OsMT-1-4c基因原核表达载体的构建、表达及重金属抗性研究
2017-03-31王小兰
唐 伟,张 美,王小兰
(1.广州大学生命科学学院,广东 广州 510006;2.中国科学院华南植物园,广东 广州 510650)
OsMT-1-4c基因原核表达载体的构建、表达及重金属抗性研究
唐 伟1,张 美2,王小兰1
(1.广州大学生命科学学院,广东 广州 510006;2.中国科学院华南植物园,广东 广州 510650)
为研究水稻抗重金属的特性,构建了水稻金属硫蛋白(metallothionein,MT)原核表达载体PGEX4T-3-OsMT-1-4c,并转化到Rossetta菌株中诱导表达,得到大小为33.643 kDa的GST-OsMT-1-4c重组蛋白。将得到的沉淀用1×PBS溶解并在低温下超声破碎,离心并电泳检测,然后纯化得到融合蛋白。并在重金属Cd2+胁迫下诱导表达重组载体和空载体菌株,发现能表达GST-OsMT-1-4c蛋白菌株比表达GST蛋白菌株抗重金属的能力强。
水稻;金属硫蛋白;原核表达;纯化;抗重金属特性
重金属在环境中是普遍存在的,对植物而言,重金属分为必需元素和非必需元素,但是非必需重金属元素和过量的必需重金属元素对植物的生长和发育都是有害的,对植物毒害作用主要表现在损伤细胞膜[1]、破坏线粒体和叶绿体的超微结构[2]、减少叶绿素和抗坏血酸含量、产生活性氧分子、降低各种的酶活性[3],继而阻碍植物呼吸代谢、光合作用[4]和细胞分裂等生命活动的正常进行。植物生长在重金属污染的环境中[5],受重金属的胁迫,植物与重金属接触的界面首先受到影响,并且这种影响随着胁迫时间的延长,随重金属浓度的升高,植物的受害亦加剧。植物受重金属的伤害程度、伤害症状与重金属的种类也有密切关系。重金属能够通过农作物直接参与食物链循环[6],最终在人体内富集[7],使人体健康受到危害。水稻是我国种植面积最大的粮食作物,稻米的产量、品质及其安全问题一直是社会关注的焦点。当重金属进入水稻体内后,大部分会积累在水稻根部[8],很少一部分向地上组织迁移,积累在主茎稻穗、叶片以及籽实中[9]。水稻籽实不同部位中的重金属含量较不均匀,胚乳中总量最高[10]。据报道,我国华南地区稻米重金属含量符合标准的只有64.8%[11]。因此,研究水稻体内响应金属胁迫的关键基因,揭示其对重金属离子的吸收、转运和解毒方面的分子机制[12],对水稻的逆境育种、提高稻米的品质和解决食品安全问题有重大意义。
金属硫蛋白(metallothionein,MT)的研究是当今生物化学和分子生物学的热点之一,研究表明金属硫蛋白是一类富含半胱氨酸,能结合重金属的低分子量的蛋白。其主要功能是与重金属离子结合,一般通过半胱氨酸残基与金属离子结合成螯合物从而减轻或消减重金属的毒害作用[13]。此外MT 具有清除活性氧(reactive oxygen species,ROS)的功能在抗衰老、抗氧化压力及细胞凋亡等过程中发挥着重要作用,且MT是一种潜在的细胞凋亡负调控因子,在化疗中对正常机体细胞有一定的保护作用[14],但其作用机理仍不清楚。由于植物体内MT的含量很低、分子量极小、富含的半胱氨酸残基极易被氧化,导致对植物MT的研究主要停留在转录水平上,而对其功能的研究相对较少。有研究报道MT具有耐受重金属Cd的特性[15],MT的大量表达能显著提高植物耐受性。当今对水稻金属硫蛋白基因家族的研究较多,但是对OsMT-1-4c基因研究相对较少,现有的文献还没有纯化得到其蛋白,也没有对其重金属特性做初步研究,因此本实验室从水稻cDNA酵母表达文库[16]中分离得到了OsMT-1-4c基因(http://rice.plantbiology.msu.edu/, LOC_ Os01g10400),并将其构建到PYES260载体上保存,其基因大小为237 bp,编码78个氨基酸,其半胱氨酸含量达到16.7 %,从而推测其具有较强的抗重金属能力。本研究构建了水稻金属硫蛋白重组表达质粒PGEX4T-3-OsMT-1-4c,转入Rossetta菌株中诱导表达重组蛋白,优化诱导条件后,产生大量可溶的谷胱甘肽巯基转移酶融合蛋白(GST-OsMT-1-4c),并纯化得到了该重组蛋白。通过在重金属胁迫下产生重组蛋白的菌株与GST的菌株其生长曲线相比较,发现产生重组蛋白的菌株比GST蛋白菌株生长状况好很多,表明OsMT-1-4c蛋白在Rossetta菌株中的表达提高了其抗重金属的能力,从而初步验证了OsMT-1-4c蛋白具有抗重金属能力。
1 材料与方法
1.1 试验材料
原核表达载体PGEX4T-3、重组质粒PYES260和大肠杆菌DH5α由本实验室保存;限制性内切酶BamHⅠ、XhoI购自TaKaRa公司; IPTG购自艾基生物技术有限公司、胶回收试剂盒和质粒提取试剂盒购自Magen公司、GST Bind Resin和填料柱购自Novagen公司、引物合成及序列测定由上海英骏生物技术有限公司提供。
LB培养基配方:胰蛋白胨(Tryptone)10 g/L,酵母提取物(Yeast extract)5 g/L,氯化钠(NaCl)10 g/L,用2.5 mol/LNaOH调节培养基的pH,使其达到7.4,去离子水定容至1 L,即配成液体LB培养基。固体培养基100 mL添加1.5 g琼脂粉,然后120℃、20 min灭菌,待LB培养基不烫手后加入氨苄青霉素,至终浓度100 μg/mL,倒平板,凝固后放入4℃ 冰箱保存备用。
2×YT培养基配方:胰蛋白胨(Tryptone)10 g/L,酵母提取物(Yeast extract)5 g/L,氯化钠(NaCl)10 g/L,用2.5 mol/L NaOH调节培养基的pH,使其达到7.2,去离子水定容至1 L,即配成液体2×YT培养基。固体培养基100 mL添加1.5 g琼脂粉,然后120℃ 、20 min灭菌,冷却后室温保存备用。
SDS-PAGE所用试剂:30%丙烯酰胺混合液(Acr∶Bis为29∶1):将60 mL水预热至37 ℃,加入丙烯酰胺(Ars)29 g及甲叉丙烯酰胺(Bis)1.0 g,溶剂定容至100 mL,并用0.45微米孔径滤纸过滤,装入棕色瓶中4℃ 保存;1.5 mol/L Tris-HCl缓冲液:取1 mol/L HCl溶液48 mL、三羟基甲烷(Tris)36.6 g,加双蒸至90 mL,调pH至8.8,然后定容至100 mL,至于玻璃瓶中保存,室温贮存; 5×电泳缓冲液:取15.1 g Tris 和94 g甘氨酸溶于800 mL去离子水中,再加入5 g SDS,调节pH至8.3,定容至1 L;10%过硫酸铵(AP):过硫酸铵0.2 g于2.0 mL EP管中,定容至2 mL,现用现配;10%SDS(十二烷基磺酸钠):取10 g SDS加去离子水80 mL,68℃水浴加热溶解定容至100 mL,室温保存。
GST-OsMT-1-4c纯化所需试剂:结合液:Na2HPO4·12H2O 2.9 g,KH2PO40.2 g,NaCl 8.0 g,KCl 0.2 g,加 0.8 L 去离子水溶解,调pH 7.3,去离子水定容至 1 L,4℃保存;洗脱液:Tris 碱0.605 g,还原型谷胱甘肽(reduced glutathione)0.307 g,加 80 mL去离子水溶解,调 pH 8.0,去离子水定容 100 mL,-20℃保存。
1.2 重组质粒PGEX4T-3-OsMT-1-4c的构建
将含有OsMT-1-4c基因序列的质粒PYES260作为PCR的模板,通过网站 Convert Primers Into In-Fusion® Primers(http://bioinfo. clontech.com/infusion/convertPcrPrimersInit.do)设计引物如下:
OsMT-1-4c-PEF:CTGGTTCCGCGTGGATCCAT GTCTTGCTGCGGAGGAAG
OsMT-1-4c-PER:ACGATGCGGCCGCTCGAGTT AGCAGTTGCAGGGATTGC
PCR反应体系(50μL):10×Buffer 5.0μL,dNTP4.0μL,Taq DNA聚合酶0.5 μL,模板1μL,引物4μL,H2O 35.5μL。胶回收纯化的目的片段连接到PGEX4T-3的载体(已由NcoI和BamHI双酶切)上,转入大肠杆菌DH5α,菌落PCR鉴定,并将鉴定正确的PGEX4T-3-OsMT-1-4c保存菌种。
1.3 GST-OsMT-1-4c诱导表达
将测序正确的PGEX4T-3-OsMT-1-4c质粒转化到Rossetta感受态细胞,挑取单菌落进行PCR鉴定阳性克隆。阳性单克隆接种于2 mL含100μg/mL 氨苄青霉素 (Amp)、50μg/mL卡那霉素(Kam)、34μg/mL氯霉素(Chl)的 LB培养基上,37℃ 200 r/min过夜培养,按体积比1∶100转接到100 mL LB(含AmpKanChl)新鲜2×YT培养基,相同条件下培养,当OD600值到0.4~0.6,加异丙基硫代半乳糖苷(IPTG)至终浓度0.2 mmol/L,诱导表达,收集菌液离心,并用SDSPAGE凝胶电泳分析诱导表达结果。
1.4 GST-OsMT-1-4c蛋白纯化
通过SDS-PAGE电泳检测融合蛋白已经大量表达,收集菌体,沉淀用1×PBS悬浮,超声破粹30 min(9 s开,9 s关)离心50 min(4℃,8 000 r/min),收集上清液和沉淀,SDSPAGE检测融合蛋白在上清液还是沉淀中,如果在上清液中,则利用有GST标签的层析柱进行蛋白纯化。
1.5 重金属抗性分析
1.5.1 适合的金属离子浓度确定 通过分光光度计测定重金属胁迫下重组大肠杆菌的生长状况,并将PGEX4T-3的空载体作为对照,比较表达OsMT蛋白的大肠杆菌Rossetta耐受重金属离子镉的能力[17]。前期实验确定了Rossetta菌对Cd2+耐受范围,因此对Cd2+通过0、0.8、1.0、1.2 mmol/L浓度梯度试验,重复3次,确定金属离子合适的浓度。诱导方法如前所述,将加入IPTG和重金属离子定为0 h,每隔1 h测定1次OD600值,以OD600为纵轴、时间为横轴作图,确定对细菌生长和增殖有一定抑制作用,且未使细菌全部死亡的浓度范围。
1.5.2 表达GST-OsMT-1-4c与GST蛋白的Rossetta菌对重金属抗性比较 用吸光度法研究表达OsMT的大肠杆菌Rossetta细胞对金属离子的耐受性,并将空载体PGEXT4T-3菌株作对照,重复3次。在Rossetta菌液中加入IPTG和合适的金属离子浓度诱导,继续培养并每隔1 h测定一次OD600值,以OD600为纵轴作图。
2 结果与分析
2.1 PGEX4T-3-OsMT-1-4c载体的构建
根据引物所在位置,PCR的大小应为 237 bp,电泳检测结果显示在200~400 bp位置有条带(图1),说明OsMT-1-4c基因已经克隆成功,然后In-fusion连接并转化DH5α菌株,37℃培养,挑取单菌落进行菌落PCR鉴定,结果显示在200~400 bp位置有条带(图2)。再对该单菌落提取质粒测序,确定OsMT已经正确插入载体PGEX4T-3中,表明已经成功构建了PGEX4T-3-OsMT-1-4c的表达载体。

图1 PCR扩增 OsMT cDNA电泳结果

图2 PGEX4T-3-OsMT-1-4c菌液 PCR电泳结果
2.2 GST-0sM-1-4c蛋白诱导表达
将转入鉴定正确的载体菌株诱导表达,设收集菌体,SDS-PAGE凝胶电泳结果显示在37℃、3 h和0.2 mmol/L IPTG时蛋白诱导表达最大(图3)。将诱导表达最大的条件大量诱导,收集菌体,超声破碎收集上清和沉淀,发现融合蛋白基本在上清液中,将上清液低温保存备用。
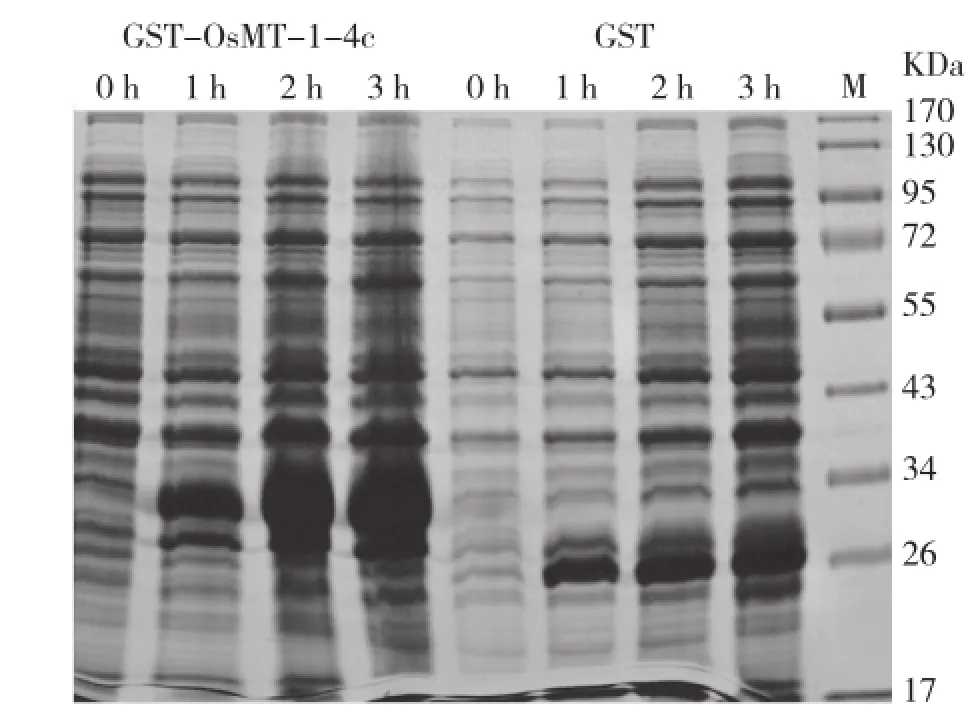
图3 GST-OsMT-1-4C和GST蛋白诱导表达
2.3 GST-0sM-1-4c蛋白纯化
将大量表达蛋白的上清液进行GST亲和层析柱纯化,得到蛋白洗脱液,再取少量洗脱液进行SDS-PAGE电泳分析,在26~34 kDa位置只有一条很粗的条带,GST大小为26 kDa,OsMT-1-4c蛋白大小为8 kDa,表明得到了纯化的融合蛋白GST-0sM-1-4c(图4)。

图4 GST-OsMT-1-4c蛋白纯化
2.4 重金属抗性分析
2.4.1 适合的金属离子浓度确定 选用Cd2+作为研究的金属离子[18],方法如前所述,每小时测定一次OD600值,重复3次,以时间为横轴作图(图5),确定对Rossetta菌的生长和增殖有一定抑制作用,又未使细菌全部死亡的浓度范围。通过比较4个浓度梯度细菌的生长状况发现,1.0 mmol/L浓度梯度下细菌的生长曲线斜率接近1,即在此浓度下对Rossetta菌的生长和增殖有一定抑制作用,又不至于死亡。从而确定实验合适浓度是1.0 mmol/L CdCl2。为了确定重组菌与对照菌(转入空载体PGEX4T-3的Rossetta菌)之间,在没有任何处理的情况下其生长状况是否一致,按上述方法培养大肠杆菌,结果显示当培养液不添加金属离子时,对照菌与重组菌生长状况基本相同,为进一步的实验排除干扰。

图5 Cd2+对含GST基因的Rossetta菌生长状况的影响
2.4.2 表达GST-OsMT-1-4c 与GST蛋白的Rossetta菌对重金属抗性比较 如图6所示,1.0 mmol/L CdCl2处理3 h内,对照菌和重组菌增长没有明显差异;处理3 h后,表达OsMT蛋白的重组菌增长率明显高于对照菌。说明表达OsMT融合蛋白确实增强了Rossetta菌对Cd离子的耐受性,提高了细菌的存活率,从而使相应的重组菌表现出耐受重金属离子的能力。

图6 Cd2+对表达GST和GST-MT-1-4c蛋白的Rossetta菌生长状况的影响
3 结论与讨论
金属硫蛋白是一类富含半胱氨酸残基的低分子量蛋白,能通过半胱氨酸残基上的巯基基团与金属离子结合,从而减轻重金属离子对植物体的危害。目前,对植物MT蛋白的研究很多,如在蚕豆中对MT基因的表达分析认为[19],MT能与金属离子直接结合[20],稳定植物内环境中金属离子的含量,并能有效的转运金属离子到有需要的组织,从而说明了植物MT基因参与了金属离子的运输的功能。在拟南芥中利用转MT-Ⅱ基因的超表达植株,分析表明,转基因拟南芥能有效维持体内氧自由基的含量,减少MDA的产生,从而缓解干旱胁迫引起的伤害,提高抗旱性[21]等。金属硫蛋白基因丰富的巯基含量及重金属结合的能力决定了功能的多样性。目前虽然对抗重金属方面研究不断深入,但是对其他功能尚不清楚,加上已知的植物MT蛋白较少,进一步阻碍了对MT功能的研究。因此,水稻金属硫蛋白表达载体的构建及抗重金属特性初步研究,将有助于进一步研究植物MT的功能。
由于金属硫蛋白在植物体内含量较少,分子量极低,且半胱氨酸极易被氧化,因此很难检测其在生物体内的表达活性。本研究的OsMT-1-4c蛋白仅为7.643 kDa左右,在大肠杆菌中表达很难直接检测。因此,本研究选用含有GST标签的蛋白表达载体PGEX4T-3,其GST大小为26 kDa,是根据唐青蓝等[22]用PGEX4T-3成功构建了原核表达载体,并与王娟等[23]选择PGEX系列构建海州香薷金属硫蛋白重组表达质粒PGEX-2T-Eh MT1相类似,为大量表达和纯化重组蛋白GST-OsMT-1-4c提供依据,为在蛋白水平上研究MT蛋白打下坚实的基础。表达载体的宿主菌为 Rossetta,是一种广泛使用的表达宿主菌,适合于 GST 表达体系,OsMT-1-4c蛋白在此宿主菌中活性稳定且产量高[24]。在IPTG诱导3 h后,目的蛋白得到大量表达,然后纯化得到GST-OsMT-1-4c重组蛋白。重金属抗性分析是通过重金属Cd2+胁迫含目的基因的菌株,通过目的蛋白的表达,比较表达GSTOsMT-1-4c蛋白与GST蛋白的菌株的生长状况,结果表明1.0 mmol/L CdCl2处理3 h内,对照菌和重组菌增长没有明显差异;处理3 h后,表达OsMT蛋白的重组菌增长率明显高于对照菌。说明OsMT-1-4c可能与金属离子相结合,从而使相应的重组菌表现出耐受金属离子的能力。通过本实验OsMT-1-4c表达,将为提高对重金属离子的耐受和毒性提供一个可行的基因策略,为今后在利用生物体抗重金属修复上提供应用前景[25]。
[1] Danielle L,Leonardo T. Lead poisoning:Successes and 21stcentury challenges[J]. Pediatrive in Review,2005,26(12):435-443.
[2] Stephan C. Toxic metal accumulation,responses to exposure and mechanisms of tolerance in plants[J]. Biochimie,2006,88(11):1707-1719.
[3] 梁琪惠,吴永胜,刘刚,等. Cr、As、Pb、Cd 复合污染对茶树叶片酶活性和细胞膜透性的影响[J]. 南方农业,2012(7):1-6.
[4] 梁文斌,薛生国,沈吉红,等. 锰胁迫对垂序商陆叶片形态结构及叶绿体超微结构的影响[J]. 生态学报,2011,31(13):3677-3683.
[5] 刘候俊,韩晓日,李军,等. 土壤重金属污染现状与修复[J]. 环境保护与循环经济,2012(7):4-8.
[6] 徐小逊,张世熔,解姗姗,等. 垂序商陆对 Cd 胁迫的生长响应和富集特征研究[J]. 西南农业学报,2012,25(4):1358-1362.
[7] 王义,黄先飞,胡继伟,等. 重金属污染与修复研究进展[J]. 河南农业科学2012,41(4):1-6.
[8] 孔令芳,朱红琼,杨晓霞. 重金属铅和汞对蚕豆根尖细胞的微核效应[J]. 大理学院学报,2011,10(4):51-53.
[9] 李荣春. Pb、Cd及其复合污染对烤烟叶片生理生化及细胞亚显微结构的影响[J]. 植物生态学报,2000,22(4):238- 242.
[10] 解凯彬,施国新,陈国祥,等. Hg2+对萍蓬草光合膜超微结构及功能的影响[J]. 农村生态环境,2002,18(1):26- 30.
[11] Bianka S,Margret S. Epidermal cell death in rice is confined to cells with a distinct molecular identity and is mediated by ethylene and H2O2through an autoamplified signal pathway[J]. Plant Cell,2009,21(1):184-196.
[12] 陈春,周启星. 金属硫蛋白作为重金属污染生物标志物的研究进展[J]. 农业环境科学学报,2009,28(3):425-432.
[13] Gupreet K,Harminder P S,Daizy R B,et al. Lead (Pb)-induced biochemical and ultrastructural changes in wheat(Triticum aestivum)roots[J]. Protoplasma,2013,250(1):53-62.
[14] 杨梅,程安春,汪铭书,等. 重组鸭α-干扰素基因工程菌表达条件的研究[J]. 四川农业大学学报,2007,25(3):337-342.
[15] 王晶,高燕,李静,等. 水稻cDNA酵母表达文库的构建及重金属镉胁迫相关基因的初步筛选[J]. 广东农业科学,2012,39(13):143-145.
[16] 张红晓,王世华,贺春玲,等. 水稻金属硫蛋白ricMT基因的耐受性分析[J]. 广东农业科学,2011,38(24):117-119.
[17] Li Y J,Wu H,Wei X X,et al. Subcellular distribution of Cd and Zn and MT mRNAexpression in the hepatopancreas of Sinopotamon henanense after single and co-exposure to Cd and Zn[J]. Comparative Biochemistry and Physiology(Part C):Toxicology & Pharmacology,2015,167:117-130.
[18] Rhonda C F,Liang Z M,Karam B S. Analysis of type 1 metallothionein c DNAs in Vicia faba[J]. Plant Mol Biol,1997,33(4):583-591.
[19] Kayo Y,Hitoshi N,Rie T,et al. Effects of soy protein isolate on LEC rats,a model of Wilson disease:mechanisms underlying enhancement of liver cell damage[J]. Biochemical and Biophysical Research Communications,2003,303:271-274.
[20] 唐中华,郭晓瑞,张洋洋,等. 拟南芥 MT-II 过量表达提高抗旱性[J]. 植物研究,2005,25(4):415-418.
[21] 唐青蓝,许庆忠,张礼林,等. pGE-4T-3-MFGF21表达载体的构建、表达与纯化[J]. 生物技术,2014,24(2):1-24.
[22] 王娟,王桂萍,张红生,等. 海州香薷金属硫蛋白(Eh MT1)的原核表达和纯化及其多克隆抗体的制备[J]. 南京农业大学学报,2010,33(4):25-30.
[23] Carstens C P,Waeshe A. Codon bias-adjusted BL21 derivatives for protein expression[J]. Strategies,1999,12:49 -51.
[24] 何永吉,刘进平,李颖君,等. 华溪蟹功能性重组金属硫蛋白的表达及抗血清制备[J]. 环境科学学报,2014,34(9):2434-2439.
(责任编辑 杨贤智)
Vector construction, prokaryotic expression and resistance to heavy metals of OsMT-1-4c
TANG Wei1,ZHANG Mei2,WANG Xiao-lan1
(1.College of Life Science, Guangzhou University,Guangzhou 510006,China;
2.South China Botanical Garden, Chinese Academy of Sciences,Guangzhou 510650,China)
In order to study the resistance of paddy rice to heavy metals, PGEX4T-3-OsMT-1-4c, the prokaryotic expression vector of metallothione for paddy rice was constructed, which was then transformed into strain Rossetta to induce the expression, about 33.643 kDa of GST-OsMT-1-4c fusion protein was obtained. The obtained precipitation was dissolved with 1×PBS and conducted ultrasonic fragmentation at low temperature,followed by the centrifugation and electrophoresis detection, and then was gained through the purification. Furthermore, when the recombinant vector and empty vector strains were induced to express under the stress of heavy metal Cd2+, it was found that the strain to express protein GST-OsMT-1-4c owned stronger resistance to heavy metals than that of protein GST.
rice;metallothionein; prokaryotic expression; purification;resistance to heavy metals
X592
A
1004-874X(2017)01-0008-07
2016-11-01
国家自然科学基金(30971912)
唐伟(1989-),男,在读硕士生,E-mail:907264698@qq.com
王小兰(1973-),女,博士,教授,E-mail:1372848641@qq.com
唐伟,张美,王小兰. OsMT-1-4c基因原核表达载体的构建、表达及重金属抗性研究[J].广东农业科学,2017,44(1):8-14.
