基于戴云山固定样地黄山松群落物种组成与结构研究
2017-05-30刘艳会刘金福何中声洪伟冯雪萍崖佐朝
刘艳会 刘金福 何中声 洪伟 冯雪萍 崖佐朝

摘要: 黄山松是亚热带中部山区主要建群种,研究其物种组成及群落结构对探讨中亚热带植被演替规律具有重要意义。戴云山自然保护区分布有大面积黄山松群落,该研究采用网格布点法,建立86块(25.82 m × 25.82 m)共5.7 hm2固定样地,对样地内黄山松群落的物种组成特征、分布区类型、胸径和树高结构、空间分布格局4个方面特征进行了探讨。结果表明:(1)样地内共出现木本植物32 603株219种,隶属于59科108属。乔灌木优势种有黄山松、马尾松、木荷、肿节少穗竹、岩柃、窄基红褐柃等,其中稀有种和偶见种分别占总物种数的33.79%、25.75%和45.21%、24.66%。(2)在区系分布类型上,热带分布类型的科属多于温带分布类型的科属,热带分布类型中又以泛热带分布类型,温带分布类型以北温带占据最大比例。(3)样地内优势种的胸径分布呈倒“J”型,表明更新状态良好。垂直结构上,各优势种树高为倒“J”型或近似倒“J”型。黄山松和其他优势乔木大部分树高为4~10 m,灌木除短尾越桔0~1 m个体数最多外,其他优势灌木1~2 m个体数最多。(4)经计算,样地内乔灌木各优势种均呈聚集分布,聚集程度有所差异,乔木层黄山松最低,灌木层短尾越桔最低。该研究结果表明戴云山保护区黄山松群落物种组成丰富、更新良好,反映了亚热带高山针叶林的典型特征,其空间分布格局可能与取样方式等因素有关。
关键词: 黄山松群落, 物种组成, 胸径树高结构, 空间分布格局, 戴云山
中图分类号: Q948.15文献标识码: A文章编号: 10003142(2017)07088110
植物群落是一定地段内不同植物在长期历史过程中逐渐形成的生态复合体。分析植物群落可以揭示物种共存规律、形成机制,反应其生境条件以及空间关联。因此研究植物群落种类组成、数量特征、物种多样性是植被生态学研究的基础(刘静等,2016)。群落结构为揭示物种共存、群落演替和生物多样性维持机制等提供重要信息(Tilman et al,2006),群落的物种组成和区系成分则反映群落结构变化,是了解群落性质的基础和关键。黄山松(Pinus taiwanensis)是中国特有种,又称台湾松,常绿乔木,为喜光树种,耐干旱瘠簿、抗风,材质好,同时病虫危害少,生长持续时间长,主要分布于浙江、台湾、福建、江西、安徽等省山地,是亚热带中山地区的主要建群种。戴云山脉保存有中国大陆最南端、面积最大、保存最完好的原生性黄山松林,群落面积6 400 hm2,是中国最大的黄山松种质基因基地(朱德煌等,2012)。迄今,本课题组对戴云山黄山松群落已做大量研究,对其主要群落类型及物种多样性(任国学等,2011)、主要树种种群更新生态位(朱德煌等,2012)、幼苗与环境因子空间异质性关联(刘金福等, 2013;苏松锦等,2015)等进行了探讨分析,揭示了黄山松的生长环境和生存现状。由于黄山松群落广布于自然保护区内,而以往的研究局限于戴云山主峰或者九仙山、陈坂岭头等地,空间尺度相对较小。目前,我国建立了多个大型固定监测样地,对探讨物种共存机制、检验生态学理论提供了良好的平台(马克平,2008)。大型固定样地主要监测南亚热带常绿阔叶林、亚热带常绿阔叶林、暖温带落叶阔叶次生林以及温带阔叶红松林等(徐丽娜和金光泽,2012),对亚热带针叶林的研究仍较少。同时,大型样地设置多为监测某一区域内的阔叶林,因而海拔跨度小。针对分布范围广的群落进行研究时存在一定局限性。
基于上述考虑,以戴云山自然保护区为研究范围,采用网格布点法,扩大监测面积和海拔跨度,建立86块、总面积达5.7 hm2的黄山松样地。主要探讨以下几个科学问题:(1)戴云山自然保护区样地内黄山松群落的物种组成;(2)样地内出现物种所属区系;(3)优势种物种个体数,胸径树高分布有何特点;(4)黄山松群落优势种的空间分布格局。讨论不同取样条件下的异同,以期为戴云山黄山松群落的保护与发展提供基础数据支持。
1材料与方法
1.1 研究区及样地概况
戴云山自然保护区位于福建泉州德化县境内,118°05′22″~118°20′15″ E,25°38′07″~25°43′40″ N,属亚热带季风气候,年平均气温为15.6~19.5 ℃,最冷月在1月份,平均气温为6.5~10.5 ℃,最热月是七月,平均气温为23~27.5 ℃,最高气温36.6 ℃。空气相对湿度87%。山地黄壤、红壤、黄红壤是土壤主要组成部分。该区地形复杂,1 000 m以上的山峰有258座,其中最高峰海拔1 856 m,被誉为“闽中屋脊”。
戴云山属典型山地森林生态系统,保留有最为集中和完整、天然更新状态最好的黄山松林,且该地生物多样性丰富,样地内乔灌木层共出现种子植物59科108属219种;乔木层主要有黄山松、木荷(Schima superba)、马尾松(Pinus massonianaa)、红楠(Machilus thunbergii)、杉木(Cunninghamia lanceolata)、罗浮栲(Castanopsis fabri)等;灌木层主要有中华野海棠(Bredia sinensis)、肿节少穗竹(Oligostachyum oedogonatum)、华丽杜鹃(Rhododendron farrerae)、小叶赤楠(Syzygium buxifolium)、荚蒾(Viburnum dilatatum)等;草本层主要有芒萁(Dicranopteris pedat)、中华里白(Hicriopteris chinensis)、狗脊蕨(Woodwardia japonica)等。
1.2 樣地设置以及调查方法
2015年在戴云山自然保护区内(118.10°~11827° E, 25.65°~25.72° N之间)采用网格布点法以550 m间距公里坐标数点,如果横、纵坐标交点在黄山松小班内,则该点为样地位置。共设置25.82 m × 25.82 m的正方形样地86个(图1),总面积约为5.7 hm2。每个样地四个角点分别设置于北N、东E、南S、西W4个方向,样点中心位置O须埋设水泥桩,便于今后复查。记录每个样方内DBH >5 cm的乔木名称、冠幅、冠下高、生活力状况并按顺时针挂牌、编号;4个方向上分别设置4 m × 4 m 小样方,记录幼树(H>30 cm 和DBH<5 cm)名称、高度、胸径并编号,查数轮生枝数。记录灌木种类、相对盖度、株数、平均高;在每个灌木样方内设置一个1 m × 1 m 草本样方,记录种类、盖度、株数和平均高度。
1.3 数据处理
对调查数据进行统计分析计算样地内植物的相对密度、相对优势度、相对频度计算各物种重要值, 相关公式见孙儒泳等(2004)。根据世界种子植物科属分布区类型划分标准划分出所有植物所属科属的分布区类型(吴征镒,2003;吴征镒等,1997);通过计算Lloyd平均拥挤度、聚块指数、扩散系数以及负二项参数,对种群空间分布格局进行分析,相关定义及计算方法见文献(郑元润,1997)。
2结果与分析
2.1 植物组成特征
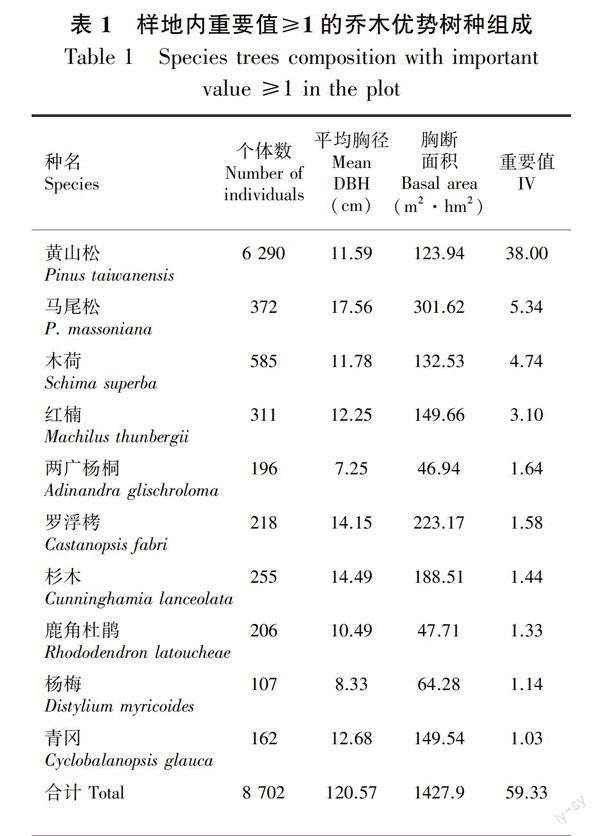
样地内黄山松群落分布有木本植物共59科108属219种。其中蔷薇科(22种)、壳斗科(20种)、樟科(18种)、冬青科(15种)等占据优势,主要优势树种包括乔木层的黄山松、马尾松、木荷、红楠、
两广杨桐、罗浮栲等,灌木层的肿节少穗竹、岗柃、窄基红褐柃、短尾越桔、满山红等。
样地内优势树种均为常绿种。Hubbell、Foster把单位公顷个体数少于等于1的种定义为稀有种,1~10的为偶见种(杨庆松等,2011)。黄山松群落固定样地中乔木层稀有种数(74种,占33.79%)低于灌木层(99种,占45.21%),灌木层偶见种数(54种,24.66)低于乔木层(56种,25.57%),乔木层只有1株物种的有25個,灌木层有46个。
由表1可知,平均胸径最大的树种为马尾松(17.56 cm),罗浮栲(14.15 cm)和杉木(14.49 cm)胸径均值较为接近,黄山松平均胸径为11.59 cm,由大到小排序,位于第7。胸高断面积排在前五位的树种为马尾松、罗浮栲、杉木、红楠和黄山松。所有乔木树种平均胸高断面积为144.85 cm2,胸高断面积之和为1 251 089 cm2,黄山松占总体胸高断面积的87.66%。
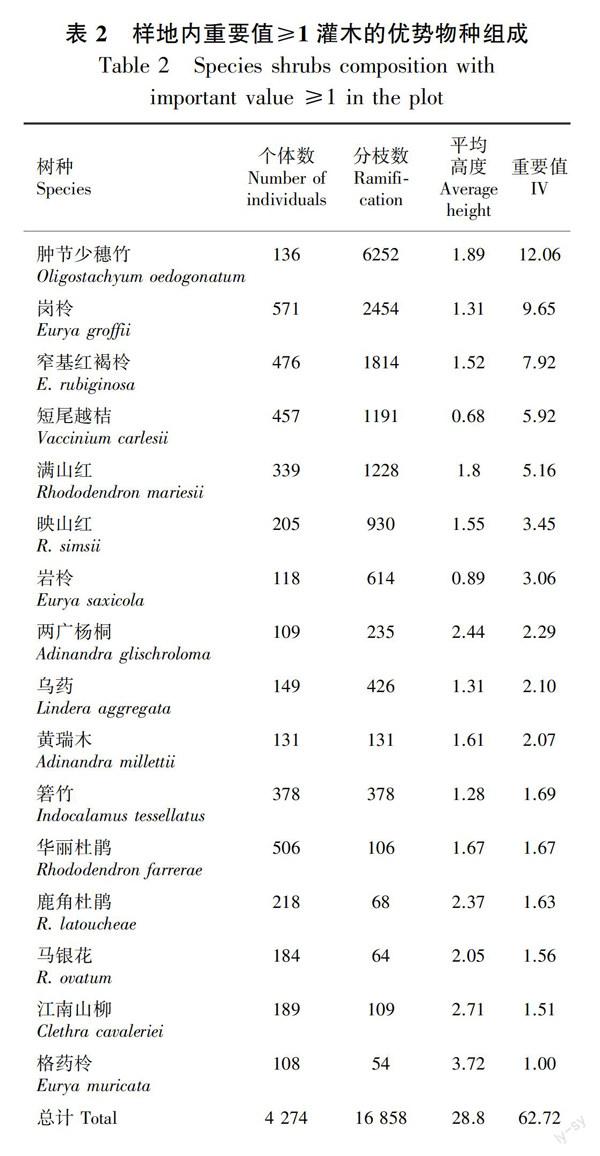
调查样地内重要值≥1的灌木有16种(表2),重要值最大的前三个物种为肿节少穗竹、岗柃、窄基红褐柃,依次为12.06、9.65、7.92。个体数量按从多到少排序前三的物种为岗柃、华丽杜鹃、窄基红褐柃。肿节少穗竹萌生枝数最多(6 252株)占总体的37%,岗柃(2454株)占14.56%,居其次。此16种灌木个体数占样地内灌木总个体数的80.02%。平均高度最大的是格药柃(3.72 m),江南山柳(2.71 m)和两广杨桐(2.44 m)次之。
2.2 分布区类型
由表3可知, 样地内热带区系类型和温带区系类型的科分别占总科数的74.58%和25.42%,其中热带区系类型的科以泛热带区系为主,主要是大戟科、樟科、山茶科等,占总科数的32.19%。温带区系类型的科中以北温带区系为主,占总科数的15.25%。样地内热带区系类型和温带区系类型的属分别占总数的52.78%和43.52%。热带区系类型中泛热带的属所占比例最高,主要包括大青属、冬青属、榕属等,占总属数的20.37%。温带区系属种以北温带分布类型的属数最多,包括李属、栎属、槭树属等,占总属数的20.37%。世界广布科有14科,占23.73%,主要包括桑科、榆科、蔷薇科等,属有4属。中国特有属包括箬竹属、杉木属、少穗竹属等。总体而言,热带分布区系类型科属多于温带分布区系类型科属。
2.3 径级结构
径级结构是植物群落稳定性和生长发育状况的重要指标。样地内共有乔灌木独立个体32 603株, 其中乔木层胸径最大的是东南石栎为111 cm,甜槠最大胸径62 cm居其次。样地内所有乔木平均胸径为10.19 cm。由图2可知,样地内黄山松和重要值前五位的树种胸径分布明显呈现倒“J”形,表明黄山松群落处于稳定与正常生长的状态。黄山松(5~7 cm) 和马尾松(5~11 cm) 初始径级个体数图 1黄山
并不是最多,分别在7~9 cm和11~17 cm处呈“峰形”分布,后呈倒“J”形。木荷、红楠、罗浮栲随着胸径变大,个体数逐渐减少, 而两广杨桐则急剧减少,其5~11 cm胸径的个体数占据了58.67%。根据实际测量,采取下限原则,将胸径划分为3个等级:小径木(5~10 cm)、中径木(10~20 cm)、大径木(>20 cm)。统计得知,样地DBH<20 cm的乔木个体数占样地内乔木总数的92.37%,小径木和中径木分别占48.43%和44.3%,大径木仅占7.54%,为最小比例。黄山松群落DBH<20 cm的占黄山松总体的94%,中径木所占比例大于小径木,接近半数。中小径级个体数和大径级个体数差异很大,表明戴云山目前生境适宜黄山松等优势树种的生存,且种群自我更新情况良好。
2.4 垂直结构
通过对样地内黄山松及其他优势树种的树高分类统计(图3)可以看出,样地内优势种树高个体数基本呈现峰形分布的凸形曲线。黄山松树高6~8 m个体数最多,其次为4~6 m个体数最多, 4~10 m黄山松占总体的75.44%。木荷各径阶树高个体数分布趋势与黄山松接近,4~10 m的个体数占总个体数的81.85%。红楠6~8 m个体数最多,占35.37%,两广杨桐 4~6 m个体数占总体的52.55%。罗浮栲各径级分布较平均。
灌木树高分布基本呈倒“J”形或近似倒“J”形。各个物种树高大部分在2 m以下。岗柃0~1 m和1~2 m个体数相当,窄基红褐柃1~2 m个体数最多。短尾越桔0~1 m个体数占最大比例。满山红和映山红分布较为平均,但1~2 m个体数仍占最大比例。
2.5 空间分布格局
种群空间格局反映的是一定环境因子对个体行为、生存和生产的影响,是分析种内和种间竞争、种子扩散、干扰等生态过程的基础(Druckenbrod et al,2005;Nathan,2006)。物种空间分布格局能够反映环境因子对植物个体生存、生产、更新的影响。通过对乔木和灌木的空间格局分析(表4,表5)可见,采用方差均值比率法,计算结果表明乔灌木各优势种均呈聚集分布。利用聚集强度指数进行分析验证发现丛生指数皆大于1,负二项参数较小,平均拥挤度远大于1,扩散系数大于1,可以推断乔木层优势树种呈集群分布。聚集程度前三位(由高到低)的是青冈、罗浮栲、木荷,黄山松的聚集程度最低,两广杨桐 、马尾松、红楠的聚集程度相近。灌木层优势种也呈集群分布,聚集程度高的物种是黄瑞木、岩柃、肿节少穗竹。聚集程度低的物种为岗柃和短尾越桔。
3讨论与结论
监测目的决定监测内容、指标和技术方法(徐海根等,2013)。固定样地的建立以黄山松群落本底调查为目的,用以监测整个保护区内黄山松群落物种组成、群落结构,以期揭示黄山松群落生境维持机制,在物种角度验证生态学假说。以大样地的方式取样跨度过大,实现难度高,局限于固定地块,对气候的反映不敏感。网格均匀布点取样,覆盖范围广,各样地间海拔、坡度、坡向等环境因素空间异质性强烈。与1 hm2大样地进行比较不仅能反映取样差异对物种多样性监测的影响,也能发现连续性样地和网格布点样地内黄山松群落维持机制的共性与差异。
3.1 物种多样性及区系类型
就物种多度而言,固定样地内木本植物达到59科108属219种,1 hm2样地物种多度为108种,隶属于34科61属, 物种数相差一倍。黄山松固定样地中稀有种和偶见种分别占33.79%和25.57%,与其他大样地(古田山、天童山、八大公山、武夷山)面积虽然不同,所占比例相似;金毛狗、福建柏、乐东拟单性性木兰、闽楠等国家二级保护植物,浙江桂、厚壳桂、多脉青冈等省级重点保护植物,长耳玉山竹、福建含笑等戴云山特有植物(郑世群,2013)也出现在样地中,而1 hm2样地中并未涵盖。可见面积增大, 物种数增加, 毋庸置疑。优势物種不同,1 hm2
样地中,乔木层优势树种较少,除黄山松外,仅有江南山柳和红楠,其他均为灌木。固定样地中优势种的数量明显增多,乔木层的马尾松、木荷、杉木、罗浮栲,灌木层的肿节少穗竹、乌药、黄瑞木、箬竹等物种出现,这与海拔跨度大有关。就保护区内主要群落而言,固定样地的研究结果更符合戴云山实际(郑世群等 2012;刘金福等 2013)。同时,也不难发现,无论取样地区、取样面积如何变化,岗柃、窄基红褐柃、两广杨桐、短尾越桔、鹿角杜鹃等物种在黄山松群落均有大量分布(朱德煌等,2012;苏松锦等,2015)。
区系特点来看,同纬度地区的大样地均表现出同一特点:热带分布的科属多于温带分布的科属,其中热带分布类型中以泛热带分布比例最大,温带分布科属以北温带数量最多。戴云山黄山松群落也表现出相同的特征,与其所处的地理位置相符。具体科属所占比例差异,是戴云山地处中亚热带南缘,地势较高、地形复杂,生境类型多样所致。总体上热带性质较显著,具有一定程度的南亚热带性质。
3.2 垂直结构和胸径分布
林木径级和树高分布结构是森林生态系统重要组成部分。戴云山黄山松群落大样地内乔木层和黄山松种群的径级树高都呈现倒“J”形(胸径低于20 cm,树高在4~10 m之间的黄山松占据主体)总体看来群落更新良好,与苏松锦等(2015)、朱德煌等(2012)研究结果一致。但在实际调查中发现,由于黄山松群落跨越海拔、区域尺度大,受自然环境资源异质性的影响,各小样地黄山松径级分布存在差异。高海拔(1 500~1 700 m)黄山松群落受到大风、土壤等自然因素影响,生长缓慢,同时由于光照充足,人为干扰少,其幼苗、幼树更新状况也比其他地区好,小径木数量多。海拔1 300~1 400 m黄山松纯林分布广,密度大,各个径级黄山松均有分布。林木个体之间处在水、肥、气、热和营养空间方面激烈竞争阶段,大径木较少,密度制约下枯木倒木也多分布于此区域。低海拔地区针阔混交,灌木茂盛,乔木层郁闭度大,随着气候的变化低海拔物种逐渐向较高海拔扩张,黄山松的适宜生境范围逐渐上移,海拔900 m处已逐渐减少,小径级居多。
大径木比例小是南方山区较为普遍的现象,就南方地区所建立的大型监测样地来看,面积虽有差异,但样地内小径木占比例均最大,可能是种群、种间竞争激烈,物种更替频繁,群落组成不稳定,处于受到一定程度干扰后的恢复演替阶段(丁晖等,2015)。人为干扰也是径级分布的重要影响因素,由于样地范围广,面积大,各地原因存在差异,部分黄山松群落是封山育林之后的天然次生林,生长年限短,树高、胸径偏小。DBH<10 cm的乔木和黄山松个体数均占各径阶的最大值,大径木和特大径木比例不到8%,黄山松甚至出现径阶缺失的现象,不排除靠近居民区的大径木被偷盗砍伐的可能性。因而,各地区林木小径级均占优势,而具体环境不同,未来演替方向也有待继续监测。
3.3 空间分布格局
种群空间分布格局依赖于研究尺度(苏松锦等,2015)。戴云山固定样地内黄山松群落乔灌木优势种均呈聚集分布,与苏松锦1 hm2样地研究结果一致。苏松锦等(2015)的研究认为,1 hm2样地内黄山松随机分布的地区是受地形影响,地形平坦、高程较低易出现均匀分布。在固定样地中,取样地形多种多样,黄山松群落亦为集群分布。Oring统计分析表明,0~26 m范围内黄山松群落乔木层会呈现集群分布的状态。固定样地内物种呈聚集分布可能是采样方法不同,单个样地尺度低于26 m所致。除取样差异外,物种本身所处演替阶段,及其群落物种特性也会是影响物种分布格局的因素(丁晖等,2015)。黄山松群落的径级树高分布状态表明群落仍在演替初期,即随着种群的发育,个体年龄和体积的增大,群落内物种对环境要求也将提高,自疏与他疏作用会使群落逐渐向均匀分布和随机分布过渡(伊力塔等,2008)。未来随着气候的变化,可能会对黄山松的种群格局产生负向影响。同时,野外调查发现,很多地区是荒废的耕地,部分乔木是在封山育林之后逐渐成长起来的,集群分布现象明显,表明人为因素也可能影响群落空间分布。就集群程度来看,鹿角杜鹃、格药柃、两广杨桐、马醉木是黄山松群落亚林层,为黄山松伴生树种,生境适应能力强,在戴云山广泛分布,集群程度高。红楠、杨梅、黄山松对海拔的适应状况不同,在900~1 700 m均有分布,集群程度没有其他树种高。黄瑞木生境适应能力强,肿节少穗竹、岗柃枝丛数多,聚集程度远高于其他灌木。集群程度也一定程度上反映了物种特性。
通过固定样地的建立对戴云山黄山松群落进行监测,在不同尺度范围下,黄山松种群如何维持,作为先锋树种其今后的演替方向如何,不同取样方式下监测结果是否相同,如何解释物种径级结构、空间分布格局上的差异,上述问题的回答,有待于做深入分析和探讨。固定样地物种组成和群落结构的分析,是研究黄山松种群物种共存机制、更新演替规律的第一步,也为生物多样性的监测和维持提供基础数据。
参考文献:
DRUCKENBROD DL, SHUGART HH, DAVIES I, 2005. Spatial pattern and process in forest stands within the Virginia piedmont [J]. J VEG Sci, 16(1):37-48.
DING H, FANG YM, YANG Q, et al, 2015. Community characteristics of a midsubtropical evergreen broadleaved forest plot in the Wuyi Mountains, Fujian Province, southeastern China [J]. Biodivers Sci, 23 (4):479-492. [丁暉, 方炎明, 杨青, 等, 2015. 武夷山中亚热带常绿阔叶林样地的群落特征 [J]. 生物多样性, 23 (4):479-492.]
LIU J, MA KM, QU LY, 2016. Species composition and community structure of dominant mangrove forests in Zhanjiang Mangrove National Nature Reserve, Guangdong Province [J]. Ecol Sci, 35(3): 1-7. [刘静, 马克明, 曲来叶, 2016. 广东湛江红树林国家级自然保护区优势乔木群落的物种组成及结构特征 [J]. 生态科学, 5(3): 1-7.]
LIU JF, ZHU DH, LAN SR, et al, 2013. Association between environment and community of Pinus taiwanensis in Daiyun Mountain [J]. Acta Ecol Sin, 33(18):5731-5736. [刘金福, 朱德煌, 兰思仁, 等, 2013. 戴云山黄山松群落与环境的关联 [J]. 生态学报, 33(18):5731-5736.]
MA KP, 2008. Large scale permanent plots: important platform for long term research on biodiversity in forest ecosystem [J]. Acta Phytoecol Sin, 32(2): 237-237. [马克平, 2008. 大型固定样地:森林生物多样性定位研究的平台 [J]. 植物生态学报, 32(2): 237-237.]
NATHAN R, 2006. Longdistance dispersal of plants [J]. Science, 313(5788):786-788.
REN GX, LIU JF, XU DW, et al, 2011. Analysis on classification and species diversity of Pinus taiwanensis community in Daiyun Mountain National Nature Reserve [J]. J Plant Resour Environ, 20(3):82-88. [任国学, 刘金福, 徐道炜, 等, 2011. 戴云山国家级自然保护区黄山松群落类型与物种多样性分析 [J]. 植物资源与环境学报, 20(3):82-88.]
SUN RY, LI QF, NIU CJ, et al, 2002. Basic ecology [M]. Beijing: Higher Education Press:140-143. [孙儒泳, 李庆芬, 牛翠娟, 等, 2002. 基础生态学 [M]. 北京:高等教育出版社:140-143.]
SU SJ, LIU JF, MA RF, et al, 2015. Spatial distribution patterns and associations of Pinus taiwanensis population in Daiyun Mountain, Southeast China [J]. Source Sci, 37(4):841-848. [苏松锦, 刘金福, 马瑞丰, 等. 戴云山黄山松种群的空间分布格局与关联性 [J]. 资源科学, 37(4):841-848.]
TILMAN D, REICH PB, KNOPS JMH, 2006. Biodiversity and ecosystem stability in a decadelong grassland experiment [J]. Nature, 441(7093): 629-632.
WU ZY, ZHOU ZK, LI DZ, et al, 1997. The arealtypes of the world families of seed plants [J]. Plant Divers Res, 25(3):245-257. [吴征镒, 周浙昆, 李德铢, 等, 1997. 世界种子植物科的分布区类型系统 [J]. 云南植物研究, 25(3):245-257.]
WU ZY, 2003. Revision of the arealtypes of the world families of seed plants [J]. Plant Divers Res, 25(5):535-538. [吴征镒, 2003. 《世界种子植物科的分布区类型系统》的修订 [J]. 云南植物研究, 25(5):535-538.]
XU LN, JIN GZ, 2012. Species composition and community structure of a typical mixed broadleavedKorean pine (Pinus koraiensis) forest plot in Liangshui Nature Reserve, Northeast China [J]. Biodivers Sci, 20 (4): 470-481. [徐麗娜, 金光泽, 2012. 小兴安岭凉水典型阔叶红松林动态监测样地:物种组成与群落结构, 生物多样性 [J]. 20 (4): 470-481.]
XU HG, DING H, WU J, et al, 2013. Principles, indicators and sampling methods for species monitoring [J]. Acta Ecol Sin, 33(7) : 2013-2022. [徐海根, 丁晖, 吴军, 等, 2013. 生物物种资源监测原则与指标及抽样设计方法 [J]. 生态学报, 33(7) :2013-2022.]
YANG QS, MA ZP, XIE YB, et al, 2011. Community structure and species composition of an evergreen broadleaved forest in Tiantongs 20 ha dynamic plot, Zhejiang Province, eastern China [J]. Biodivers Sci, 19 (2):215-223 [杨庆松, 马遵平, 谢玉彬, 等. 2011. 浙江天童20 ha常绿阔叶林动态监测样地的群落特征 [J]. 生物多样性, 19 (2):215-223.]
YI LT, HAN HR, CHENG XQ, 2008. Spatial distribution patterns of Quercus liaotungensis population in Lingkong Mountains [J]. Acta Ecol Sin, 28(7):3254-3261. [伊力塔, 韩海荣, 程小琴, 2008. 灵空山林区辽东栎(Quercusliaotungensis)种群空间分布格局 [J]. 生态学报, 28(7):3254-3261.]
ZHENG YR, 1997. The applicability of various methods in analysis of Picea mongolica population spatial distribution pattern [J]. Acta Pytoecol Sin, 21(5):480-484. [郑元润, 1997. 不同方法在沙地云杉种群分布格局分析中的适用性研究 [J]. 植物生态学报, 21(5):480-484.]
ZHU DH, LIU JF, HONG W, et al, 2012. Studies on regeneration niche of main species in Pinus taiwanensis community in Daiyun Mountain [J]. J Trop Subtrop Bot, 20(6):561-565. [朱德煌, 刘金福, 洪伟, 等, 2012. 戴云山黄山松群落主要树种更新生态位研究 [J]. 热带亚热带植物学报, 20(6):561-565.]
ZHENG SQ, LIU JF, HUANG ZS, et al, 2012. Nutrition ecological niche of dominant arbor species in Castanopsis fabri forest in Daiyun Mountains [J]. J Trop Subtrop Bot, 20(2):177-183. [郑世群, 刘金福, 黄志森, 等, 2012. 戴云山罗浮栲林主要乔木树种营养生态位研究 [J]. 热带亚热带植物学报, 20(2):177-183.]
ZHENG SQ, 2013. Studies on plant diversity and its evaluation of Fujian Daiyun Mountain National Nature Reserve [D]. Fuzhou: Fujian Agriculture and Forestry University: 1-140. [郑世群, 2013. 福建戴云山国家级自然保护区植物多样性及评价研究 [D]. 福州: 福建农林大学: 1-140.]
