诱集植物香根草对二化螟幼虫致死的作用机制
2017-02-16鲁艳辉高广春郑许松吕仲贤
鲁艳辉,高广春,郑许松,吕仲贤
(1浙江省农业科学院植物保护与微生物研究所浙江省植物有害生物防控省部共建国家重点实验室培育基地,杭州 310021;2嘉兴学院医学院药学系,浙江嘉兴 314001)
诱集植物香根草对二化螟幼虫致死的作用机制
鲁艳辉1,高广春2,郑许松1,吕仲贤1
(1浙江省农业科学院植物保护与微生物研究所浙江省植物有害生物防控省部共建国家重点实验室培育基地,杭州 310021;2嘉兴学院医学院药学系,浙江嘉兴 314001)
【目的】香根草(Vetiveria zizanioides)能够有效诱集水稻害虫二化螟(Chilo suppressalis)雌成虫在其上产卵,但孵化出的幼虫在香根草上不能完成生活史。本研究旨在明确香根草对二化螟幼虫致死的作用机制,为开发以香根草为基础的水稻螟虫可持续控制新技术提供理论依据。【方法】通过有机溶剂萃取和硅胶柱层析方法提取香根草有毒活性物质,并利用饲料混毒法测定各提取组分对二化螟3龄幼虫的生物活性,同时利用生物化学方法测定水稻和香根草中营养物质含量以及二化螟3龄幼虫取食水稻和香根草后体内相关解毒酶和消化酶的活性变化。【结果】香根草石油醚萃取物处理二化螟3龄幼虫9、12、15 d后,死亡率分别为38.89%、57.41%、85.19%,显著高于乙酸乙酯萃取物、正丁醇萃取物等其他提取组分处理二化螟幼虫后的死亡率。进一步从香根草石油醚萃取物中分离获得组分1(Fr1)和组分5(Fr5),对二化螟3龄幼虫的具有较高毒性,以0.05 g·mL-1浓度混入人工饲料饲喂3 d后死亡率分别为85.00%和67.67%,说明香根草中含有对二化螟幼虫具有致死作用的有毒活性物质。香根草中总蛋白、纤维素、总糖、氨基酸等营养物质含量分别为9.45 μg、1.61%、1.45%、4.00%,均显著低于水稻中相应营养物质含量,其中水稻中蛋氨酸含量是香根草中的7.0倍。而香根草中单宁含量则显著高于水稻中的含量,是水稻中的1.31倍。此外,二化螟取食水稻和香根草3 d后,取食香根草的3龄幼虫体内蛋白酶、淀粉酶、海藻糖酶和蔗糖酶等消化酶的活性显著低于取食水稻幼虫体内相应消化酶的活性;6 d后,取食香根草的3龄幼虫体内细胞色素P450酶活性显著低于取食水稻的幼虫;9 d后,取食香根草的3龄幼虫体内羧酸酯酶CarE的活性显著低于取食水稻的幼虫。与取食水稻的3龄幼虫相比,取食香根草的幼虫体内谷胱甘肽硫转移酶GSTs的活性也有所降低,但是两者在统计学上没有显著差异。【结论】香根草作为二化螟的致死型诱集植物,对二化螟幼虫的致死作用主要表现在两方面:一是香根草中含有对二化螟幼虫具有致死作用的有毒活性物质,这些物质通过抑制幼虫体内解毒酶CarE和P450酶的活性,使幼虫逐渐丧失解毒代谢能力,最终死亡;二是香根草相对水稻营养物质匮乏,二化螟幼虫取食香根草后营养不均衡,从而影响体内消化酶活性,造成消化功能紊乱,影响其正常生理活动,最终死亡。
香根草;二化螟;致死作用;营养物质;解毒酶;消化酶
0 引言
【研究意义】二化螟(Chilo suppressalis)是危害水稻最重要的害虫之一,广泛分布于亚欧大陆。二化螟在苗期危害水稻可造成枯鞘、枯心和虫伤株,在孕穗期和穗期危害可造成枯孕穗和白穗等,严重影响水稻的质量和产量[1-3]。近年来,种植结构和耕作制度调整,杂交稻面积逐年增加[4],使越冬代有效虫源面积扩大,更加剧了二化螟的危害[5-6]。目前,多采用化学药剂对二化螟进行防治[7]。但是,化学农药的不合理使用,导致二化螟抗药性不断提高,加大了二化螟的猖獗危害。同时,化学农药带来的农药残留超标、环境污染等问题也日益突出。这些不利因素促使人们迫切寻求环境友好的作物保护方式[8]。【前人研究进展】香根草(Vetiveria zizanioides)又名岩兰草,是一种多年生草本植物,常用于生态环境治理、水土保持等方面。已有研究报道,香根草可以作为玉米田诱集植物引诱玉米禾螟(Chilo partellus),从而降低其对玉米的危害[9]。室内盆栽试验表明,香根草具有引诱稻螟雌成虫产卵的特性,二化螟在香根草上的产卵量是水稻上的4倍左右[10-11]。田间防控效果调查发现,种植香根草的小区稻螟造成的枯心率比对照区降低了50%以上[10]。此外,田埂种植香根草除了可诱集二化螟成虫在其上产卵外,且孵化后的幼虫在香根草上不能完成生活史,仅有极少数的幼虫能存活至2龄、3龄,从而显著降低了二化螟种群数量,减少了其对水稻的危害[11]。因此,香根草作为诱集植物防治稻螟已成为水稻害虫绿色防控的重要措施之一,具有很大的开发潜力[12-14]。诱集植物根据其不同特征可分为传统型诱集植物、致死型诱集植物和基因工程诱集植物[15]。致死型诱集植物是指该诱集植物对害虫极具吸引力,害虫一旦取食或者在其上产卵,害虫及其后代将无法生存[16]。【本研究切入点】对二化螟而言,香根草属于典型的致死型诱集植物,但香根草对二化螟幼虫的致死作用机制尚未见报道。为更好地建立以香根草为基础的水稻螟虫绿色防控新技术,明确其对二化螟幼虫的致死作用机制尤为重要[8]。【拟解决的关键问题】通过测定香根草提取物组分对二化螟幼虫的生物活性,比较香根草和水稻中的营养物质含量及取食水稻和香根草后二化螟幼虫体内解毒酶和消化酶活性的变化,从毒理学和营养学两方面揭示香根草对二化螟幼虫致死的作用机制,为进一步深入了解其致死作用机制及建立稻螟绿色防控新技术提供重要的理论依据,为化学农药的减施、水稻害虫的可持续治理及水稻的安全生产提供有力保障。
1 材料与方法
试验于2015—2016年在浙江省农业科学院完成。
1.1 供试植物
水稻品种浙优 12号购自浙江省农科种业有限公司。播种于温室塑料槽内,约30 d后移栽至直径为12 cm的塑料盆中,至45 d分蘖期时剪取茎秆以供营养物质测定及二化螟3龄幼虫的饲喂试验。
香根草种苗购自深圳市鑫淼森园林草坪有限公司。将香根草种苗种于温室,以分蘖苗作为繁殖体,待香根草分蘖较多时剪取茎秆供营养物质测定及二化螟3龄幼虫的饲喂试验。
1.2 供试昆虫
二化螟种群于2015年5月采自杭州市萧山区义桥镇(30°04′ N,120°12′ E)水稻田,室内建立并维持种群,幼虫采用人工饲料饲养,饲料配方由中国水稻研究所提供[17],置于温度(27±2)℃,相对湿度(70±5)%,光周期16L﹕8D的智能人工气候室内饲养。待二化螟化蛹后将蛹收集起来,放置在干净的养虫笼中,保湿,让其继续发育羽化,对羽化后的成虫以10%的蜂蜜水提供营养。
选取第1代二化螟3龄幼虫用于香根草活性物质的生物测定及水稻和香根草的饲喂试验。
1.3 香根草活性物质的制备及其对二化螟幼虫的生物活性测定
香根草活性物质的制备:称取0.5 kg香根草茎秆,阴干后用剪刀剪为长约2 cm的小节,置于多功能提取浓缩罐中,加80%的工业乙醇12 L室温浸泡20 h后,80℃恒温提取8 h,浓缩后得到乙醇提取物。将残渣置于中药煎药机,加蒸馏水6 L煎煮1.5 h,得水提取液。合并提取液,过滤浓缩蒸干得浸膏。试验重复两次,合并得总提取物36.6 g。取24.4 g浸膏制成1 L混悬液,依次用等体积石油醚、乙酸乙酯、正丁醇萃取 3次,分别浓缩得到石油醚萃取物6.0 g,乙酸乙酯萃取物4.3 g,正丁醇萃取物5.0 g和萃取后母液蒸干得到的水部位8.6 g,分别用于测定其对二化螟3龄幼虫的生物活性。
石油醚萃取物的硅胶柱层析:按上述方法称取1 kg香根草茎秆制备石油醚萃取物8.0 g,利用硅胶(100—200目)柱层析,分别以石油醚、石油醚:三氯甲烷(2﹕1)、石油醚:三氯甲烷(1﹕4)、三氯甲烷、乙酸乙酯各3 L梯度洗脱,分别收集流份减压浓缩干燥,获得5部分组合物,编号为Fr1—Fr5,分别测定其对二化螟3龄幼虫的生物活性。
香根草提取物的生物活性测定:采用饲料混毒法[18]。利用丙酮将香根草各提取物进行超声溶解,然后加入到人工饲料中配制成浓度为0.05 g·mL-1的含样品饲料,将饲料切成大小形状一致的小块,放入无菌的6孔培养板中,每孔接入5头3龄幼虫,覆盖吸水纸及盖子防止试虫逃逸。每板为1个重复,共重复3次,总计90头试虫。对照采用混入相应比例丙酮的常规人工饲料。所有处理均置于人工气候室内饲养(条件设置同试虫饲养),每3 d记录试虫死亡情况,共记录15 d,以毛笔轻触不动的幼虫记为死亡,其中Fr1—Fr5记录3 d的死亡率。
1.4 水稻和香根草中营养物质含量测定
将剪取的水稻和香根草茎秆自然阴干,分别磨成粉末状。
水溶性蛋白含量测定:采用考马斯亮蓝法[19]。每个样品称取100 mg,加入1 mL pH 7.0的磷酸缓冲液(PBS),充分匀浆后,50℃振荡2 h,然后在4℃,12 000 ×g条件下离心15 min,取40 μL上清液,利用酶标仪(Infinite M200,Tecan)在595 nm下测定水稻和香根草样品中的蛋白含量。以牛血清白蛋白为标准蛋白制作标准曲线,计算各样品中蛋白含量,每个处理重复3次。
纤维素含量测定:采用比色法测定。准确称取样品粉末 100 mg,加入 100 mL预冷的 60%的硫酸(H2SO4),在冰上消化处理30 min,过滤。取滤液2 mL于15 mL的玻璃试管中,加0.5 mL 2%蒽酮试剂和5 mL浓H2SO4,盖上塞子,沸水浴中加热10 min,取出冷却。取250 μL反应液在620 nm下测定OD值,以纤维素标准曲线计算各样品的纤维素含量,每个处理重复3次。
总糖含量测定:采用容量分析法测定。称取样品干粉100 mg,送至农业部农产品及转基因产品质量安全监督检测测试中心完成(杭州),每个处理重复3次。
氨基酸含量测定:采用离子交换色谱法测定。准确称取样品300 mg,送至农业部农产品及转基因产品质量安全监督检测测试中心完成(杭州),每个处理重复3次。
单宁含量测定:采用单宁含量试剂盒(苏州科铭生物技术有限公司)按说明书测定。分别称取水稻和香根草样品粉末100 mg,加入1 mL蒸馏水,充分匀浆后,80℃水浴提取30 min,然后在25℃,8 000×g条件下离心10 min,取15 μL上清液,利用酶标仪分别测定两个样品在760 nm下的吸光度,根据说明书计算单宁含量。每个处理重复3次。
1.5 取食水稻和香根草后二化螟幼虫体内解毒酶活性测定
利用水稻和香根草茎秆饲喂二化螟3龄幼虫,分别于饲喂3、6、9、12和15 d后取样,幼虫饥饿3 h后,在液氮中速冻并立即转置-80℃冰箱中保存,供后续酶活性测定试验用。每处理收集幼虫5头,共3次重复。
羧酸酯酶(CarE)活性测定:参照LU等[20]的方法稍加改进。用0.1 mol·L-1,pH 7.0的PBS制备酶液。每个样品称取100 mg,加入pH 7.0的PBS 1 mL,充分匀浆后,在4℃,12 000×g条件下离心15 min,取40 μL上清液加入96孔酶标板中,每孔加入100 μL底物与显色剂的混合液(用0.2 mol·L-1、pH 6.0的PBS,配制含10 mmol α-乙酸萘酯和1 mmol固蓝RR盐,混合后经过滤得到的滤液即为底物与显色剂的混合液)。然后加入5 μL酶液和45 μL 0.1 mol·L-1、pH 7.6的PBS,利用动力学方法测定10 min内OD450值变化,以反应速度表示酶活力(mol·min-1·mg-1protein)。
谷胱甘肽-S-转移酶(GSTs)活性测定:参照HABIG等[21]的方法稍加改进。用0.1 mol·L-1,pH 6.5的PBS制备酶夜,在96孔酶标板中每孔依次加入45 μL 0.1 mol·L-1、pH 7.6的磷酸缓冲液(PBS)、5 μL酶液(制备方法同蛋白酶)、50 μL 1.2 mmol·L-1的1-氯-2,4-二硝基苯(CDNB)和50 μL 6 mmol·L-1的还原型谷胱甘肽(GSH)。利用动力学方法测定10 min内OD340值变化,以反应速度表示酶活力(nmol·min-1·mg-1protein)。
细胞色素P450酶活性测定:参考ROSE等[22]的方法稍加改进。利用0.1 mol·L-1、pH 7.6的PBS(含1 mmol·L-1EDTA,1 mmol·L-1DTT,1 mmol·L-1苯基硫,1 mmol·L-1PMSF)制备酶液。在96孔酶标板中依次加入2 μmol·L-1对硝基苯甲醚(溶解于0.1 mol·L-1pH 7.8的磷酸缓冲液)50 μL,9.6 mmol·L-1的NADPH 5 μL和45 μL制备好的酶液。利用动力学方法测定10 min内 OD405值变化,以反应速度表示酶活力(nmol·min-1·mg-1protein)。
1.6 取食水稻和香根草后二化螟幼虫体内消化酶活性测定
样品处理及收集同1.5。
蛋白酶活力的测定:参考曹挥等的方法测定[23-24]。将收集的二化螟幼虫转移至预冷的组织研磨器中,加入1 mL pH 7.0 PBS(含0.3%的Triton X-100)研磨,在4℃,12 000×g条件下离心15 min,取上清液作为酶源。取20 µL酶液,在37℃放置5 min,加入50 µL 37℃预热的 0.5%酪蛋白溶液,混匀后 37℃放置 15 min,然后加入50 µL 10%的三氯乙酸,混合后离心(10 000×g,4℃,15 min),取上清液20 µL,放入酶标板内,加入100 µL 0.55 mol·L-1Na2CO3,20 µL Folin-酚试剂,室温放置30 min,利用酶标仪在680 nm下测定OD值。以酪氨酸标准曲线计算蛋白酶活性。
淀粉酶活力测定:参照张炬红等[24]的方法测定。取10 µL 2%的淀粉,20 µL 0.2 mol·L-1PBS(pH 6.0),10 µL酶液(制备方法同蛋白酶),依次加入1.5 mL的离心管内,室温下反应10 min后,37℃水浴60 min,加250 µL 3,5-二硝基水杨酸终止反应,然后沸水浴5 min,自然冷却后,从中取出250 µL样品加在酶标板内,用酶标仪于550 nm波长测定OD值。以麦芽糖标准曲线计算淀粉酶活性。
海藻糖酶活力测定:参考张炬红等[24]的方法。取10 µL 3%的海藻糖、20 µL 0.2 mol·L-1PBS(pH 5.5)和10 µL酶液(制备方法同蛋白酶),依次加入1.5 mL的离心管内,室温下反应10 min后,37℃中水浴60 min,加入250 µL 3,5-二硝基水杨酸终止反应,然后在沸水浴中放置5 min,自然冷却后从中取出250 µL样品加在酶标板内,在酶标仪550 nm下测定OD值。以葡萄糖标准曲线计算海藻糖酶活性。
蔗糖酶活力测定:参考张炬红等[24]的方法。取10 µL 4%的蔗糖、20 µL 0.2 mol·L-1PBS(pH 5.5)和10 µL酶液(制备方法同蛋白酶),依次加入1.5 mL的离心管内,室温下反应10 min后,37℃中水浴60 min,随后加入250 µL 3,5-二硝基水杨酸终止反应,然后沸水浴持续5 min,自然冷却后从中取出250 µL样品加在酶标板内,用酶标仪在550 nm下测定OD值。以葡萄糖标准曲线计算蔗糖酶活性。
1.7 数据统计与分析
采用Microsoft Excel 2010处理数据,采用DPS 14.50软件进行差异显著性分析(P≤0.05)[25]。
2 结果
2.1 香根草活性物质提取及对二化螟 3龄幼虫的生物活性测定
香根草总提取物、水部位、石油醚萃取物、乙酸乙酯萃取物和正丁醇萃取物5种组分对二化螟3龄幼虫的生物活性测定结果表明,乙酸乙酯萃取物处理二化螟3龄幼虫15 d后,死亡率为1.85%,与对照无显著差异;香根草粗提物、水部位和正丁醇萃取物处理后随时间延长,二化螟死亡率逐渐升高,但直至处理15 d后,死亡率均低于20%;石油醚萃取物处理9、12、15 d后,二化螟死亡率分别为38.89%、57.41%、85.19%(图1)。
进一步将石油醚萃取物通过硅胶柱层析,获得编号为Fr1—Fr5的5个组分。生测结果表明,处理3 d后,Fr1对二化螟的死亡率为85.00%,显著高于其他组分。Fr3和Fr5对二化螟幼虫的死亡率分别为48.33%和66.67%,二者无显著差异;Fr2对二化螟幼虫的死亡率为30.00%,与Fr3无显著差异;Fr4对二化螟幼虫的死亡率最低,仅为8.33%(图2)。
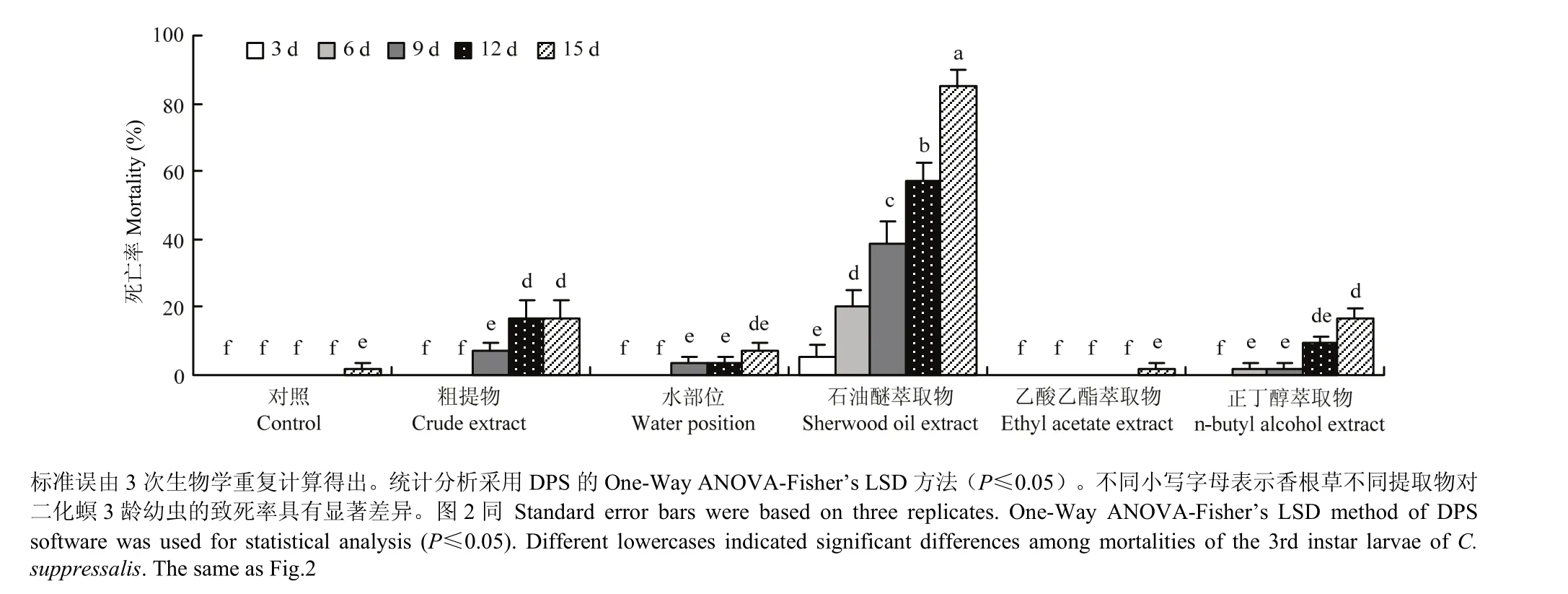
图1 香根草提取物对二化螟3龄幼虫的生物活性Fig. 1 Bioassay of vetiver extracts against the 3rd instar larvae of C. suppressalis
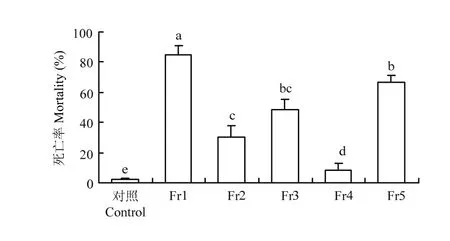
图2 香根草石油醚萃取物中不同组分对二化螟3龄幼虫的生物活性Fig. 2 Bioassay of sherwood oil extracts of vetiver against the 3rd instar larvae of C. suppressalis
2.2 水稻和香根草茎秆中营养物质含量
除单宁外,水稻中其他营养物质含量均显著高于香根草中的含量。其中水稻和香根草中总蛋白的含量分别为13.26和9.45 μg,纤维素含量分别为2.92%和1.61%,总糖含量分别为9.60%和1.45%(图3)。而单宁在香根草中的含量是水稻中含量的 1.31倍(图3)。此外,从水稻和香根草中均鉴定出17种氨基酸,总含量分别为14.23%和4.00%,且这17种氨基酸在水稻中的含量为香根草中含量的2.5—7.0倍。其中水稻中蛋氨酸含量是香根草中的7.0倍(表1)。说明除单宁外,水稻中各种营养物质含量均比香根草中丰富。
2.3 取食水稻和香根草后二化螟幼虫体内解毒酶活
性
对取食水稻和香根草后二化螟幼虫体内解毒酶活性测定结果表明,取食不同寄主植物9、12、15 d后,取食水稻的幼虫体内 CarE活性显著高于取食香根草的幼虫体内CarE活性;GSTs在取食水稻和香根草的幼虫体内无显著差异;细胞色素 P450酶活性则在取食不同寄主植物6—15 d,二者均呈现显著差异(图4)。
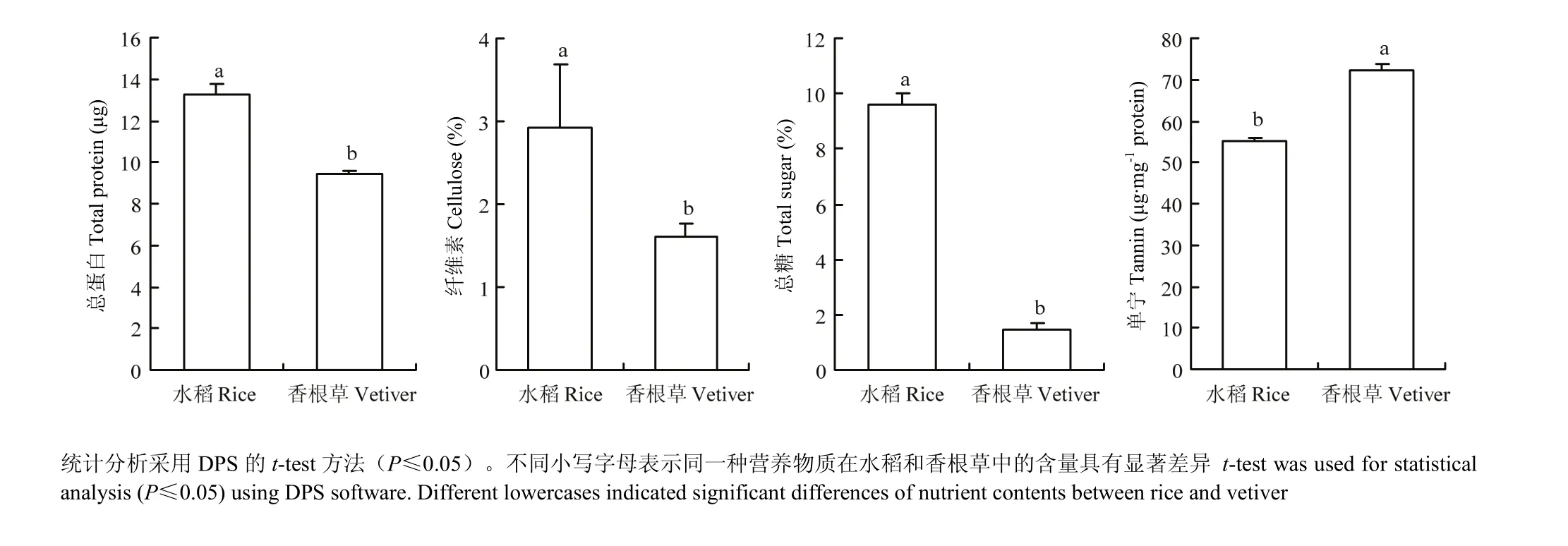
图3 水稻和香根草中的营养物质含量Fig. 3 Nutrient contents in rice and vetiver
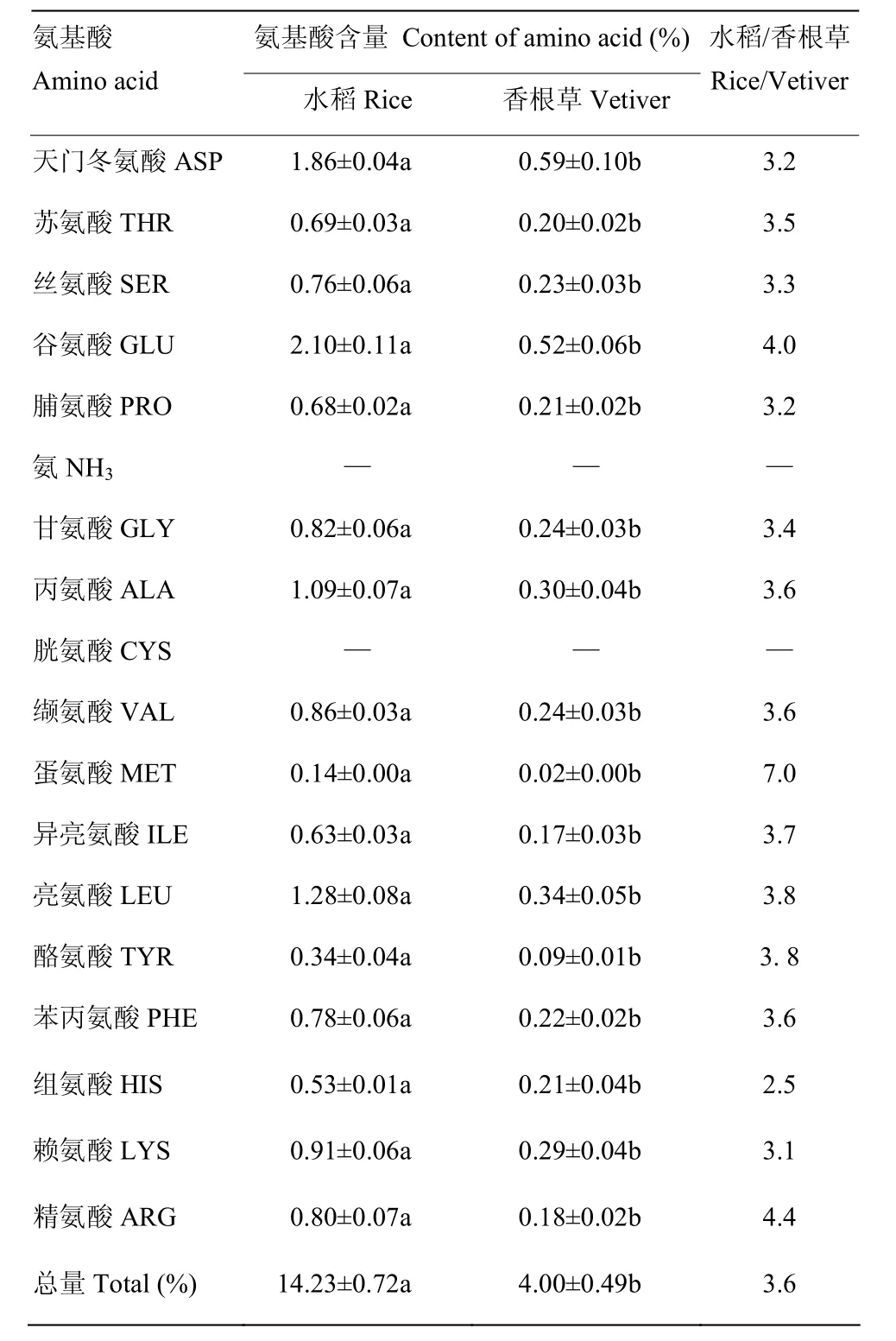
表1 水稻和香根草茎秆中氨基酸种类和含量Table 1 Amino acid compositions and contents in rice and vetiver stem
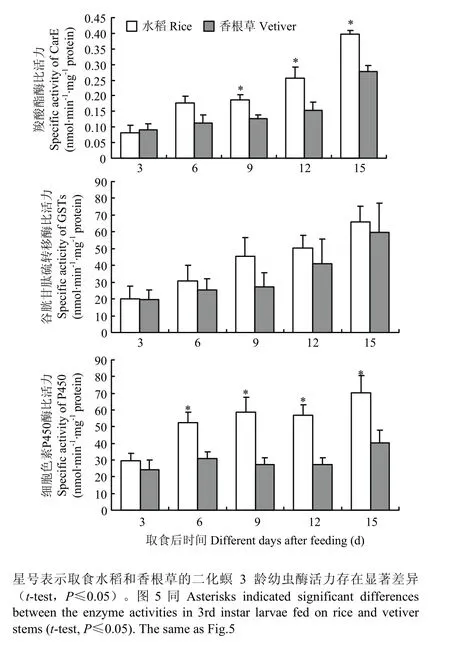
图4 二化螟3龄幼虫取食水稻和香根草后体内解毒酶活性变化Fig. 4 Activity changes of detoxifying enzymes after the 3rd instar larvae feeding on rice and vetiver stems
2.4 取食水稻和香根草后二化螟幼虫体内消化酶活性比较
分别取食不同寄主植物3、6、9、12、15 d后,取食香根草的二化螟幼虫体内蛋白酶、淀粉酶、海藻糖酶和蔗糖酶活性均显著低于取食水稻的二化螟幼虫体内相应酶的活性(图5)。
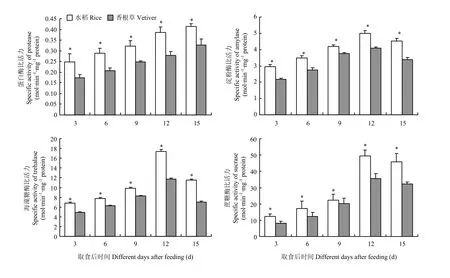
图5 二化螟3龄幼虫取食水稻和香根草后体内消化酶活性变化Fig. 5 Activity changes of digestive enzymes in the 3rd instar larvae after fed on rice and vetiver stems
3 讨论
香根草作为诱集植物防治二化螟已成为稻螟绿色防控的重要措施之一,具有很大的开发潜力[12,14,26-27]。笔者实验室前期研究结果表明,香根草能诱集二化螟雌成虫在其上大量产卵,但孵化出的幼虫在香根草上不能存活[11],因此香根草对二化螟而言属于典型的致死诱集型植物。如果能够开发鉴定出香根草中的杀虫活性物质,明确其对二化螟幼虫致死的作用机制,将为以香根草为基础的稻螟绿色防控新技术提供更充分的理论依据。
本研究从解毒代谢和营养代谢两方面揭示了香根草对二化螟幼虫的致死作用机制。首先,发现香根草的石油醚萃取物对二化螟幼虫具有很强的毒杀作用,15 d后幼虫死亡率高达85.19%,说明香根草中含有对二化螟幼虫具有毒杀作用的活性成分,且存在于石油醚萃取物中;进一步对石油醚萃取物进行分离,在获得的5种组分中,Fr1和Fr5对二化螟幼虫具有较高的致死作用,死亡率分别为85%和67%。笔者实验室前期研究表明,香根草石油醚提取物对二化螟初孵幼虫的毒性较高,3 d后死亡率高达84%左右[13]。这均说明香根草中的杀虫活性物质主要存在于 Fr1和 Fr5组分中,这些有毒物质进入二化螟体内后可能通过抑制其解毒代谢酶活性而降低了对有毒物质的代谢能力,最终引起昆虫中毒死亡。研究还表明,昆虫取食不同寄主植物后对其解毒代谢酶及相关代谢能力的影响不同[28]。昆虫体内参与外来物质代谢的解毒酶主要包括CarE、GSTs和细胞色素P450酶等酶系。本研究表明,香根草中含有对二化螟幼虫具杀虫活性的有毒物质,二化螟取食后,幼虫体内CarE和细胞色素P450酶活性受到显著抑制,导致其对香根草有毒物质的解毒代谢能力下降,引起昆虫死亡。但Fr1和Fr5只是活性成分的粗提物,究竟是哪些活性物质发挥了杀虫作用目前还不能确定,有待进一步研究证实。
此外,香根草中营养成分匮乏可能是二化螟幼虫不能存活的另一重要因素。本研究表明,香根草中总蛋白、纤维素、总糖、氨基酸等营养物质的含量均显著低于水稻中相应物质的含量,而这些营养物质是二化螟生长发育所必需的。一般来讲,植食性昆虫通过取食寄主植物获得营养物质,满足其生长发育的基本营养物质有糖类、蛋白质、氨基酸、纤维素等。其中糖类主要为昆虫的生命活动提供能量需求,纤维素是昆虫的主要碳水化合物[29-30];蛋白质和氨基酸的利用会影响昆虫取食、生长和繁殖[30-31]。香根草中单宁含量高可能是二化螟取食后营养吸收受阻的一个重要因素。单宁对昆虫生长发育具有强烈抑制作用,是常见的植物抗虫物质。单宁与消化酶结合,可抑制消化酶活性,影响其正常生理功能,进而阻碍昆虫对营养成分的吸收[32]。所以,取食香根草的二化螟幼虫可能由于取食植物中的有毒物质或无法利用植物中的营养物质,造成由于营养物质缺乏,结合有毒物质的毒杀作用,最终导致其死亡。
4 结论
香根草中有毒物质抑制了二化螟体内 CarE和细胞色素 P450酶的活性;香根草内营养物质缺失,单宁含量高,从而影响二化螟体内消化酶活性的变化,这些可能是二化螟幼虫取食香根草后致死的两大重要因素。研究结果可为开发以香根草为基础的稻螟绿色防控新技术提供理论依据。
[1] 肖海军, 何海敏, 薛芳森. 二化螟滞育生物学特性的研究进展. 生物灾害科学, 2012, 35(1): 1-6.
XIAO H J, HE H M, XUE F S. Research progress in characteristics of diapause biology in Chilo suppressalis. Biological Disaster Science, 2012, 35(1): 1-6. (in Chinese)
[2] 杨光平, 刘玉娣, 侯茂林. 二化螟滞育幼虫的蛋白和核酸含量以及保护酶活性的变化. 昆虫学报, 2013, 56(3): 251-256.
YANG G P, LIU Y D, HOU M L. Changes of the protein and nucleic acid contents and the activities of protective enzymes in diapausing larvae of the rice stem borer, Chilo suppressalis (Lepidoptera: Pyralidae). Acta Entomologica Sinica, 2013, 56(3): 251-256. (in Chinese)
[3] 鲁艳辉, 赵燕燕, 张发成, 郑许松, 朱平阳, 吕仲贤. 二化螟滞育生物钟蛋白 TIME-EA4基因的克隆及时空和温度诱导表达分析.昆虫学报, 2016, 59(4): 392-401.
LU Y H, ZHAO Y Y, ZHANG F C, ZHENG X S, ZHU P Y, LÜ Z X. Cloning and spatiotemporal and temperature-induced expression profiling of diapause bioclock protein TIME-EA4 gene in the rice stem borer, Chilo suppressalis (Lepidoptera: Pyralidae). Acta Entomologica Sinica, 2016, 59(4): 392-401. (in Chinese)
[4] 彭少兵. 转型时期杂交水稻的困境与出路. 作物学报, 2016, 42(3): 313-319.
PENG S B. Dilemma and way-out of hybrid rice during the transition period in China. Acta Agronomica Sinica, 2016, 42(3): 313-319. (in Chinese)
[5] 姜卫华. 二化螟的抗药性及综合防治研究[D]. 南京: 南京农业大学, 2011.
JIANG W H. Study on insecticide resistance and integrated control of rice stem borer, Chilo suppressalis[D]. Nanjing: Nanjing Agricultural University, 2011. (in Chinese)
[6] 张扬, 王保菊, 韩平, 韩召军. 二化螟抗药性检测方法比较和抗药性监测. 南京农业大学学报, 2014, 37(6): 37-43.
ZHANG Y, WANG B J, HAN P, HAN Z J. Comparison of methods for testing insecticide resistance in Chilo suppressalis and the resistance monitored. Journal of Nanjing Agricultural University, 2014, 37(6): 37-43. (in Chinese)
[7] 胡君, 陈文明, 张真真, 郑雪松, 靳建超, 苏建亚, 高聪芬, 沈晋良.长江流域稻区二化螟抗药性监测. 中国水稻科学, 2010, 24(5): 509-515.
HU J, CHEN W M, ZHANG Z Z, ZHENG X S, JIN J C, SU J Y, GAO C F, SHEN J L. Insecticide resistance monitoring of Chilo suppressalis in the drainage area of the Yangtze River, China. Chinese Journal Rice Science, 2010, 24(5): 509-515. (in Chinese)
[8] 梁齐, 鲁艳辉, 何晓婵, 郑许松, 徐红星, 杨亚军, 田俊策, 吕仲贤.诱集植物在害虫治理中的最新研究进展. 生物安全学报, 2015, 24(3): 184-193.
LIANG Q, LU Y H, HE X C, ZHENG X S, XU H X, YANG Y J, TIAN J C, LÜ Z X. Mini review of the significance of trap crop in insect pest management. Journal of Biosafety, 2015, 24(3): 184-193. (in Chinese)
[9] VAN DEN BERG J, MIDEGA C, WADHAMS L J, KHAN Z R. CanVetiveria zizanioides grass be used to manage insect pests on crops//MAHA C S, CHEN Z, SONG H, LUO F H. Third International Conference on Vetiver-ICV3. Guangzhou, China: Vetiveria zizanioides and Waterrd, 2003: 262-273.
[10] 陈先茂, 彭春瑞, 姚锋先, 关贤交, 王华伶, 邓国强. 利用香根草诱集诱杀水稻螟虫的技术及效果研究. 江西农业学报, 2007, 19(12): 51-52.
CHEN X M, PENG C R, YAO F X, GUAN X J, WANG H L, DENG G Q. Study on technique and effect of vetiver for trapping and killing rice borer. Acta Agriculturae Jiangxi, 2007, 19(12): 51-52. (in Chinese)
[11] 郑许松, 徐红星, 陈桂华, 吴降星, 吕仲贤. 苏丹草和香根草作为诱虫植物对稻田二化螟种群的抑制作用评估. 中国生物防治, 2009, 25(4): 299-303.
ZHENG X S, XU H X, CHEN G H, WU J X, LÜ Z X. Potential function of Sudan grass and vetiver grass as trap crops for suppressing population of stripped stem borer, Chilo suppressalis in rice. Chinese Journal of Biological Control, 2009, 25(4): 299-303. (in Chinese)
[12] 夏岳章, 孙文岳. 香根草对水稻螟虫的诱杀及应用. 浙江农业科学, 2012(12): 1693-1695, 1698.
XIA Y Z, SUN W Y. The trapping function of vetiver grass and its application to rice stem borer. Zhejiang Agricultural Science, 2012(12): 1693-1695, 1698. (in Chinese)
[13] 高广春, 李军, 郑许松, 徐红星, 杨亚军, 田俊策, 吕仲贤. 香根草提取物对二化螟生长发育及体内保护酶活力的影响. 科技通报, 2015, 31(5): 97-101.
GAO G C, LI J, ZHENG X S, XU H X, YANG Y J, TIAN J C, LÜ Z X. Effects of extracts from Vetiveria zizanioides on growth and development, activities of protective enzymes of C. suppressalis. Bulletin of Science and Technology, 2015, 31(5): 97-101. (in Chinese)
[14] LÜ Z X, ZHU P Y, GURR G M, ZHENG X S, CHEN G H, HEONG K L. Rice pest management by ecological engineering: a pioneering attempt in China//HEONG K L, CHENG J A, ESCALADA M M. Rice Planthoppers: Ecology, Management, Socio-Economics and Policy. Springer, 2015: 161-178.
[15] SHELTON A M, BADENES-PEREZ F R. Concepts and applications of trap cropping in pest management. Annual Review of Entomology, 2006, 51: 285-308.
[16] SHELTON A M, NAULT B A. Dead-end trap cropping: a technique to improve management of the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Crop Protection, 2004, 23: 497-503.
[17] 胡阳, 傅强, 罗举, 赖凤香. 二化螟人工饲养的饲料配方及制备方法和大规模饲养方法: CN102356811A[P]. 2012-02-22[2016-07-21].
HU Y, FU Q, LUO J, LAI F X. Artificial diet of Chilo suppressalis and preparation method and mass rearing method: CN102356811A[P]. 2012-02-22[2016-07-21]. (in Chinese)
[18] 王军, 闫超, 蔡彩虹, 冯岗, 张静, 戴好富. 钩枝藤生物碱对斜纹夜蛾幼虫的毒杀活性. 热带作物学报, 2015, 36(9): 1650-1654.
WANG J, YAN C, CAI C H, FENG G, ZHANG J, DAI H F. Insecticidal activity of the alkaloids from Ancistrocladus tectorius against the larvae of Spodoptera litura. Chinese Journal of Tropical Crops, 2015, 36(9): 1650-1654. (in Chinese)
[19] BRADFORD M M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 1976, 72(1/2): 248-254.
[20] LU Y H, GAO X W. Multiple mechanisms responsible for differential susceptibilities of Sitobion avenae (Fabricius) and Rhopalosiphum padi (Linnaeus) to pirimicarb. Bulletin of Entomological Research, 2009, 99: 611-617.
[21] HABIG W H, PABST M J, JAKOBY W B. Glutathione S-transferases: the first enzymatic step in mercapturic acid formation. The Journal of Biological Chemistry, 1974, 249(22): 7130-7139.
[22] ROSE R L, BARBHAIYA L, ROE R M, ROCK G C, HODGSON E. Cytochrome P450-associated insecticide resistance and the development of biochemical diagnostic assays in Heliothis virescens. Pesticide Biochemistry and Physiology, 1995, 51: 178-191.
[23] 曹挥, 刘素琪, 赵莉蔺, 师光禄, 曾鑫年. 瑞香狼毒提取物对山楂叶螨的生物活性及酶活性影响. 林业科学, 2003, 39(1): 98-102.
CAO H, LIU S Q, ZHAO L L, SHI G L, ZENG X N. The effects of the extracts from Stellera chamaejasme L. on the biological and enzyme activity of Tetranychus viennensis Zacher. Scientia Silvae Sinicae, 2003, 39(1): 98-102. (in Chinese)
[24] 张炬红, 郭建英, 万方浩, 夏敬源. 转Bt基因抗虫棉对棉蚜消化酶的风险评价. 华北农学报, 2008, 23(增刊): 238-242.
ZHANG J H, GUO J Y, WAN F H, XIA J Y. Assessment of transgenic Bt insect-resistant cotton on the digestive enzymes of Aphis gossypii (Homoptera: Aphidae). Acta Agriculturae Boreali-Sinica, 2008, 23(Suppl.): 238-242. (in Chinese)
[25] TANG Q Y, ZHANG C X. Data Processing System (DPS) softwarewith experimental design, statistical analysis and data mining developed for use in entomological research. Insect Science, 2013, 20(2): 254-260.
[26] 赵中华, 杨普云. 2011年全国农作物病虫害绿色防控工作进展. 中国植保导刊, 2012, 32(8): 16-19.
ZHAO Z H, YANG P Y. The 2011 national progress in the green prevention and control of diseases and pests of crops. China Plant Protection, 2012, 32(8): 16-19. (in Chinese)
[27] 陈桂华, 朱平阳, 郑许松, 姚晓明, 张发成, 盛仙俏, 徐红星, 吕仲贤. 应用生态工程控制水稻害虫技术在金华的实践. 中国植保导刊, 2016, 36(1): 31-36.
CHEN G H, ZHU P Y, ZHENG X S, YAO X M, ZHANG F C, SHENG X Q, XU H X, LÜ Z X. A demonstration of rice pest management by ecological engineering technologies in Jinhua, China. China Plant Protection, 2016, 36(1): 31-36. (in Chinese)
[28] 陈澄宇, 康志娇, 史雪岩, 高希武. 昆虫对植物次生物质的代谢适应机制及其对昆虫抗药性的意义. 昆虫学报, 2015, 58(10): 1126-1139.
CHEN C Y, KANG Z J, SHI X Y, GAO X W. Metabolic adaptation mechanism of insects to plant secondary metabolites and their implications for insecticides resistance of insects. Acta Entomologica Sinica, 2015, 58(10): 1126-1139. (in Chinese)
[29] 吴坤君, 李明辉. 棉铃虫营养生态学的研究: 食物中糖含量的影响.昆虫学报, 1992, 35(1): 47-52.
WU K J, LI M H. Nutritional ecology of the cotton bollworm, Heliothis armigera (Hübner): effects of dietary sugar concentration on development and reproduction. Acta Entomologica Sinica, 1992, 35(1): 47-52. (in Chinese)
[30] 董洁, 穆利霞, 廖森泰, 邹宇晓, 刘凡, 沈维治, 刘吉平. 部分食用昆虫不同发育时期主要营养成分与活性物质研究综述. 蚕业科学, 2014, 40(4): 737-742.
DONG J, MU L X, LIAO S T, ZOU Y X, LIU F, SHEN W Z, LIU J P. Research progress on main nutritional and active substances of some edible insects at different developmental stages. Science of Sericulture, 2014, 40(4): 737-742. (in Chinese)
[31] MARTIN R C. Reduction of European corn borer (Lepidoptera: Pyralidae) damage by intercropping corn with soybean. Journal of Economic Entomology, 1989, 82(5): 1455-1459.
[32] 王予彤. 青杨诱导抗性及单宁对舞毒蛾营养利用和几种酶活性的影响[D]. 呼和浩特: 内蒙古农业大学, 2015.
WANG Y T. Effects of green poplar induced resistance and tannin on the nutrient utilization and enzyme activities of gypsy moth (Lymantria dispar)[D]. Hohhot: Inner Mongolia Agricultural University, 2015. (in Chinese)
(责任编辑 岳梅)
The Lethal Mechanism of Trap Plant Vetiveria zizanioides Against the Larvae of Chilo suppressalis
LU YanHui1, GAO GuangChun2, ZHENG XuSong1, LÜ ZhongXian1
(1State Key Laboratory Breeding Base for Zhejiang Sustainable Pest and Disease Control, Institute of Plant Protection and Microbiology of Zhejiang Academy of Agricultural Sciences, Hangzhou 310021;2Department of Pharmacy, School of Medicine Science, Jiaxing University, Jiaxing 314001, Zhejiang)
【Objective】 Vetiver grass (Vetiveria zizanioides) can effectively attract female adults of Chilo suppressalis to layeggs, while the larvae can not complete their life cycles by feeding on vetiver. The objective of this study is to illuminate the lethal mechanism of trap plant-vetiver against the larvae of C. suppressalis and provide a theoretical basis for the development of sustainable control technology for C. suppressalis based on the application of trap plant vetiver in rice based ecosystem. 【Method】The toxic substances were extracted from vetiver by the methods of extraction and silica gel column chromatography, and the biological activities of these extracts against 3rd instar larvae of C. suppressalis were determined by mixing them in the artificial diet. The nutrient contents of rice and vetiver, and the detoxifying and digestive enzymes activities of C. suppressalis larvae fed on rice and vetiver were analyzed using biochemical methods. 【Result】 The mortalities of the C. suppressalis 3rd instar larvae after 9, 12, 15 days treated by sherwood oil extract of vetiver were 38.89%, 57.41%, and 85.19%, respectively, which were significantly higher than the mortalities of larvae treated by ethyl acetate extract, n-butyl alcohol extract and other fractions. The fraction 1 (Fr1) and fraction 5 (Fr5) were isolated and obtained from sherwood oil extract of vetiver, and the mortalities of C. suppressalis the 3rd instar larvae to Fr1 and Fr5 (0.05 g·mL-1) were 85.00% and 67.67% after 3 days treatment, respectively. These results indicated that vetiver contains toxic substances, which have lethal effect on C. suppressalis larvae. Results of nutrient contents determination showed that the contents of total protein, cellulose, total sugar and amino acids in vetiver were 9.45 μg, 1.61%, 1.45%, 4.00%, respectively, significantly lower than those in rice, and the methionine (MET) content in rice was 7-fold higher than that in the vetiver. However, the content of tannin was 1.31-fold higher than that in rice. In addition, the protease, amylase, trehalase and sucrase enzyme activities of C. suppressalis larvae on 3rd day after fed (DAF) on vetiver were significantly lower than those in the larvae fed on rice; and on the 6th DAF, the activity of P450 enzyme in the larvae fed on vetiver was significantly lower than that in the larvae fed on rice; and the esterase activity of C. suppressalis larvae on the 9th DAF on vetiver was significantly lower than that in the larvae fed on rice. Compared to glutathione S-transferase (GSTs) activity of the 3rd instar larvae feeding on rice, the enzyme activity of the larvae feeding on vetiver was also decreased, however, there was no significant difference by statistical analysis. 【Conclusion】 Vetiver as a dead-end trap plant of C. suppressalis, there were probably two lethal mechanisms of vetiver against C. suppressalis. Firstly, vetiver contains toxic substances, which have lethal effect on C. suppressalis and these substances inhibited the esterase and cytochrome P450 enzyme activities, leading to the function loss of larvae in detoxification and metabolism. Secondly, compared to rice, lack of nutrients in vetiver can not meet the demand of the larvae, and led to digestive function and physiological activity disorder and ultimately death.
vetiver; Chilo suppressalis; lethal effect; nutrient; detoxifying enzyme; digestive enzyme
2016-07-21;接受日期:2016-11-19
国家重点研发计划(2016YFD0200800)、重点实验室培育基地(2010DS700124-ZZ1601)、国家公益性行业(农业)科研专项(201303017)
联系方式:鲁艳辉,E-mail:luyanhui4321@126.com。通信作者吕仲贤,E-mail:luzxmh@163.com
