太平湖浮游动物动态演替与环境因子的相关性研究
2017-05-16冯世敏吴明姝熊莲李东京王俊莉刘冬燕
冯世敏吴明姝熊 莲李东京王俊莉刘冬燕,
(1. 上海师范大学环境科学系, 上海 200234; 2. 上海师范大学城市生态与环境研究中心, 上海 200234)
太平湖浮游动物动态演替与环境因子的相关性研究
冯世敏1吴明姝1熊 莲1李东京1王俊莉1刘冬燕1,2
(1. 上海师范大学环境科学系, 上海 200234; 2. 上海师范大学城市生态与环境研究中心, 上海 200234)
2012年11月至2014年10月, 对太平湖浮游动物群落进行了为期两年的调查研究。共鉴定出浮游动物45属89种, 其中轮虫 29属69种、枝角类5属7种、桡足类2属4种和原生动物9属9种; 优势种主要来自于轮虫异尾轮虫属(Trichocereca)和龟甲轮虫属(Keratella)。浮游动物的丰度值存在明显的季节变化, 表现为夏季最大,平均达1326 ind./L, 秋季春季次之, 分别为608和605 ind./L, 冬季最小为216 ind./L; 垂直分布表现为春夏季太平湖表层浮游动物丰度最高, 中间层次之, 底层最小, 秋冬季则表现为中间层最高。浮游动物群落Shannon-Wiener多样性指数和Margalef丰富度指数中间层普遍高于表层和底层, Pielou均匀度指数表现为底层要高于表层和中间层, 季节变化表现为夏秋季显著高于冬春季的现象, 水质评价表明夏秋季水质好于春冬季。聚类和多维尺度分析表明: 太平湖浮游动物可分为夏秋季类群与春冬季类群, 两类群均表现为湖心与上下游区域群落结构差异较大, 其中春冬季类群差异较明显; 相关和逐步回归分析表明: 透明度和水温为太平湖浮游动物群落结构变化的主要环境影响因子; 依据结构方程模型(SEM)和冗余分析(RDA)的结果显示, 在溶解氧和水温较高的水环境中浮游动物丰度值表现为较大, 其中水温对轮虫的影响高于对枝角类和桡足类的影响。
太平湖; 浮游动物; 相关性分析; 冗余分析; 结构方程模型(SEM)
浮游动物是水生生态系统中的重要组成部分,具有个体微小、数量多、代谢活动强等特点, 多以浮游植物、腐屑、细菌等为食, 其本身又为浮游动物食性鱼类提供饵料, 在水生生态系统中起承上启下的作用, 是水生生态系统能量流动和物质循环的重要环节, 在组成食物链、调节湖泊自净能力和保持湖泊生态平衡等方面起着重要作用[1—3]。此外,浮游动物与水质质量关系密切, 有些种类对环境变化反应很敏感, 水质的任何变化都可能影响它们的生理功能、种类丰度、群落结构等[4,5], 因此, 浮游动物的群落组成、丰度、优势种、生物多样性等与水体的营养程度密切相关, 是水质污染及营养水平的重要指标, 从而为水生生态系统健康评估提供很好的参数[6—8]。掌握浮游动物群落结构特征对研究渔业资源开发、水产养殖业方面也具有重要意义[1]。基于这些特点, 浮游动物群落的研究已成为了水生生物学研究的重要内容之一。
太平湖流域地处安徽省黄山市境内, 位于东经117°28′—118°21′, 北纬30°0′—30°32′, 1970年建成,流域面积2800 km2, 库长约50 km, 最大水深约70 m,平均水深约40 m, 是安徽省最大的水库型湖泊, 在2012年被列入国家第二批生态环境保护专项, 2014年又成为全国15个重点保护湖泊之一。最近几年, 关于安徽省太平湖生态环境的调查研究较少,其中“八五”期间中国科学院水生生物研究所对太平湖进行了水质特征、浮游生物的调查[9,10], 近两年陆林和刘亚凤等[11,12]又分别对太平湖旅游地演化过程和底质沉积物多种重金属分布特征时行研究, 而对太平湖浮游动物群落变化研究甚少。鉴于对太平湖生态环境保护的重要性, 其生态本底调查及研究急需拓展, 本研究对安徽太平湖进行了为期两年的调查, 利用浮游动物种类组成、丰度、生物量、优势种和生物多样性指数等指标, 结合聚类分析及冗余分析探讨了太平湖浮游动物的群落结构及时空动态变化, 分析现阶段太平湖的水质状态及其影响浮游生物群落结构稳定性的因素, 并利用多种指标对太平湖水库的水生态环境进行评价, 旨在为太平湖的水生态环境保护提供基础数据和参考资料。
1 材料与方法
1.1 采样点布设和采样时间
2012年11月—2013年9月期间每隔2个月对太平湖进行一次采样, 时间分别为2012年11月和2013年1月、3月、5月、7月和9月, 2013年9月—2014年10月期间按每个季度采样一次, 分别为2013年12月和2014年4月、7月和10月。从上游至下游共设5个采样点, 分别记为H1、H2、H3、H4、H5 (图 1)。
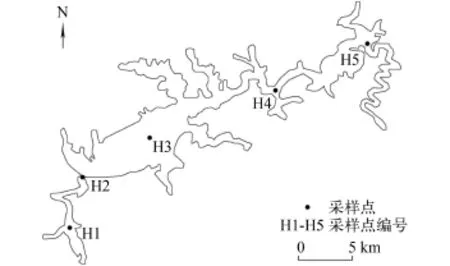
图 1 太平湖采样点布设图Fig. 1 The distribution of sampling sites in the Taiping Lake
1.2 水质理化指标的测定
水质检测项目有透明度(SD)、水深(D)、溶解氧(DO)、水温(T)和叶绿素a (Chl.a)。SD利用萨氏盘测量; DO利用HQ30D53便携式溶氧仪在现场测定; Chl.a样品现场抽滤后闭光保存带回实验室, 依照国标水质叶绿素的测定分光光度法(SL 88-2012)测定Chl.a的浓度。
1.3 样品采集与处理
用25号浮游生物网于水面以下约0.5 m处, 反复做“∞”型拖曳约5—10min, 提出水面得到浓缩样,滴加1 mL 4%甲醛溶液固定保存, 带回实验室进行定性分析。用5 L的有机玻璃采水器分别采集上(水面下0.5 m处)、中、下(距水底0.5 m处)三层各5 L水(共15 L), 加入75 mL鲁哥试液固定保存, 静置24h后, 经沉淀浓缩至约50 mL, 带回实验室进行定量分析。
种类鉴定主要参考王家楫、周凤霞和Koste等[13—15]。计数时充分摇匀定量样品, 用移液枪吸取1 mL样品, 注入1 mL的计数框中, 在光学显微镜10×10倍视野下进行全片计数, 计数2 次, 取平均值。
1.4 浮游动物群落结构参数的计算
将得到的数据分类整理, 依据以下公式换算成浮游动物的丰度, 描述浮游动物群落特征的参数采用Shannon-Wiener多样性指数(H′)、Margalef多样性指数(d)和Pielou均匀度指数(J′)。
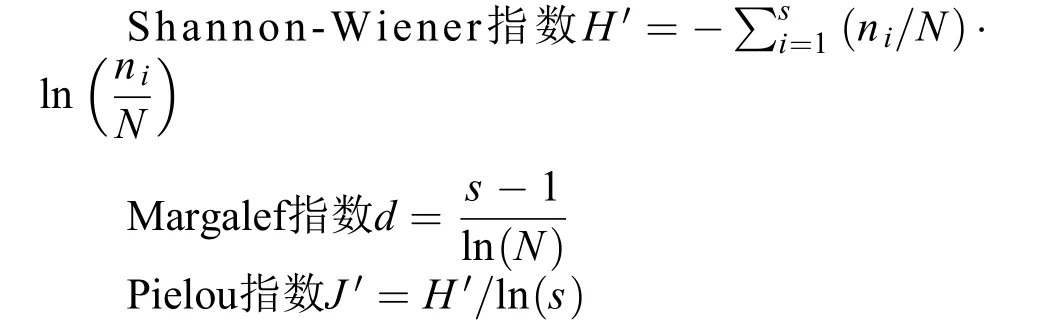
式中, ni为第i种浮游动物个体数, N为1 L水中浮游动物的个数(ind./L), S为浮游动物种数。
1.5 数据的统计学处理
采用SPSS19.0 统计软件进行相关、逐步回归分析、聚类和多维尺度分析以及非参数统计检验中的独立样本T检验和克鲁斯凯-沃利斯方差分析,冗余分析的统计学计算采用Canoco4.5软件处理,结构方程模型(Structure Equation Modeling, SEM)采用AMOS软件进行分析, 所有分析都以0.05作为显著性置信限。
2 结果
2.1 理化指标时空分布
各样点水温与溶解氧无明显变化, 水温19.1—19.7℃, 溶解氧7.47—8.75 mg/L (图 2)。太平湖水温与溶解氧存在着显著的季节变化(图 3), 水温在夏秋季最高25.2—27.7℃, 春冬季较低7.9—12.9℃。
水深与透明度从H1到H5即上游到下游依次递增, 上游H1最浅(15±3.2) m, 透明度为(2.2±0.4) m,下游H5最深达到(48.9±0.5) m, 透明度为(3.9±1) m,溶解氧百分比与浮游植物叶绿素a含量从上游至下游递减趋势非常明显(图 2)。由于季节的影响, 水深、透明度、溶解氧百分比和浮游植物叶绿素a都存在显著的时间变化(图 3), 夏季水位最高, 平均值为29.7 m, 冬季水位最低, 平均值为20.6 m。透明度则相反表现为春冬季高于夏秋季, 但变化幅度不大。溶解氧百分比春夏季高于秋冬季, 5月份左右达最大值(118.1±9.7)%, 9、10月份达到最低为(62.5±9.3)%, 与叶绿素a变化相反, 叶绿素a在9、10月出现峰值为(14.28±9.03) μg/L, 5月左右出现谷值为(3.03±1.13) μg/L。
2.2 浮游动物群落结构组成与分布
调查周期内, 全湖共检测到浮游动物45属89种(表 1), 隶属于轮虫、枝角类、桡足类和原生动物。其中, 轮虫最多, 共计29属69种, 占种类组成的77.5%, 占绝对优势; 原生动物次之, 共计9属9种, 占种类组成的10.1%; 枝角类共计5属7种, 占种类组成的7.9%; 桡足类最少共计2属4种, 占种类组成的4.5%。通过公式计算, 利用优势度(Y>0.02)筛选出浮游动物的优势种, 结果共检测到优势种32种, 主要是以轮虫为主共23种, 原生动物4种、桡足类3种和枝角类2种, 其中轮虫中的异尾轮虫属(Trichocereca)和龟甲轮虫属(Keratella)占优势。
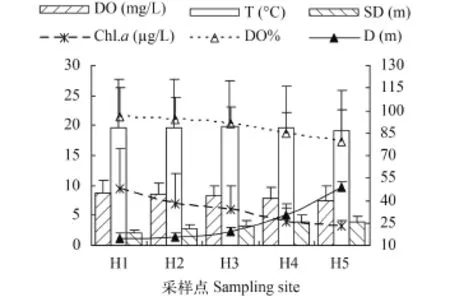
图 2 各调查站位理化指标的周年平均值Fig. 2 Annual averages of physical and chemical index in each site
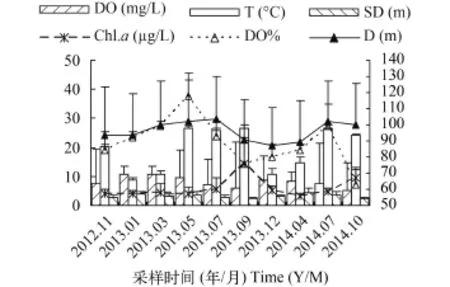
图 3 各理化指标平均值时间变化Fig. 3 Annual variation of the verages of physical and chemical index
浮游动物种类时空分布 太平湖浮游动物中主要以轮虫为主, 从上游H1到下游H5轮虫变化呈现先降后增再降的变化规律, 其余各浮游动物种类数无明显变化(图 4), 其中上游H1和下游H4区域的种类数较多, 全年累计出现种类数据分别达72种和66种, 而中部地区H2累计出现种类最少, 为56种。
调查期间太平湖浮游动物季节变化较明显(图5), 轮虫表现为夏秋季明显高于冬春季, 这可能由于水温对轮虫的生长发育有着重要影响, 轮虫变化与水温变化趋势基本一致(图 3、图 5), 对其进行相关分析得到相关系数R=0.873 (P<0.01), 轮虫与水温呈极显著相关。而原生动物夏季最高, 秋季则未检测出, 枝角类与桡足类季节变化不明显, 随着太平湖时间演替枝角类有所增加(图 5)。从优势种来看春季主要以疣毛轮虫属(Synchaeta)和多肢轮虫属(Polyarthra)为主要优势种, 夏秋季主要以异尾轮虫属(Trichocereca)和龟甲轮虫属(Keratella)为主要优势种, 冬季则以疣毛轮虫属(Synchaeta)和无节幼体(Nauplii)为优势种。
浮游动物丰度与生物量动态分布 太平湖浮游动物的丰度值在各采样点均呈明显的季节变化, 表现为夏季最大, 平均值达1326 ind./L, 秋季春季次之, 分别为608和605 ind./L, 冬季最小, 为216 ind./L, 这与浮游动物组成季节变化规律相同。总体上从上游H1到下游H5浮游动物丰度呈递减趋势, 周年变化为200—1531 ind./L, 年平均值为689 ind./L。从生物量角度分析, 其与丰度季节变化有所不同, 从上游H1至湖中心H3区域最大峰值出现在春季, 平均值为2.5 mg/L, 下游H4至H5区域则秋季最大, 平均值为2.9 mg/L。整个太平湖平面分布从上游到下游春夏季依然呈递减趋势与丰度变化相同, 而秋冬季呈正V字形即湖中心H3点最小(图 6)。
由图 7所示垂直分布表现为在春夏季时太平湖的表层浮游动物丰度最高, 中间层次之, 底层最小,如2013年7月表层平均值达2981 ind./L, 中间层为2155 ind./L, 底层为1578 ind./L, 秋冬季时则表现为中间层最高如2014年10月平均值为563 ind./L。总体分析, 在浮游动物群落演替过程中丰度有所下降, 而各层浮游动物生物量演替规律则基体保持一致。
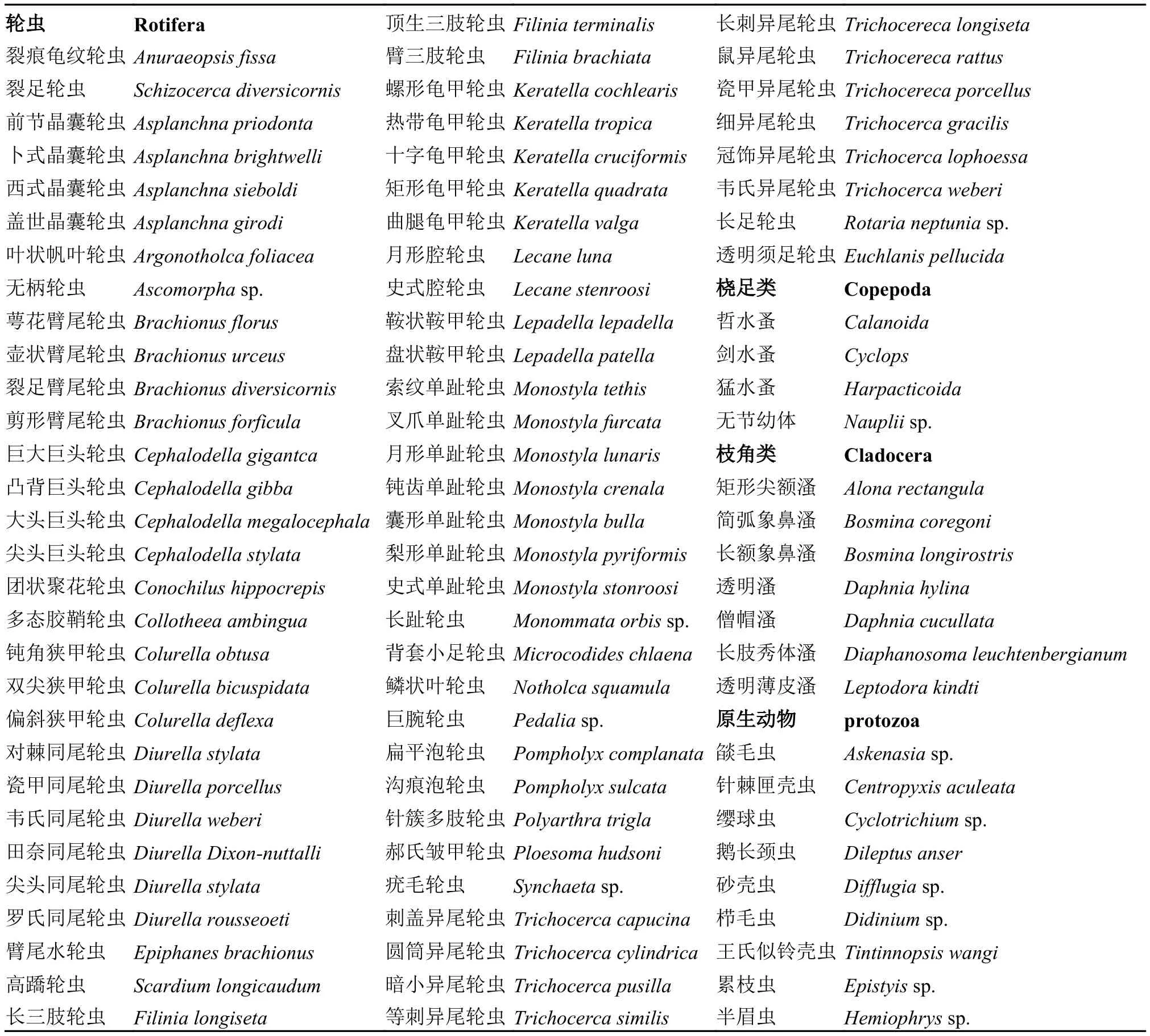
表 1 太平湖中的浮游动物种类Tab. 1 The zooplankton species found in the Taiping lake
浮游动物优势种变化 由图 8、图 9看出,在整个调查期间各浮游动物优势种占总优势种比例表现为轮虫最大, 所占比例达71.9%, 原生动物次之为12.5%, 桡足类占9.4%, 枝角类最少占6.3%, 说明安微太平湖浮游动物主要是以轮虫为主要优势种。从季节变化看, 春季除轮虫外主要以桡足类,夏季以原生动物类为优势种, 在秋季时优势种主要以轮虫类和桡足类为主, 原生动物类则未出现。从水平分析看优势轮虫类所占比例表现为上下游高于湖中心区域, 枝角类作为优势种主要出现在湖中心区域H2和H3处, 桡足类从H1到H5所占比例先增后减, 说明太平湖湖中心区域桡足类作为优势种高于上下游, 原生动物类则相反表现为上下游所占比例高于湖中心区域(图 9)。总量上分析优势种类存在显著周期性, 每年的9、10月份出现一次峰值, 3、4月份出现谷值即秋季优势种类最多春季最少,这与叶绿素a的变化规律相同, 平面分析表现为H2优势种类最多, 从H2到H5依次减少, 整体呈倒V字形, 即太平湖中部区域优势种类数大于上下游区域。
2.3 浮游动物群落结构参数
太平湖不同采样时间不同层次的浮游动物群落结构参数(物种Margalef丰富度指数、Shannon-Wiener多样性指数和Pielou均匀度指数)见表 2。
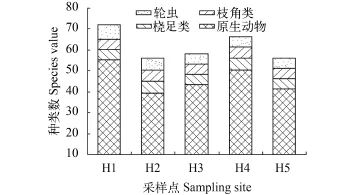
图 4 浮游动物种类的水平变化Fig. 4 The region distribution of the zooplanktons
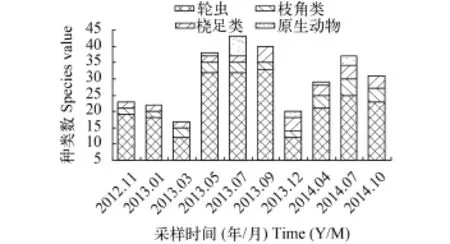
图 5 太平湖浮游动物时间分布Fig. 5 Annual distribution of zoopanktons in Taiping Lake
图 6 浮游动物群落丰度与生物量季节变化Fig. 6 The abundance and biomass seasonal changes of zooplankton communities at different site

图 7 演替过程中浮游动物丰度与生物量垂直分布Fig. 7 The vertical distribution of abundance and biomass of zooplanktons in the succession

图 8 太平湖优势种时间分布百分比Fig. 8 The proportion of time distribution of dominant species in the Taiping Lake
由表 2可以发现, 太平湖不同层次浮游动物的Margalef丰富度指数(d)介于0.31—2.03, 平均值为1.48; Shannon-Wiener多样性指数(H′)介于0.73—2.44, 平均值为1.56; Pielou均匀度指数(J′)介于0.40—2.33, 平均值为0.70。综合考虑各层次, 中间层的Shannon-Wiener多样性指数和Margalef丰富度指数普遍高于表层和底层, Pielou均匀度指数则表现为底层要高于表层和中间层。各多样性指数季节变化表现为夏秋季显著高于冬春季, 其中Shannon-Wiener多样性指数和Pielou均匀度指数秋季最高,平均值分别为2.05和0.82, 春季最低平均值分别为1.10和0.60, 而Margalef丰富度指数则表现为夏季最高冬季最低, 其值分别为1.95和0.96, 3种多样性指数季节差异均非常显著(F=1.812, P=0.000; F= 0.590, P=0.002; F=0.352, P=0.000)。
2.4 浮游动物群落的时空划分
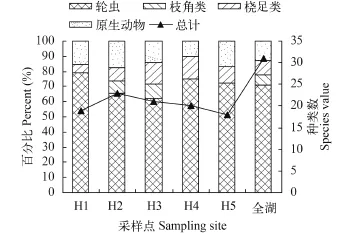
图 9 太平湖优势种水平分布百分比Fig. 9 The proportion of region distribution of dominant species in the Taiping Lake
以不同季节不同采样点获得的浮游动物样品的丰度、生物量和多样性指数以及各理化因子为变量采用欧式距离进行系统聚类(图 10), 结果显示20个样点的浮游动物大致聚类成夏秋季类群和春冬季类群。夏秋季类群浮游动物种类数和丰度值较高, 多样性指数也较高, 而春冬季类群种类数少,丰度低, 多样性指数也较低。在夏秋季类群内上游与湖中心区域各采样相似性较高, 而春冬季类群再现湖中心区域与下游各采样点相似性较高。
对两个季类群分别进行多维尺度分析并建立MDS排序图, 图 10A为夏秋季类群各样点MDS排序图其Stress值为0.0008小于0.005, RSQ为1.000,图 10B为春冬季类群各样点MDS排序图Stress值为0.0015, RSQ为1.000, 表明MDS排序图能很好地显示在同一组浮游动物的相似性和不同组的差异性[16]。夏秋季类群大体可分为四类, 上游H1各层群落相似性较大, 中上游的表层与中间层浮游动物群落结构相似, 而中下游底层群落结构较相似, 总体群落分布较为均匀。春冬季类群与夏秋季类群有所不同, 主要集中在湖心区域, 上游各层春季群落相似性也较差, 总体表现为湖心与下游群落相似性较大, 在浮游动物现存量和多样性指数上都比夏秋季类群低, 这与聚类分析结果相同。
2.5 浮游动物群落的理化因子分析
浮游动物与环境因子相关性 通过Spss19统计分析软件将太平湖浮游动物种类数、丰度和生物量与各环境因了进行相关分析(表 3), 浮游动物丰度与水深和透明度呈极显著负相关, 而与水温、溶解氧百分比和叶绿素a呈显著正相关; 生物量则与透明度呈负相关; 浮游动物种类种则与透明度和溶解氧呈极显著负相关与水温与叶绿素a呈极显著正相关。通过多元逐步回归分析分别筛选出影响浮游动物丰度、生物量和种类数的主要因子,结果表现太平湖透明度(SD)和温度(T)两个因子入选了浮游动物丰度回归方程, 方程为 Y=–248.894 (SD)+23.262 (T)+1073.319 (P<0.01); 浮游动物生物量的回归方程只有透明度入选; 而浮游动物种类数与环境因子的回归方程中透明度、溶解氧、溶解氧百分比和叶绿素a四个环境因子入选, 其方程为Y=–2.369 (DO)–0.876 (SD)+0.211 (DO%)+0.173 (Chl.a)+12.526 (P<0.05)。从三个回归方程看出透明度和水温为太平湖浮游动物群落结构变化的主要环境影响因子。
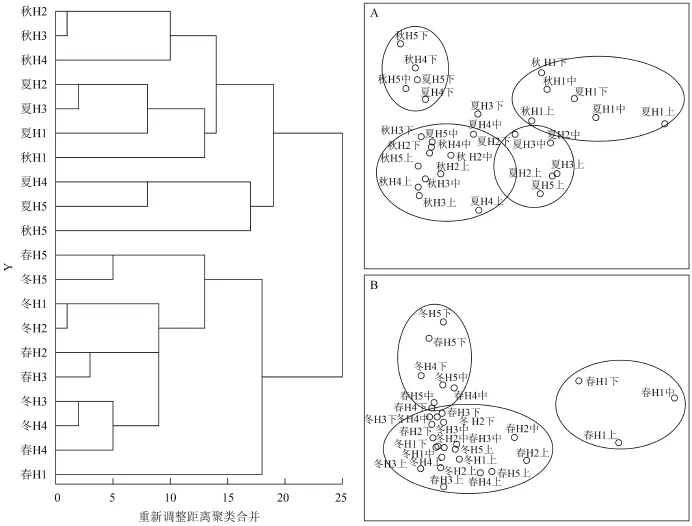
图 10 太平湖浮游动物群落系统聚类和多维尺度分析Fig. 10 The cluster analysis and MDS of zooplankton community in the Taiping Lake
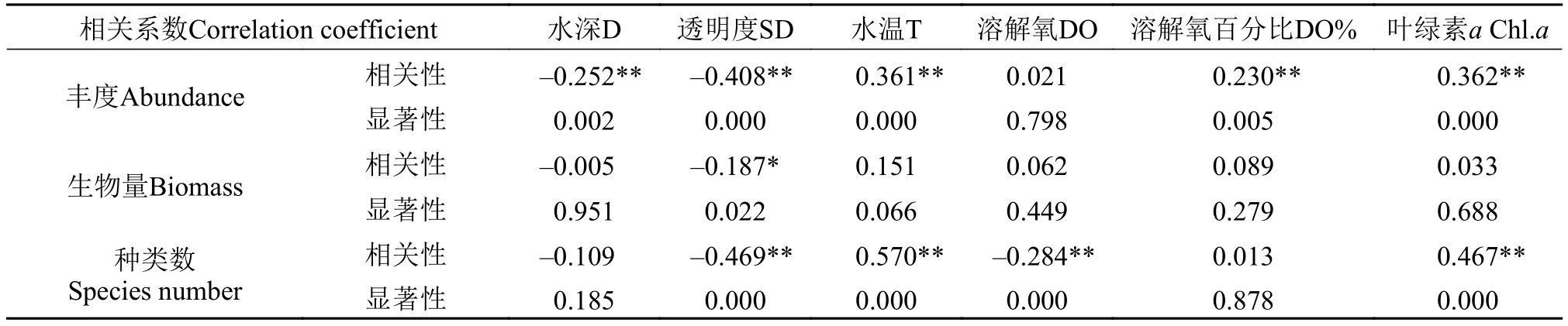
表 3 浮游动物与环境因了相关分析Tab. 3 The correlation analysis of zooplankton density, biomass and environmental factors
浮游动物与环境因子冗余分析 对太平湖浮游动物和各环境因子先进行降趋对应分析(DCA),结果显示最大的排序轴梯度长度小于4, 因此选择对物种数据与环境因子数据作RDA分析, 分析结果如表 4、表 5和图 11所示。
由表 4可见, 排序轴一和排序轴二的特征值分别为0.288和0.140, 分别解释了物种环境方差的61.7%和29.9%, 达总标准特征值的99.1%。RDA排序中的透明度(SD)、水深(D)、温度(T)、溶解氧(DO)和叶绿素a (Chl. a)等环境因子共解释了物种组成总变异的46.3%。轴一的物种环境相关系数为0.837, 轴二的物种环境相关系数为0.780, 表明太平湖环境因子与浮游动物群落组成关系密切。
由表 5可得, 物种两排序轴与环境因子两排序轴的相关系数分别为0, 表明排序结果是可信的。从表中也看出水温和叶绿素a是RDA第一物种排序轴的重要影响因子, 呈正相关相关系数分别为0.6661,与透明度呈负相关相关系数为–0.1356; 第二物种排序轴与水深、叶绿素a呈负相关性, 其相关系数分别为–0.1264和–0.1577, 而与透明度、溶解氧呈正相关性, 其相关系数分别为0.1306和0.149。从RDA运行结果显示, 轮虫在1轴上方, 对温度的影响较高, 而枝角类与桡足类在1轴下方, 对水温的影响低于轮虫, 但对叶绿素a影响高于轮虫, 原生动物类对透明度要求较高。(图 11)。
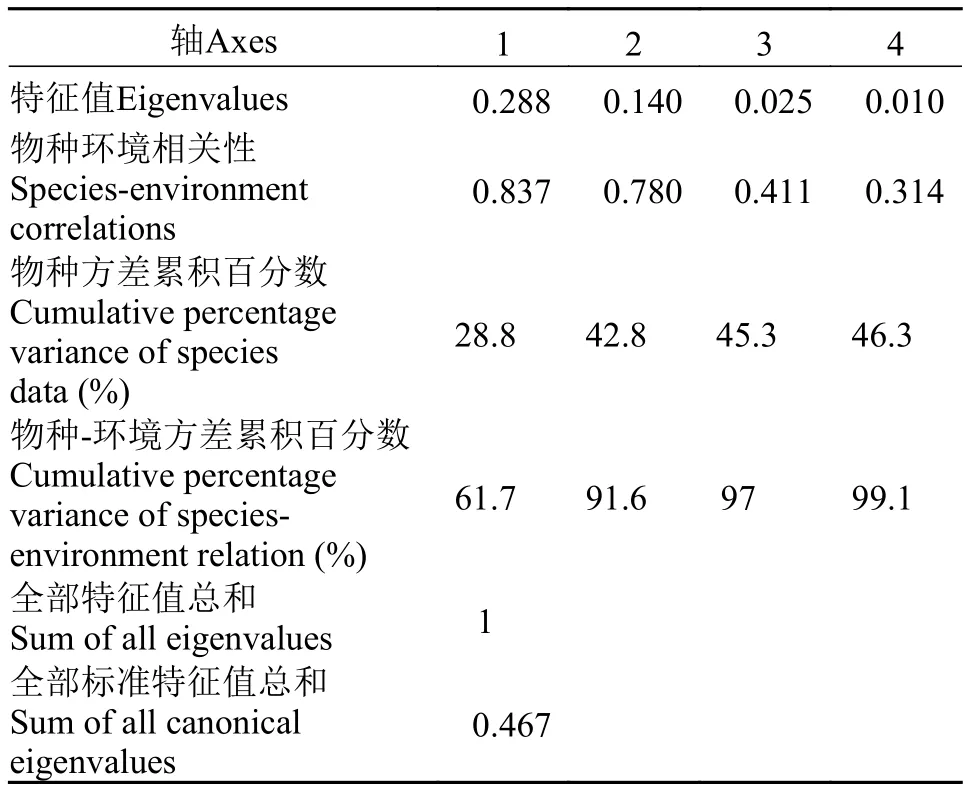
表 4 太平湖浮游动物种类和环境因子间RDA分析的统计信息Tab. 4 The axes of RDA performed on zooplankton in the Taiping Lake
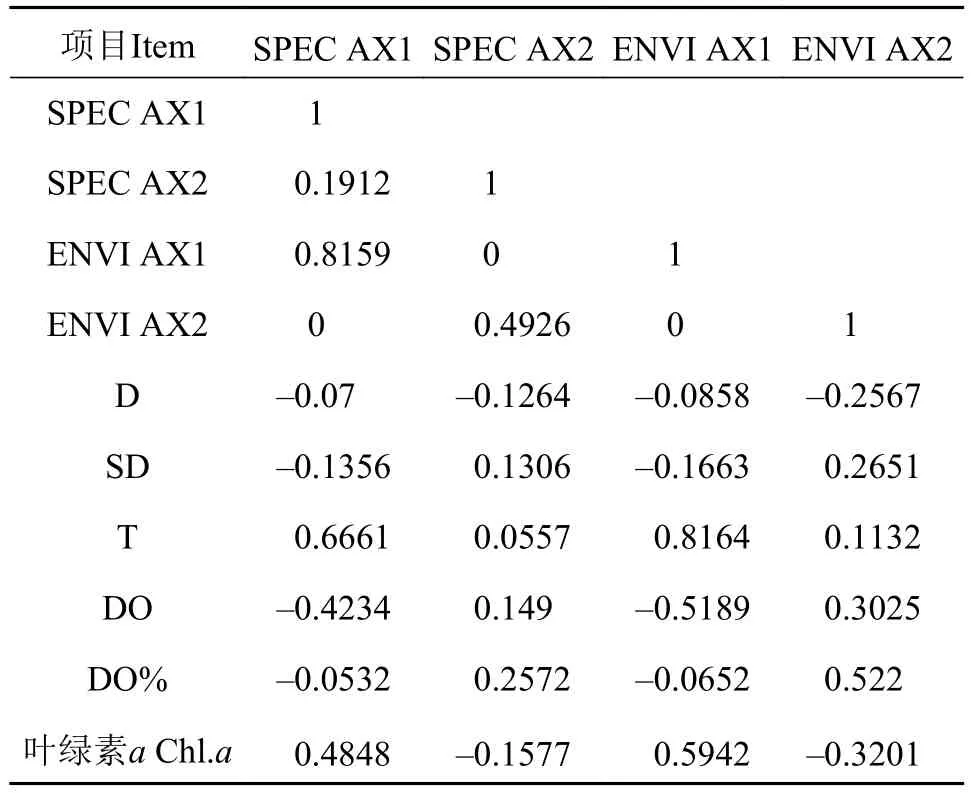
表 5 前2个排序轴和环境因子间的相关系数Tab. 5 Correlation coefficients of environmental factors with the first two axes of RDA
浮游动物与环境因子结构方程模型 结构方程模型(Structure Equation Modeling, SEM)融合了传统多变量统计分析中的“因素分析”与“线性模型回归分析”的统计技术, 对于各种因果模型可以进行模型辨识、估计与验证, 近年来在生态学研究中的应用也非常广泛[17]。为研究主要环境因子如何对浮游动物生物量和丰度的影响, 本文通过回归分析筛选出的影响因子(溶解氧、叶绿素a、透明度以及水温)进行模型的假设。通过对模型进行违规估计检验、拟合优劣指标检验和模型修订指标检验, 选出有显著作用的因子和路径, 结果如图12所示, 该模型的近似均方根误差(RMSEA=0.022<0.05)与卡方自由度之比(CMIN/DF=1.074<3)表明观测数据与模型呈较好的拟合。
从图 12可知浮游动物丰度受溶解氧和水温直接正向作用, 受透明度直接负向作用, 其中水温对浮游动物的作用较大, 而浮游动物丰度又对其生物量作用较大。透明度还可通过水温间接影响浮游动物丰度; 水温能通过对水体溶解氧的影响, 间接对浮游动物丰度和生物量起作用, 水温也能直接影响浮游动物生物量, 其中间接影响生物量作用比较大, 而直接影响丰度作用比较大, 叶绿素a主要通过间接影响浮游动物, 随着叶绿素a含量升高会导致水体透明度的下降从而影响水温和浮游动物丰度。
3 讨论
3.1 浮游动物群落结构特征及其影响因子
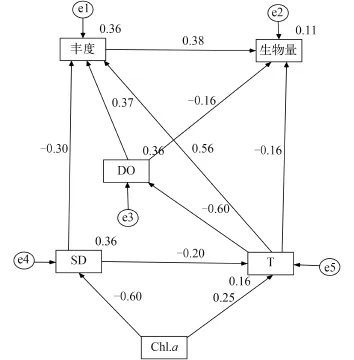
图 12 环境因子对浮游动物的直接或间接作用最终简化模型Fig. 12 The final structural equation model of direct and indirect effects of environmental factors on zooplankton
对浮游动物群落结构, 种类组成、丰度值和优势种的研究的报道很多, 如李喆等[18]在乌苏里江下游段发现了轮虫22种属、枝角类11种属和桡足类4种属, 浮游动物丰度的年均值达160 ind./L。在江西赣江, 计勇等[19]发现浮游动物群落由15属25种轮虫、6属7种枝角类和5属5种桡足类组成, 群落的优势种来自于臂尾轮属和腔轮属, 浮游动物的丰度值在丰水期为375 ind./L、枯水期为172 ind./L。而本文对太平湖进行为期两年的调查研究中, 发现浮游动物共检测到89种, 其中小型浮游动物轮虫种类所占比例最多, 与上述报道结果相同, 而轮虫种类存在较明显季节变化(图 5), 表现为夏秋季高于春冬季的格局, 这可能是由于浮游植物在夏秋季丰度高,且多为轮虫喜食的绿藻种类, 有利于轮虫的生长繁殖[20,21]; 原生动物种类次之, 季节变化表现为夏季最高, 秋季则未检测出, 这可能由于原生动物受多种环境因子的影响, 夏季光照、水温和营养盐等较秋季更高, 从而更适宜原生动物生长, 原生动物除受各种环境因子的影响, 亦受种间关系的相互作用,由于水体中各种生物存在相互依存、相互制约的复杂关系, 导致原生动物种类数发生变化; 四季中大型浮游动物枝角类与桡足类所占比例则均较少,有研究表明枝角类与轮虫存在较强的竞争关系, 当轮虫数量大量升高时, 会给枝角类的生长带来抑制作用[22]。浮游动物丰度季节变化规律与种类变化相同表现为夏季较高, 但其生物量则表现为春季较高, 这由于春季时个体较大的枝角类数量有所增加,当生物个体越大, 其在单位体积中所能达到的数量越小, 而所能达到的生物量则越大, 因而生物量春季较高。太平湖浮游动物丰度垂直分布表现为春夏季表层最高, 中间层次之, 底层最小, 而秋冬季则表现为中间层最高, 可能由于春夏季表层水温升高较快, 更易达到浮游动物最适温度, 而秋冬季温度逐渐下降, 但中间层水温变化不明显, 固春冬季中间层表现最高。整个调查期间太平湖轮虫种类在各季节中所占比例均较大这与大部分湖泊研究结果相同[23—25]。对太平湖浮游动物种类、丰度、多样性指数和各理化指标进行聚类分析(图 10), 结果表明太平湖浮游动物主要可分为两类群即夏秋季类群和春冬季类群, 夏秋季类群种类、丰度和多样性指数均较高, 而春冬季类群相对较低, 从整个湖区划分看, 两类群的浮游动物群落结构均存在湖心与上下游两个差异较显著的区域, 且春冬季这种差异更明显, 这可能与周边水文条件有关。
大量研究表明捕食和温度是影响浮游动物群落结构、丰度以及浮游动物群落演替最主要的生物与非生物因子[26—28]。滤食性鲢鳙鱼类对浮游动物的捕食所带来的下行效应和以浮游植物等为饲料的上行效应对浮游动物种类组成和生物量产生重要影响。本研究发现, 太平湖浮游动物主要以轮虫类小型个体为主, 这可能就是由于桡足类和枝角类等较大个体承受了较大的摄食压力所致, 依据体积效率假说, 鱼类对浮游动物的捕食具有明显的选择性, 通常在同等条件下将优先选择个体较大的种类为食[29]。另一方面, 温度对浮游动物群落结构具有显著影响, 每一种浮游动物都有其最适的温度范围, 超过这个范围就会影响浮游动物的生长, 原生动物生存的温度可以在0—35℃, 其最适温度为10—25℃。太平湖浮游动物丰度与种类数季节变化表现为夏秋季高于春冬季对其进行相关分析和逐步回归分析也显示出温度是其主要的影响因子,这与吴建新等[30]对田湾核电站海域浮游动物与温度的研究结果相同。通过RDA排序同样显示温度与第一排序轴有较好的相关性, 其相关系数为0.6661, 对太平湖浮游动物动态变化具有重要影响。从RDA排序图显示轮虫与温度具有较强的正相关性, 且在夏季温度较高时种类最多。绝大部分轮虫在15—25℃内, 其发育时间会随着温度的升高而缩短, 轮虫总数的高峰一般在水温20℃以上时出现[23], 进一步说明太平湖轮虫季节变化较明显。
除温度对浮游动物群落的影响外, 水体透明度和叶绿素a含量也影响了浮游动物群落的变化, 水体中浮游植物的升高可能引起水体透明度的下降,而浮游植物又是浮游动物的重要食物来源之一, 浮游植物数量的高低与浮游动物的生长繁殖存在密切相关, 而从某一方面叶绿素a含量能够反应浮游植物的变化, 温新利等[22]对芜湖市镜湖轮虫群落结构分析结果显示叶绿素a浓度与轮虫丰度具有显著的相关性, 林青等[25]对滴水湖研究也表明浮游动物密度与生物量也也与叶绿素a呈极显著正相关关系和回归关系, 而本研究结果也表明浮游动物丰度和种类数与叶绿素a呈极显著正相关, 从排序图中也可看出叶绿素a与透明度之间存在一定的负相关性,进一步讨论叶绿素a和透明度对浮游动物群落结构的影响, 对其建立结构方程模型(图 12)。通过结构方程模型得出叶绿素a主要通过影响水体透明度和水温间接作用于浮游动物, 这也进一步证明水体透明度和水温对浮游动物群落的影响。
3.2 太平湖水质评价
群落生物多样性是衡量群落稳定性的一个重要尺度, 其下降会破坏生态系统功能, 导致可更新资源的利用率下降, 而多样性指数则是衡量浮游动物群落健康状态和规模的尺度。一般而言, Margalef丰富度指数(d)大小与浮游动物种类数相关, 并主要反映物种种类的分布情况, 浮游动物种类数越多, 每种的个体丰度分布越均匀, 水体水质也就越好; Shannon-Wiener多样性指(H′)常用于反映群落的复杂程度, 物种多样性指数越大表明群落复杂程度越高水质越好; Pielou均匀度指数(J′)则表示浮游动物个体数目分配的均匀程度高低, 越高显明浮游动物群落结构越稳定[2]。结合Valentin和Heip等[31,32]的观点, 选择Shannon-Wiener多样性指数, 并结合Pielou均匀度指数和Margalef丰富度指数对太平湖水质进行综合性评价, 其中, Shannon-Wiener多样性指数: 0—1, 重度污染; 1—2, 中度污染; 2—3, 轻度污染; 大于3, 清洁; Pielou均匀度指数: 0—0.3, 重度污染; 0.3—0.5, 中度污染; 0.5—0.8, 轻度污染; 大于0.8, 清洁; Margalef丰富度指数: 0—1,重度污染; 1—2, 中度污染; 2—3轻度污染; 大于3,清洁[33]。这3个指数的数值越大, 水体的清洁度越高, 污染级别越低。
由Shannon-Wiener多样性指数和Margalef丰富度指数可以发现, 太平湖水质处于中度污染, 从Pielou均匀度指数指示水质处于轻度污染。从季节变化分析表现出夏秋季水质较春冬季好, 这可能由于在冬季和春季两个采样时间段内, 浮游动物的种类数较少, 且相对集中; 其次从初春开始, 太平湖进入涨水期, 这个过程会将上游的一些污染物、树枝,甚至垃圾等推进湖内, 从而引起了太平湖水质的变化, 改变浮游动物的生境, 影响浮游动物的生长和繁殖, 进而导致了浮游动物多样性指数的改变。因此, 在太平湖保护过程中, 应严格控制周边污染源与水生植被的种植及鱼类的合理投放与捕捞。
[1]Sun Z Q, Shi X L, Xu L L, et al. The protozoan community structure and its response to the change of water quality in atypical wetland landscape in summer [J]. Acta Hydrobiologica Sinica, 2013, 37(2): 290—299 [孙志强,施心路, 徐琳琳, 等. 景观湿地夏季原生动物群落结构与水质关系. 水生生物学报, 2013, 37(2): 290—299]
[2]Li H, Qi F, Xie C G, et al. Zooplankton in Bosten Lake: community characteristics and seasonal changes in distribution [J]. Journal of Fishery Sciences of China, 2013, 20(4): 832—842 [李红, 祁峰, 谢春刚, 等. 博斯腾湖浮游动物群落结构特征与分布的季节性变化. 中国水产科学, 2013, 20(4): 832—842]
[3]Jie B F, Zhang Z H, Cheng G P, et al. Analysis of plankton community structure and diversity in Guijiang River [J]. Guangdong Agricultural Sciences, 2014, 41(6): 171—177 [介百飞, 张紫皇, 程光平, 等. 桂江浮游生物的群落结构及多样性分析. 广东农业科学, 2014, 41(6): 171—177]
[4]Tang J, Xiang J G, Gu P H, et al. Community structure of zooplankton and assessment of water quality in Weishan Lake [J]. Chinese Journal of Fisheries, 2014, 27(2): 32—37 [唐俊, 向建国, 谷庞华, 等. 微山湖浮游动物群落结构及水质评价. 水产学杂志, 2014, 27(2): 32—37]
[5]Wang Z J, Du G L, Hong J M. Study advances on structure and diversity of zooplankton community [J]. Journal of Capital Normal University (Natural Science Edition), 2008, 29(3): 41—43, 50 [王正军, 杜桂森, 洪剑明. 浮游动物群落结构和多样性的研究. 首都师范大学学报(自然科学版), 2008, 29(3): 41—43, 50]
[6]Tao X M, Wang X Y, Wang L Q, et al. A Study of the community structure of metazoan zooplankton in Gehu Lake, Jiangsu Province, China [J]. Journal of Ecology and Rural Environment, 2013, 29(1): 81—86 [陶雪梅, 王先云, 王丽卿, 等. 滆湖后生浮游动物群落结构研究. 生态与农村环境学报, 2013, 29(1): 81—86]
[7]Carpenter K E, Johnson J M, Buchanan C. An index of biotic integrity based on the summer polyhaline zooplankton community of the Chesapeake Bay [J]. Marine Environmental Research, 2006, 62(3): 165—180
[8]Duan M, Zhu L, Feng J F, et al. Preliminary study on the ecological criteria calculation method based on the variation of plankton communities [J]. Research of Environmental Sciences, 2012, 25(2): 125—132 [段梦, 朱琳, 冯剑丰, 等. 基于浮游生物群落变化的生态学基准值计算方法初探. 环境科学研究, 2012, 25(2): 125—132]
[9]Kuang Q J, Xia Y C. Community dynamics of algae and limiting nutrient in Taipinghu reservoir [J]. Journal of Lake Sciences, 1995, 7(3): 235—239 [况琪军, 夏宜琤.太平湖水库藻类演替与营养侦别研究. 湖泊科学, 1995, 7(3): 235—239]
[10]Zhang S Y, Liu Q X, Hua L. Physicochemical characteristics of Taiping reservoir [J]. Acta Hydrobiologica Sinica, 2000, 24(5): 530—536 [张水元, 刘衢霞, 华俐. 太平湖水库水质的理化特征. 水生生物学报, 2000, 24(5): 530—536]
[11]Lu L, Tian N, Yu H, et al. The evolution process and mechanism of Taiping lake in Anhui province [J]. Journal of Natural Resources, 2015, 30(4): 604—616 [陆林,天娜, 虞虎, 等. 安徽太平湖旅游地演化过程及机制. 自然资源学报, 2015, 30(4): 604—616]
[12]Liu Y F, Qian L P, Huang H H, et al. Distribution patterns of heavy metals in surface sediments and environmental quality assessment on the Taipinghu basin of Anhui Province [J]. Journal of Fudan Univerisity (Natural Science), 2015, 54(2): 220—227 [刘亚风, 钱丽萍, 黄洪辉, 等. 安徽太平湖流域底质沉积物多种重金属和砷的分布特征及环境评价. 复旦学报(自然科学版), 2015, 54(2): 220—228]
[13]Wang J J. Freshwater Rotifer Fauna in China [M]. Beijing: Science Press. 1961, 288 [王家楫. 中国淡水轮虫志. 北京: 科学出版社. 1961, 288]
[14]Zhou F X, Chen J H. Atlas of Microbiology in Freshwater [M]. Beijing: Chemical Industry Press. 2005, 35—359 [周凤霞, 陈剑虹. 淡水微型生物与底栖动物图谱. 北京:化学工业出版社. 2005, 35—359]
[15]Koste W. Rotatoria: Die Rädertiere Mitteleuropas [M]. Berlin: Publ. Gebrüder Borntraeger. 1978, 673
[16]Du P, Xu X Q, Liu J J, et al. Spatial heterogeneity of macro-and meso-zooplankton in Xiangshan Bay in spring and summer [J]. Acta Ecologica Sinica, 2015, 35(7): 2308—2321 [杜萍, 徐晓群, 刘晶晶, 等. 象山港春、夏季大中型浮游动物空间异质性. 生态学报, 2015, 35(7): 2308—2321]
[17]Tao M, Wang Y M, Xie B W, et al. Spatio-temporal distribution of plankton and driving environmental factors in the Tuojiang River [J]. Acta Hydrobiologica Sinica, 2016, 40(2): 301—312 [陶敏, 王永明, 谢碧文, 等. 沱江浮游生物群落时空分布及相关环境因子分析. 水生生物学报, 2016, 40(2): 301—312]
[18]Li Z, Jiang Z F, Ma B, et al. Zooplankton community structure and current status in river health assessment for the lower branch of River Wusuli [J]. Chinese Journal of Fisheries, 2011, 24(3): 48—52, 59 [李喆, 姜作发, 马波,等. 乌苏里江下游浮游动物群落结构及在河流健康评价中的地位. 水产学杂志, 2011, 24(3): 48—52, 59]
[19]Ji Y, Zhang J, Sun X Q, et al. Spatio-temporal distribution characteristics of zooplankton and water quality assessment in middle and lower reaches of Ganjiang River [J]. Journal of Hohai University (Natural Sciences), 2012, 40(5): 509—513 [计勇, 张洁, 孙晓秋, 等. 赣江中下游浮游动物时空分布特征及水质综合评价. 河海大学学报(自然科学版), 2012, 40(5): 509—513]
[20]Nogueira M G. Zooplankton composition, dominance and abundance as indicators of environmental compartmentalization in Jurumirim Reservoir (Paranapanema River), SaoPaulo, Brazil [J]. Hydrobiologia, 2001, 455(1): 1—18
[21]Gilbert J J. Competition between Rotifers and Daphnia [J]. Ecology, 1985, 66(6): 1943—1950
[22]Wen X L, Xi Y L, Zhang L, et al. Analysis of community structure of rotifera and ecological assessment of water quality in Lake Jinghu, Wuhu city [J]. Acta Hydrobiologica Sinica, 2006, 30(2): 152—158 [温新利, 席贻龙, 张雷, 等. 芜湖市镜湖轮虫群落结构分析及水质的生态学评价. 水生生物学报, 2006, 30(2): 152—158]
[23]Yang L J, Lü G H, Zhu J Q, et al. Characteristics of zooplankton community in hengshan reservoir and water quality assessment [J]. Acta Hydrobiologica Sinica, 2014, 38(4): 720—728 [杨亮杰, 吕光汉, 竺俊全, 等. 横山水库浮游动物群落结构特征及水质评价. 水生生物学报, 2014, 38(4): 720—728]
[24]Zhang T, Ma X H, Wang G P, et al. Community structure and spatial distribution of plankton of plankton in the Poyang lake national nature reserve, China [J]. Acta Hydrobiologica Sinica, 2014, 38(1): 158—165 [张婷, 马行厚, 王桂苹, 等. 鄱阳湖国家级自然保护区浮游生物群落结构及空间分布. 水生生物学报, 2014, 38(1): 158—165]
[25]Lin Q, You W H, Xu F J, et al. Zooplankton community structure and its relationship with environmental factors in Dishui Lake [J]. Acta Ecologica Sinica, 2014, 34(23): 6918—6929 [林青, 由文辉, 徐凤洁, 等. 滴水湖浮游动物群落结构及其与环境因子的关系. 生态学报, 2014, 34(23): 6918—6929]
[26]Wang T. Predation and Competition: Interspecies Interaetions of Zooplankton Communities in LiuxiheReservoir [D]. Doctoral Dissertation, Jinan University, Guangzhou. 2010 [望甜, 捕食与竞争-流溪河水库浮游动物群落的种间关系研究, 博士学位论文, 暨南大学, 广州. 2010]
[27]Aleksandra M L, Hillebrand H, Lengfellner K, et al. Temperature effects on phytoplankton diversity-The zooplankton link [J]. Journal of Sea Research, 2014, 85(3): 359—364
[28]Dumont H J. Biogeography of rotifers [J]. Hydrobiologia, 1983, 104(1): 19—30
[29]Yang Y F, Huang X F, Liu J K, et al. Effects of fish stocking on the zooplankton community structure in a shallow lake in China [J]. Fisheries Management and Ecology, 2005, 12(2): 81—89
[30]Wu J X, Yan B L, Feng Z H, et al. Zooplankton ecology near the Tianwan Nuclear Power Station [J]. Acta Ecologica Sinica, 2011, 31(22): 6902—6911 [吴建新, 阎斌伦,冯志华, 等. 田湾核电站海域浮游动物生态特征. 生态学报, 2011, 31(22): 6902—6911]
[31]Valentin L, Macedo E, Tenenbanumdr, et al. The specific diversity indices for analyzing phytoplankton succession: application to the Caho Frio (RJ, Brazil) upwelling ecosystem [J]. Neritica, 1991, 6: 1—2
[32]Heip C, Engels P. Comparing species diversity and evenness indices [J]. Journal of the Marine Biological Association of the United Kingdom, 1974, 54(3): 559—563
[33]Wang X Q, Zeng Y Y, Wu H H, et al. Investigation on plankton community composition and water quality state in the mainstream of Xiangjiang River [J]. Acta Hydrobiologica Sinica, 2013, 37(3): 488—494 [王晓清, 曾亚英,吴含含, 等. 湘江干流浮游生物群落结构及水质状况分析. 水生生物学报, 2013, 37(3): 488—494]
THE RESEARCH OF DYNAMIC SUCCESSION OF ZOOPLANKTON AND ITS CORRELATION WITH ENVIRONMENTAL FACTORS IN TAIPING LAKE
FENG Shi-Min1, WU Ming-Shu1, XIONG Lian1, LI Dong-Jing1, WANG Jun-Li1and LIU Dong-Yan1,2
(1. Environmental Science from Shanghai Normal University, Shanghai 200234, China; 2. The Ecological and Environmental Research Center at Shanghai Normal University City, Shanghai 200234, China)
The community characteristics and distribution of zooplankton in Taiping Lake were investigated from November 2012 to October 2014. The study identified a total of 89 species of the zooplankton belonged to 45 genera, including 69 Rotifera belonged to 29 genera, 7 Cladocera belonged to 5 genera, 4 Copepoda belonged to 2 genera and 9 Protozoa belonged to 9 genera. The dominant species were the Rotifera (Trichocereca) and (Keratella). We found the variability of spatial and temporal distribution of zooplankton abundance in Taiping Lake with the maximum of 1326 ind. /L in summer and the minimum of 216 ind./L in winter while in spring and autumn they were 608 ind./L and 605 ind. /L respectively. The vertical distribution of zooplankton abundance in water body showed that the surface abundance had the highest concentration, followed by the middle layer and the bottom layer in spring and summer, while the middle layer had the highest abundance in autumn and winter. Both Shannon-Wiener diversity index and Margalef abundance index suggested that the middle layer was generally higher than the surface and bottom, while Pielou index indicated that the bottom layer had the highest one in all three layers. All the diversity indexes suggested significantly higher value in summer and fall than that in spring and winter, and the water quality in summer and fall was better than that in spring and winter. The cluster analysis and multi-variate analysis revealed that zooplankton can divide into spring-winter group and summer-fall group, and there were comparatively big differences in zooplankton community structure from the central area to upstream and downstream of Taiping Lake with more obvious differences in the spring-winter group. The correlation and stepwise regression analysis displayed a significant relationship between zooplankton community and environmental factors. These results indicated that the transparency and the water temperature were the main environmental impact factors in regulating zooplankton community structure in Taiping Lake. Structure Equation Modeling (SEM) and Redundancy Analysis (RDA) indicated that zooplankton abundance was higher in habitats with high temperature and dissolved oxygen, and that the temperature had bigger influence on Rotifera than on Cladocera and Copepoda.
Taiping Lake; Zooplankton; Correlation analysis; RDA analysis; Structure Equation Modeling (SEM)
S932.8
A
1000-3207(2017)03-0700-12
10.7541/2017.88
2016-05-09;
2016-07-10
国家自然科学基金(31170441; 31070419); 上海师范大学原创与前瞻性预研项目(DYL201304)资助 [Supported by the National Natural Science Foundation of China (31170441; 31070419); Original and Forward-looking Pre-research Project of Shanghai Normal University (DYL201304)]
冯世敏(1990—), 男, 江西上饶人; 硕士研究生; 主要从事水域生态研究。E-mail: uc_ok@126.com
刘冬燕, E-mail: liudy@shnu.edu.cn
