落结构具有重要意义。
3.2 径级结构
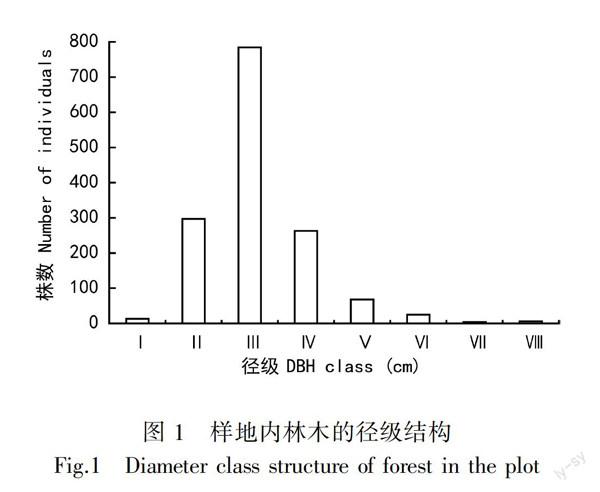
由图1可知,样地内植株径级呈近似正态分布,林分内中小径木数量居多且存在一定量的大径木。
以Ⅲ-Ⅴ级的个体数量最多,占总林分个体数的76.37%;胸径在Ⅰ-Ⅱ级个体数量为310株,占总林分个体数的21.23%;胸径在Ⅵ-Ⅶ级的个体数为29株,占总林分个体数的2.0%,胸径在Ⅷ级的个体数量最少,占总林分个体数的0.4%。由图2可知,主要树种的种群径级结构也都接近正态分布,个体数量都在第Ⅲ级出现峰值。辽东栎种群和油松种群的径级结构更为相似,在Ⅰ-Ⅷ级范围内都有分布,表明它们进入该群落的时间相对更早;槲树种群在Ⅵ和Ⅶ级个体数量较少,山杨种群只分布在Ⅱ-Ⅵ级,无大径级个体。
3.3 主要乔木树种的空间分布格局
选取优势度排在前4 位的辽东栎、槲树、油松、山杨进行种群的空间分布格局分析。主要树种的空间分布如图3所示。辽东栎个体数最多,在整个样地内均有较多分布,但中央分布较少;槲树的分布较集中于中部和左上部;油松在西南角聚集明显,其他地方表现为均为分布;山杨在样地内分布比较均匀。
主要树种的空间分布格局分析如图4所示。辽东栎种群和槲树种群的空间分布类型较一致。辽东栎种群在0~29 m的范围内呈显著性聚集分布,当r=7 m时,辽东栎聚集尺度最大,此时L(r)=0.818 55,r>29 m表现出随机分布格局;槲树种群在0~30 m的尺度范围内呈现出显著的集群分布,在大于30 m的尺度上呈现随机分布;油松种群在3~6 m、26~44 m尺度上呈现集群分布,当r=40 m时,聚集尺度达到最大,之后变为随机分布;山杨种群在0~17 m随机分布,17~42 m范围内呈现集群分布,当r=24 m时,L(r)=1.824 41聚集程度達到最大,r>42 m为随机分布。以上结果说明,辽东栎和槲树在小尺度为聚集分布,大尺度为随机分布,油松和山杨出现斑块性聚集的格局特征,这可能与种子散布、种内种间竞争有关。
3.3 主要乔木树种的空间关联性
主要树种的空间关系分析如图5所示。辽东栎与其他3个树种在整个尺度范围内表现相似的关联性,即随着尺度的增加由正关联变为无关联。如辽东栎和槲树在0~27 m的范围内两者正关联,在大于27 m的范围内两者无关联;辽东栎和油松在0~26 m的范围内两者正关联,在大于26 m的范围内两者无关联;辽东栎和山杨在0~31 m尺度范围内表现为正关联,在其他尺度范围内呈无关联;槲树与油松、山杨在整个尺度上都表现为显著正关联,表明它们之间关联性较大;油松和山杨,在8~32 m范围内呈正关联,在0~9 m和大于32 m的尺度呈无关联。以上结果表明,主要树种均在一定的尺度内表现出正相关,说明在一定的尺度范围内它们之间可能存在着某种互相依赖的关系或者有着相似的生态习性。
4讨论与结论
4.1 群落特征分析
样地内物种组成丰富,草本种类最多,灌木次之,乔木最少。科的分布区类型以世界广布为主,属的分布区类型以北温带分布为主,与柴宝峰和李毳(1997)的研究结果一致。优势度排在前四位的辽东栎、槲树、油松和山杨,这4个树种的相对多度之和为70%,而相对显著度之和达到78%,是该样地的主要组成树种。径级结构是衡量植物群落稳定性和生长发育状况的重要指标(农友等,2015)。整个样地内林木总径级结构与主要乔木种群的径级结构基本相似, 呈近似的正态分布,中龄期>幼龄期>老龄期,这种径级结构可以保证林分健康持续图 2样地内主要乔木种群径级结构 a. 辽东栎; b. 槲树; c. 油松; d. 山杨。下同。
4.2 主要树种的空间分布格局
格局分析是研究群落结构、种内种间关系以及生物与环境相互作用的重要手段(刘小恺等,2009)。种群空间分布格局受尺度大小的影响,在较小的尺度范围内可能受到种内或种间竞争、种子扩散限制等因素的影响,而在大尺度范围内可能受自身的生物学特性、种群分布区的异质性和外界无机环境等因素的影响。(张震等,2010;Harms et al,2001;Lin et al,2011)。本研究分析了针阔混交林样地内优势度排在前4位的辽东栎、槲树、油松和山杨的空间格局,结果充分体现了植物种群空间分布格局具有一定的尺度依赖性。辽东栎和槲树的分布格局类型大体相似,即在小尺度下为聚集分布, 随着尺度的增大转向随机分布的特征。造成这种分布格局的原因是辽东栎和槲树的种子是啮齿类动物的取食对象,动物取食和搬运等过程可能是导致种群在大尺度下呈随机分布的因素之一;油松种群在3~6 m和26~44 m的尺度上为聚集分布,这主要与种子扩散机制有关。由于油松种子质量大自然脱落以后不易扩散,散落在母株周围,进而表现出聚集分布的特征(张笑菁等,2010)。山杨在整个尺度上表现出随机分布-聚集分布-随机分布的格局特征,这可能是由种间竞争、人为干扰、种子散布等因素造成的斑块性或随机过程,这与段晓梅(段晓梅,2015)的研究结果一致。因此,太岳山南部针阔混交林主要树种的空间分布可能与种子扩散、自身的生物学特性及动物取食等密切相关,但如何准确判断某个因子的贡献大小还有待深入研究。
4.3 各种群的空间关联分析
分析同一群落内不同植物种群的空间分布格局及其空间关联性,对更深层次地理解植物本身的生态学特性及种间相互作用和群落稳定性变化具有重要意义(周先叶等,2000;闫海冰等,2010;Lan et al,2012)。不同树种间的正关联表明植物利用资源的相似性和生态位的重叠性,负关联性则表明树种间存在竞争关系,是对环境和空间资源利用相适应的结果,也是生态位分离的体现(张俊艳等,2014)。从本研究结果看,辽东栎与其他三个树种都在中小尺范围内表现为正相关,说明在中小尺度范围内它们互相依赖,共同发展,而在大尺度范围内表现为不相关,说明在大尺度上没有明显的种间竞争,生态位重叠较少。槲树与油松、山杨在整个尺度范围内都表现出显著正相关,油松和山杨在中尺度表现正相关,体现它们利用资源的相似性和较大的生态位重叠。因此,生态需求的异同可能是种间关联存在与否及关联性质的重要决定因素。
通常,群落演替初期,种间未形成特定的关系而表现为随机性(郭垚鑫等,2011),物种之间的关联程度往往较低,甚至产生较大的负关联,竞争较为激烈(郭忠玲等,2004),随着演替进行,群落种间关系逐渐趋向于正相关,物种组成和群落结构趋于完善和稳定,物种之间达到共存,从而发展为顶级群落。(邢福和郭继勋,2001)。主要树种均在一定的尺度范围内表现为正相关,说明太岳山南部针阔混交林中各个乔木种群处于相对稳定的状态,之后各物种会出现激烈的竞争,然后向顶级群落发展。由于处在中海拔地区,水热光照条件好而形成这种优势度不太明显的针阔混交林,但随着全球气候变暖,而温度的升高更有利于落叶阔叶树种的生长,辽东栎作为该群落优势度排在第一的树种,随着演替的进行,有可能发展为以辽东栎为建群种的群落。
本研究通过分析群落特征、主要树种的空间分布格局与空间关联性,提供了针阔混交林群落潜在生态学过程信息,说明各个种群处于相对稳定状态,由此稳定群落结构。该文只调查了一年的数据,今后还需通过长期观测来持续研究。可从优势种的更新、种内种间的共存关系,不同树种的年龄组成、群落演替地位与作用等方面来深入探究群落稳定性结构机制。
参考文献:
CAO J, BAI YH, ZHANG QD, et al, 2015. Interspecific association analysis of dominant species in Sambucus williamsii community in Taiyue Mountain of Shanxi Province [J]. Chin J Ecol, 34(5):1313-1319. [曹静,白玉宏,张钦弟,等,2015. 山西太岳山接骨木群落優势种种间关联性分析 [J]. 生态学杂志, 34(5):1313-1319.]
CHAI BF, LI C, 1997. Flora characteristics of seed plants in Taiyue Mountain of Shanxi Province [J]. Mount Res, 15(3):162-166. [柴宝峰, 李毳, 1997. 山西太岳山种子植物区系特征 [J]. 山地研究, 15(3):162-166.]
DRUCKENBROD DL, SHUGART HH, DAVIES I, 2005. Spatial pattern and process in forest stands within the Virginia piedmont [J]. J Veg Sci, 16(1):37-48.
DUAN XM, 2015. Study on population dynamics and spatial pattern of PoplarBirch Forests in Taiyue Mountain of Shanxi Province [D]. Linfen: Shanxi Normal University:1-51. [段晓梅, 2015. 山西太岳山杨桦林动态与空间格局分析 [D]. 临汾:山西师范大学:1-51.]
GREIGSMITH P, 1983. Quantitative plant ecology [M]. Oxford:Blackwell Scientific Publication: 54-104.
GUO DG, SHANG GUAN TL, BAI Z, et al, 2011. The ecological responses of Pinus tabulaeformis forests in Taiyue Mountains of Shanxi to artificial Harvesting [J]. Acta Ecol Sin, 31(12):3296-3307. [郭东罡, 上官铁梁, 白中, 等, 2011. 山西太岳山油松种群对采伐干扰的生态响应 [J]. 生态学报, 31(12):3296-3307.]
GUO KX, KANG B, LI G, et al, 2011. Species composition and point pattern analysis of standing trees in secondary Betula albosinensis forest in Xiaolongshan of west Qinling Mountains [J]. Chin J Appl Ecol, 22(10):2574-2580. [郭垚鑫, 康冰, 李刚, 等,2011. 小陇山红桦次生林物种组成与立木的点格局分析 [J]. 应用生态学报, 22(10):2574-2580.]
GUO ZL, MA YD, ZHENG JP, et al, 2004. Biodiversity of tree species, their populations spatial distribution pattern and inter specific association in mix eddeciduous broadleaved forest in Changbai Mountains [J]. Chin J Appli Ecol, 15(11):2013-2018. [郭忠玲, 马元丹, 郑金萍, 等,2004. 长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究 [J]. 应用生态学报, 15(11):2013-2018.]
HE FL, DUNCAN RP, 2000. Densitydependent effects on tree survival in an oldgrowth Douglas fir forest [J]. J Ecol, 88(4):676-688.
HUO MM, GUO DG, ZHANG J, et al, 2014. Tree community characteristics and spatial patterns of the forms Pinus tabuliformisQuercus wutaishanica in Lingkong Mountain [J]. Acta Ecol Sin, 34(20):5925-5935. [霍萌萌, 郭东罡, 张婕, 等, 2014. 灵空山油松-辽东栎林乔木树种群落学特征及空间分布格局 [J].生态学报, 34(20):5925-5935.]
HARMS KE, CONDIT R, HUBBELL SP, et al, 2001. Habitat associations of trees and shrubs in a 50ha neo tropical forest plot [J]. J Ecol, 89(6):947-959.
JOHN R, DALLING JW, HARMS KE, et al, 2007. Soil nutrients influence spatial distributions of tropical tree species [J]. Proc Nat Acad Sci, USA, 104(3):864-869.
JIN GY, ZHAO XM, KANG FF, et al, 2013. The great rainfall effect on soil respiration of Pinus tabulaeformis plantation in Taiyue Mountain [J]. Acta Ecol Sin, 33(6):1832-1841. [金冠一, 赵秀海, 康峰峰, 等, 2013. 太岳山油松人工林土壤呼吸对强降雨的响应 [J]. 生态学报, 33(6):1832-1841.]
JING X, DUAN WB, CHEN LX, et al, 2015. Spatial distribution pattern of main populations and gap makers in Picea koraiensis and Abies nephrolepis forest of Xiaoxingan Mountains, Northeast China [J]. Chin J Appl Ecol, 26(10):2928-2936. [景鑫, 段文标, 陈立新, 等, 2015. 小兴安岭谷地云冷杉林主要种群及林隙形成木的空间格局 [J]. 应用生态学报, 26(10):2928-2936.]
LI B, 2000. Ecology [M]. Beijing:Higer Education Press:45-47. [李博, 2000. 生态学 [M]. 北京:高等教育出版社:45-47.]
LIN C, MA QY, HAN HR, et al, 2002. Photosynthesis Characteristic of Quercus liaotungensis in Taiyue Mountain Region [J]. Acta Ecol Sin, 22(9):1339-1406. [蔺琛, 马钦彦, 韩海荣, 等, 2002. 山西太岳山辽东栎的光合特性 [J]. 生态学报, 22(9):1339-1406.]
LIU XK, LIU MS, HUANG Z, et al, 2009. Pattern analysis of interspecifc relationships in four arid communities in sand lake, Ningxia Hui Autonomous Region, China [J]. J Plant Ecol, 33(2):320-330. [刘小恺, 刘茂松, 黄峥, 等, 2009. 宁夏沙湖4种干旱区群落中主要植物种间关系的格局分析植物 [J]. 植物生态学报,33(2):320-330.]
LIN YC, CHANG LW, YANG KC, et al, 2011. Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation [J]. Oecologia, 165(1):175-184.
LAN G, GETZIN S, WIEGAN T, et al, 2012. Spatial distribution and inter specific associations of tree species in a tropical seasonal rain forest of China [J]. PLoS ONE, 7(9):e46074.
NONG Y, ZHENG L, JIA HY, et al, 2015. Community characteristics and spatial distribution of dominant tree species in a secondary forest of Daqing Mountains, south western Guangxi, China [J]. Biodivers Sci, 23(30):321-331. [農友, 郑路, 贾宏炎, 等, 2015. 广西大青山次生林的群落特征及主要乔木种群的空间分布格局 [J]. 生物多样性, 23(30):321-331.]
RIPLEY BD, 1977. Modeling spatial patterns [J]. J Royal Statist Soc Ser B, 39(2):172-212.
RIPLEY BD, 1976. The secondorder analysis of stationary point processes [J]. J Appl Probab, 13(2): 255-266.
WIEGAND T, MOLONEY KA, 2004. Rings, circles and nullmodels for point pattern analysis in ecology [J]. Oikos, 104(2):209-229.
WANG YL, ZHANG QD, HAO XJ, et al, 2012. Structure and spatial distribution of Acer ginnala population in Qiliyu,Shanxi [J]. Acta Bot BorealOccident Sin., 32(5):1027-1035. [王祎玲,张钦弟,郝晓杰,等, 2012. 山西七里峪茶条槭的种群结构与空间分布格局 [J]. 西北植物学报, 32(5):1027-1035.]
XIAO D, WANG XJ, ZHANG K, et al, 2016. Effects of nitrogen addition on leaf traits of common species in natural Pinus tabuliformis forests in Taiyue Mountain, Shanxi Province, China [J]. J Plant Ecol, 40 (7):686-701. [肖迪, 王晓洁, 张凯, 等, 2016. 氮添加对山西太岳山天然油松林主要植物叶片性状的影响 [J]. 植物生态学,40(7):686-701.]
XING F, GUO JX, 2001. Comparative analysis of interspecific association for three grazing successional stages of Cleistogenes squarrosa steppe [J]. J Plant Ecol, 25(6):693-698. [邢福, 郭继勋, 2001. 糙隐子草草原3个放牧演替阶段的种间联结对比分析 [J]. 植物生态学报, 25(6):693-698.]
YAN M, BI RC, ZHONG ZC, 2003. Investigation and study on plant resources of the Taiyue Moutains [J]. Terr Nat Resourc Study, (3)82-83. [闫明, 毕润成, 钟章成, 2003. 太岳山植物资源的调查与研究 [J]. 国土与自然资源研究, (3)82-83.]
YAN HB, HAN YZ, YANG XQ, et al, 2010. Spatial distribution patterns and associations of tree species in typical natural secondary mountain forest communities of Northern China [J]. Acta Ecol Sin, 30(9):2311-2321. [闫海冰, 韩有志, 杨秀清, 等, 2010. 华北山地典型天然次生林群落的树种空间分布格局及其关联性 [J]. 生态学报, 30(9):2311-2321.]
YI LT, HAN HR, CHENG XQ, et al, 2008. Spatial distribution patterns of Quercus liaotungensis population in Lingkong Mountains [J]. Acta Ecol Sin, 28(7):3254-3261. [伊力塔,韩海荣,程小琴,等, 2008. 灵空山辽东栎种群空间分布格局 [J]. 生态学报, 28(7): 3254-3261.]
ZHANG JT, MENG DP, 2004. Spatial pattern analysis of individuals in different ageclasses of Larix principisrupprechtii in Luya Mountain reserve, Shanxi , China [J]. Acta Ecol Sin, 24(1):35-40.
ZHANG JT, 2004. Quantitative ecology [M]. Beijing:Science Press:17-21. [張金屯, 2004, 数量生态学. 北京:科学出版社:17-21.]
ZHANG J, SHANG G TL, DUAN YH, et al, 2014. Age structure and dynamics of Quercus wutaishanica population in Lingkong Mountain of Shanxi Province,China [J]. Chin J Appli Ecol, 25(11):3125-3130. [张婕, 上官铁梁, 段毅豪, 等, 2014.灵空山辽东栎种群年龄结构与动态 [J]. 应用生态学报, 25(11):3125-3130.]
ZHANG XQ, ZHAO XH, KANG FF, et al, 2010. Spatial pattern of the trees in a natural Pinus tabulaeformis forest in Taiyue Mountain [J]. Acta Ecol Sin, 30(18) :4821-4827. [张笑菁, 赵秀海, 康峰峰, 等, 2010. 太岳山油松天然林林木的空间格局 [J]. 生态学报, 30(18):4821-4827.]
ZHANG Z, LIU P, DING Y, et al, 2010. Species compositions and spatial distribution pattern of tree individuals in the Schrenk spruce forest, northwest China [J]. J Nanjing For Univ (Nat Sci Ed), 34(5):157-160. [张震, 刘萍, 丁易, 等, 2010. 天山云杉林物种组成及其种群空间分布格局 [J].南京林业大学学报(自然科学版), 34(5):157-160.]
ZHOU XY, WANG BS, LI MG, et al, 2000. An analysis of inter specific associations in secondary succession forest communities in Heishiding Natural Reserve, Guangdong Province [J]. J Plant Ecol, 24(3):332-339. [周先叶, 王伯荪, 李鸣光, 等, 2000. 广东黑石顶自然保护区森林次生演替过程中群落的种间联结性分析 [J].植物生态学报, 24(3):332-339.]
ZHANG JY, CHENG KW, ZANG RG, 2014. The spatial distribution patterns and associations of the principal trees and shrubs in a natural tropical coniferous forest on Hainan Island, China [J]. Biodivers Sci, 22(2):129-140. [张俊艳, 成克武, 臧润国, 2014. 海南岛热带天然针叶林主要树种的空间格局及关联性 [J]. 生物多样性, 22(2):129-140.]
