新颖语义联结形成的右半球优势效应*
2017-02-01赵庆柏魏琳琳周治金赵黎莉
赵庆柏 魏琳琳 李 瑛 ,3 周治金 赵黎莉 唐 磊
(1青少年网络心理与行为教育重点实验室, 华中师范大学心理学院, 武汉 430079)(2心理学国家级实验教学示范中心, 华中师范大学心理学院, 武汉 430079)(3长沙师范学院学前教育系, 长沙 410100)
1 引言
新颖性(或原创性)是创造活动的一个重要特征。无论是文艺创作、技术创新、管理创新, 还是科学发现等, 都需要提出新颖观念(或思想)。一般认为, 新颖的观念是通过概念(信息)间的新颖联结而形成的。基于这一观点, Mednick (1962)提出, 创造性思维是对已有元素进行重新联结并形成新颖有用的组合的过程, 并据此编制了远距离联想测验。而创造的生成−选择加工模型(Bink & Marsh,2000)则很好地说明了新观念的产生过程。该模型认为, 创造活动是产生−合成和选择这两个综合过程相互作用的结果。在产生−合成阶段, 创造者从相关领域进行广泛的信息搜索, 这些信息的片断临时地组合在一起形成某些新颖的组合; 而在选择阶段, 创造者选择那些新颖且适当的组合体。
除了一般性的创造性理论外, 在创造性思维的具体表现形式上也体现了新颖信息联结形成的重要性。顿悟问题解决是创造性思维的典型形式。顿悟问题解决中, 个体往往会陷入思维僵局, 需要通过问题表征重构或表征变换(破解组块、松懈约束)等方式打破心理定势, 通过形成概念间新颖联结,从而找到解决问题的新方法或新思路。经典的猩猩解决问题、鸽子解决问题, 就是通过形成工具间新颖的联结, 最终解决了问题。顿悟的原型启发理论(张庆林, 邱江, 曹贵康, 2004)也认为, 顿悟问题解决需要发现两个问题之间的相同或相似之处,并在两者之间建立信息间的新颖联结。由此可见,形成信息之间的新颖联结是创造活动的重要加工过程。
由于创造是包含多种基本认知功能的复杂的思维活动, 单纯的行为实验很难对其关键的认知加工过程进行剥离, 因此研究者多采用神经成像手段对创造性问题解决中新颖信息联结形成的认知神经机制进行探讨。研究发现, 新颖信息联结存在右半球优势。Jung-Beeman等人(2004)采用复合远距离联想任务探究顿悟问题解决的神经机制, 功能核磁共振成像(fMRI)结果显示, 相比于非顿悟解决,顿悟解决激活了右半球的颞上回前部; 脑电图(EEG)结果显示, 在顿悟解决前的 0.3 s, 右半球颞叶前部有一个高频率的 gamma波爆发。实验结果表明, 右半球颞上回前部在原本不相关的信息之间建立联系起到了重要作用。Jung-Beeman (2005)采用粗语义编码理论来解释右半球在创造性思维中的作用。该理论认为, 大脑两半球在编码语义信息上的方式是不同的, 左半球采用精细语义编码, 激活与问题解决语义紧密的信息; 而右半球采用粗语义编码, 广泛激活与问题有关的所有信息, 这些信息一般处于无意识水平, 对解决顿悟问题有显著的促进作用。此外, 在谜题类顿悟问题解决中, 研究者同样发现了右半球优势(Shen et al., 2013; 沈汪兵, 刘昌, 张小将, 陈亚林, 2011), 其中右侧海马/海马旁回在新颖联结形成中起关键作用(Luo & Niki,2003; Qiu & Zhang, 2008; Zhao et al., 2013)。连通性分析发现, 顿悟问题解决存在一条从前额−颞叶−海马的右半球神经通路, 负责新颖语义信息的选择和新颖联结的形成(Zhao, Zhou, Xu, Fan, & Han,2014)。
然而, 并非所有的创造性问题解决研究都支持新颖信息联结的右半球优势。在字谜的原型启发范式研究中, 研究者发现相对于非顿悟条件, 顿悟激活了左侧额下/额中回, 认为该脑区与新颖联结产生有关(Qiu et al., 2010); 同样, 在字谜的诱发范式研究中, 字谜语义比常规语义具有更负的 N400成分, 且表现出左半球优势, 研究者认为当去除强的无效的联结并产生弱的新异的联结之后, N400 反映了整合新异联结的过程(邢强等, 2013)。在创造性认知的概念扩展研究中, 研究者发现高原创且高适切的概念扩展激活了双侧额下回、左侧颞极和左侧额极皮层(Kröger et al., 2012), 其中左侧额极脑区负责了概念知识的整合(Abraham et al., 2012)。甚至,在右半球处于加工优势的视觉空间任务中, 研究发现相对于控制任务, 将基本图形联结成新图的创造性任务更多的激活了左侧大脑, 包括后部顶叶皮层、前运动皮层、背外侧前额皮层和内侧前额皮层(Aziz-Zadeh, Liew, & Dandekar, 2013)。类似的, 在面孔加工任务中, 右半球在加工常见面孔时存在优势, 而左半球在加工非常见面孔时表现更佳, 且只有左半球负责的非常见面孔加工与言语和视觉创造力存在相关(Kenett, Anaki, & Faust, 2015)。除此之外, 也有研究指出, 创造性思维加工需要左、右两个半球的共同参与(Mayseless & Shamay-Tsoory,2015; Whitman, Holcomb, & Zanes, 2010)。
对于创造性问题解决中新颖信息联结是否存在右半球加工优势, 先前研究结论并不统一。究其原因, 可能在于:(1)先前研究采用的实验材料类型不同, 涉及言语(Jung-Beeman et al., 2004; Zhao et al.,2013; Zhao, Li, Shang, Zhou, & Han, 2014)、视觉(Aziz-Zadeh etal., 2013; Kenett et al., 2015)以及言语和视觉混合(Qiu et al., 2010)等多种类型。元分析研究发现创造性顿悟具有任务特异性, 不同的实验任务材料会诱发不同的神经活动(Shen et al., 2016)。(2)先前研究中问题解决时间和分析方法存在差异。根据创造的生成−选择加工模型(Bink & Marsh,2000), 创造性问题解决会广泛的激活各种问题相关信息并进行组合, 然后根据任务需求进行选择(保留或舍弃), 其中新颖且有效的信息联结是创造性问题解决的关键。在一步式顿悟问题中, 一旦新颖且有效的信息联结得以产生, 问题便可解决, 也就是说, 新颖信息联结出现在创造性问题解决的晚期, 在问题解决的前一刻。因此, 对新颖信息联结的认知神经机制的探讨应该采用反应锁时分析方法(Jung-Beeman et al., 2004; Zhao et al., 2014), 或者关注刺激锁时分析的晚期成分(赵庆柏, 柯娓,童彪, 周治金, 周宗奎, 2017)。而在先前部分研究(Luo & Niki, 2003; Qiu et al., 2010)中, 顿悟问题的解决时间较短, 只有两三秒钟, 导致低时间分辨率(2s)的 fMRI技术无法将新颖信息联结与思维僵局以及定势打破等过程分离; 此外, 即使在解决时间较长的任务中, 如果对问题解决的不同阶段不做区分, 而是对整个问题解决过程进行分析(fMRI数据分析时长为被试反应时长) (Zhao, Zhou et al., 2014),也可能会淹没新颖语义联结的右半球优势效应。(3)先前部分研究对结果的解释可能存在放大或者偏差。比如, Qiu等人(2010)在字谜的原型启发任务中, 发现顿悟更多的激活了左外侧前额皮层, 认为该脑区与思维定势打破和新颖联结产生有关。由于外侧前额皮层与执行控制和注意转移有关(Kaping,Vinck, Hutchison, Everling, & Womelsdorf, 2011;Ragland et al., 2015), 因此可以认为它在抑制无关信息和转换问题表征中起到关键作用, 但与新颖信息联结产生的关系则相对较远。
由于创造性问题解决往往包含问题表征、思维僵局、定势打破/化解僵局和新颖联结形成等多个过程(Ohlsson, 2011), 且在一步式问题中定势打破和新颖联结形成具有突发性和瞬时性, 因此即使对长时程问题解决采用反应锁时分析能够抓住新颖联结形成的关键过程, 但依然很难将其与思维僵局以及定势打破剥离开来。此外, 任务的复杂性、难度以及被试能力等都会影响到创造性问题解决的认知过程及其结果(Wu, Knoblich, & Luo, 2013), 使得新颖语义联结过程的分离更加困难。
相对于创造性问题解决过程, 创造性言语理解过程更加简单, 更适合用于新颖语义联结研究。隐喻是一种常见且被广泛研究的创造性言语形式, 研究发现新颖隐喻理解同样具有右半球优势。Mashal,Faust, Hendler和 Jung-Beeman (2007)采用 fMRI技术, 比较了加工直意、常用隐喻、新颖隐喻和无关词对的大脑激活模式, 结果发现与常用隐喻和直意词对条件相比, 新颖隐喻激活了大脑右半球颞上沟后部、右侧额下回和左侧额中回, 其中右侧颞上沟后部在言语创造中发挥重要作用。采用相同材料和任务的经颅磁刺激研究发现, 对大脑右侧颞上沟进行刺激将会干扰新颖隐喻的加工, 但对常用隐喻加工则无影响; 而对大脑左侧额下回进行刺激则会削弱直意和常用隐喻的加工, 但对新颖隐喻加工并无影响(Pobric, Mashal, Faust, & Lavidor, 2008)。研究结果表明, 右半球在将两个看似无关概念整合成有意义的隐喻表达过程中发挥重要作用。元分析发现,新颖隐喻的右半球加工优势效应主要发生在句子水平的语义相关性判断任务中, 起到激活广泛语义并整合远距离语义概念的作用(Yang, 2014), 而在新颖隐喻的产生任务中左半球发挥了重要作用, 特别是左背外侧前额皮层负责了策略搜索与选择的执行控制功能(Benedek et al., 2014)。
然而, 在汉语隐喻研究中, 新颖语义联结的右半球优势则未得到很好的支持。王小潞(2007)采用事件相关电位技术(ERP)对新隐喻和死隐喻理解进行比较, 结果发现二者在N400和晚期正成分(LPC)上均未存在显著差异。Tang, Qi, Wang, Jia和Ren(2017)在汉语科学(新颖)隐喻的研究中发现, 汉语科学隐喻所诱发的 N400具有右半球优势, 但创造性思维中的 N400更多反映的是认知冲突的监测;在更能反映语义整合和新颖语义联结产生的晚期成分上, 科学隐喻则未表现出半球优势。此外,Ahrens等人(2007)比较了汉语非常规隐喻、常规隐喻以及直意句子条件下大脑的激活水平差异, 结果发现非常规隐喻比直意句子更多的激活了双侧额叶和颞叶皮层, 比常规隐喻更多的激活了双侧额中回和中央前回以及右侧额上回, 结果并未显示出明显的右半球优势。
由综述可知, 不管是在创造性问题解决还是在创造性言语理解中, 新颖语义联结过程均表现出右半球优势效应, 但也存在不一致的研究结果。这可能是由于先前研究大多主要关注创造性与非创造性思维的差异, 而并未直接指向新颖语义联结形成这一关键子过程, 因此在任务材料类型(言语、视觉或者言语与视觉混合)、任务要求(理解或生成)以及分析方法(刺激锁时或反应锁时)等因素上并未统一,最终导致结果的差异。根据创造的生成−选择加工模型(Bink & Marsh, 2000), 创造问题解决会先后经历信息的产生、综合以及选择等过程, 由于凸显寻常信息往往激活较强(Giora, 1997), 因此在新颖且有效的信息联结形成之前, 要经历多轮的寻常信息联结的产生与摒弃过程(赵庆柏, 李松清, 陈石, 周治金, 成良, 2015); 也就是说, 一步式创造性问题解决包含了漫长的信息搜索与选择以及短暂的灵光一现。相对而言, 在创造性言语理解中, 创新观点的呈现极大的缩短了信息的搜索与选择过程, 更加突显了新颖语义联结的形成过程。因此, 本研究拟选取谜题型歇后语作为实验材料, 直接对语义联结的新颖性进行操控, 分别采用fMRI和ERP技术在空间和时间上对新颖语义联结形成的神经机制,特别是右半球优势效应进行探讨。谜题型歇后语是最能体现歇后语创造性的一种语言形式, 其前语境(引子)像是谜面, 后衬像是谜底, 例如“打靶不在中央——偏心”和“用狗来追踪——文物”。后衬通过语义双关和语音双关等形式激活其非凸显语义, 并与引子之间建立新颖的语义联结(Zhao, Li et al.,2014)。谜题型歇后语的理解过程实际上就是激活非凸显语义进而形成新颖语义联结过程。本研究实验 1和实验 2分别选取谜题型歇后语的阅读理解任务和语义关联性判断任务, 并分别采用 fMRI和 ERP技术, 试图在不同成像手段和不同言语理解任务下验证新颖语义联结右半球加工优势的稳定性。
2 实验1 谜题型歇后语的阅读理解实验
2.1 实验目的
采用谜题型歇后语的阅读理解任务, 初步验证汉语新颖语义联结过程存在右半球优势。
2.2 研究方法
2.2.1 被试
被试为10名在校大学生和研究生(5名男性, 5名女性), 年龄在19~24岁之间, 平均年龄20.56岁。被试身体健康状况良好, 视力或矫正视力正常, 均为右利手, 体内无任何金属植入物, 无精神病史及长期服药史。实验前告知被试注意事项, 并签署知情同意书。
2.2.2 实验设计
采用单因素被试内设计, 自变量为歇后语前语境与靶子词之间的关联类型:新颖语义关联条件和寻常语义关联条件, 因变量为大脑激活强度。
2.2.3 实验材料
实验材料为语义双关歇后语, 在该类歇后语中后衬的引申义是其凸显意义, 其字面意义为非凸显意义, 平时几乎不用。而歇后语的前语境(引子)恰恰是与后衬的非凸显意义存在联系。比如“胡子上贴膏药——毛病”, “毛病”的凸显意义是“差错、问题”, 但是在歇后语中, 需要抑制其凸显意义, 激活其非凸显的字面意义“毛发生病”, 才能在前语境和后衬之间建立起语义联结, 因此这种联结是内隐且新颖的。选取语义双关歇后语60条, 作为新颖语义关联条件的材料, 后衬均为双字词。另请45名学生根据这 60条歇后语的前语境进行自由联想, 要求他们写出看到每条前语境想到的第一个双字词。根据被试的回答, 综合考虑联想频次、词汇熟悉性、词类和情绪效价等几个方面因素来选择寻常语义关联条件的歇后语后衬。比如, “胡子上贴膏药”的寻常语义关联的后衬为“滑稽”。请 23名同学对这60对歇后语的新颖性进行7点评分, 1分代表“一点都不新颖”, 7分代表“非常新颖”。结果显示, 新颖语义关联条件新颖性平均得分为4.97 ± 0.61, 寻常语义关联条件新颖性平均得分为1.32 ± 0.38, 二者差异极其显著, 配对t
(59) = 28.67,p
< 0.001。实验材料分为两组, 每组包含 30句新颖语义关联歇后语和 30句寻常语义关联歇后语, 同一前语境的两条歇后语分属不同的材料组, 每名被试只完成一组实验材料。2.2.4 实验程序
实验采用fMRI组块设计。为了保证被试将注意力集中于歇后语的阅读理解, 实验采用学习−测验任务, 先让被试学习理解一个组块的材料, 包含5个项目, 再呈现一个探测项目, 要求被试判断是否学习过。一个学习−测验单元为一个组块, 根据实验材料组块分为两类:新颖语义关联组(NR)、寻常语义关联组(CR), 各6个组块。在正式实验前告知被试:寻常语义组块的实验材料前后是一种寻常的语义联结, 而新颖语义组块的实验材料前后存有新颖语义联结, 其后将有测验, 因此要注意理解记忆。组块呈现的顺序在被试内平衡, 在被试间随机。每个歇后语项目具体流程为:黑色屏幕上首先出现白色空屏2000 ms, 然后呈现歇后语的前语境(引子)2000 ms, 再呈现后衬2200 ms, 间隔600 ms开始学习下一条歇后语。5条歇后语呈现完后间隔2000 ms,呈现探测项目, 要求被试在4 s内按键判断该项目是否在学习阶段出现过。每个组块需时32 s。两个组块之间, 屏幕中央呈现“+”注视符24 s。具体流程如图1所示。实验前被试需练习4个组块, 以确保完全理解和熟悉实验的任务。练习时, 每次判断后给予被试正误反馈, 正式实验中不提供反馈。
2.2.5 fMRI数据采集

图1 正式实验流程图。实验包含NR和CR两种组块, 每个组块包含若干个歇后语项目, 每个项目用时4200 ms。
使用西门子公司的Siemens Trio 3.0T磁共振扫描仪和配备EPI功能的头线圈进行脑成像扫描。采用磁化准备快速梯度回波序列获取每个被试T1加权像。扫描参数:TR/TE = 9.7 ms / 4 ms, 翻转角9度, 视野(FOV) 256 mm × 256 mm, 轴向 128 层, 层厚1.25 mm。功能性图像采用多次激发梯度回波EPI序列, 在T2加权图像的层面上进行BOLD功能磁扫描, 取32层3.75 mm厚隔层扫描记录全脑活动。成像参数为:平面分辨率3 mm × 3 mm, TR = 2 s,TE = 35 ms, FA = 80°, FOV = 192 mm × 192 mm,Matrix = 64×64。为防止头动, 在头线圈与被试头部之间用海绵块填塞。
2.2.6 脑成像数据分析方法
使用SPM5软件对得到的fMRI功能像数据进行分析处理。首先进行层间时间校正, 参考层选择图像扫描顺序的中间层; 随后进行头动校正, 将每个实验序列中的所有图像与该序列的第一幅图像对齐; 然后将功能像和结构像进行对齐, 分割, 把不同的大脑图像进行2 mm × 2 mm × 2 mm的大小重采样处理, 标准化到MNI的EPI模板上, 最后进行8 mm的高斯平滑处理。数据分析锁定的是谜题型歇后语阅读理解的后衬阶段, 再认阶段不做分析。采用一般线性模型得到新颖语义关联条件、寻常语义关联条件以及二者差异的个体激活图, 并对其进行组分析。由于谜题型歇后语的解决过程相对复杂, 同类材料中不同题目的认知加工过程存在差异, 甚至对于相同题目不同被试的认知加工也可能不完全相同, 因此 fMRI组分析参考成语谜语任务研究(Zhao et al., 2013)采用相对宽松的阈值, 对两个条件的比较, 选择阈值p
< 0.001 (未矫正)且连续激活体素大于20的区域。2.3 结果
如图2所示, 新颖语义关联条件比寻常语义关联条件在右侧颞上回(MNI坐标, [45, 12, −15],t
(9) =8.36)激活更强。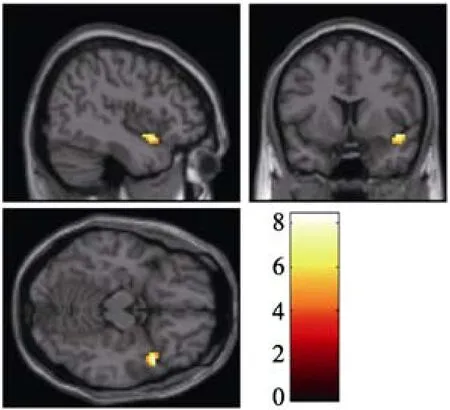
图2 新颖语义关联条件比寻常语义关联条件(NR-CR)比较的脑激活图
2.4 讨论
实验1采用谜题型歇后语的阅读理解任务探讨新颖语义联结的神经机制。实验选取语义双关歇后语作为新颖语义联结的实验材料, 该类歇后语在其语义通达过程中, 需要对后衬词汇的凸显语义进行抑制, 对词汇组块进行破解, 激活每个单字含义,并进行重新组合形成后衬新的非凸显语义, 最终与前语境形成新颖的语义联结。根据右半球粗语义编码理论(Beeman & Bowden, 2000), 大脑左半球激活与问题紧密相关的精细语义信息, 而右半球则负责广泛激活与问题远距离相关的粗语义信息, 因此右半球在语义双关歇后语加工中应发挥重要作用。实验 1结果显示, 相对于寻常语义关联条件, 新颖语义关联条件在右侧颞上回表现出更强的激活。右侧颞叶是言语创造活动的经典脑区, 反映了新颖语义信息的激活与整合, 具有跨材料(复合远距离联想测验、字谜、成语谜语和隐喻等)和任务要求(理解和生成)的稳定性(Jung-Beeman et al., 2004;Mashal et al., 2007; Zhang, Tian, Wu, Liao, & Qiu,2011; Zhao, Li et al., 2014)。实验1结果与先前研究具有较强的一致性, 支持了新颖语义联结的右半球优势加工。
实验1也存在一些不足。首先, 被试样本量并不充足, 可能导致实验结果随机性增大。其次, 组分析选取了相对宽松的统计阈值, 这是由于创造性思维是包含多个基本认知过程的复杂活动, 导致在实验材料和被试思维过程上很难实现较高的同质性, 因此需要宽松的统计阈值才能获得显著的统计结果。尽管如此, 实验 1的结果符合理论预期, 且与先前文献高度一致, 因此是可信的。然而, 实验1采用的是组块设计, 尽管持续重复刺激容易获得关键脑区的神经信号, 但信号重叠也淹没了关键脑区的动态神经加工过程, 加之 fMRI较低的时间分辨率, 导致实验1无法在时间上对新颖语义联结进行锁定。此外, 学习−测验范式存在持续的记忆需求,对创造性言语理解过程可能有所削弱。因此, 实验2拟采用高时间分辨率的ERP技术, 在语义关联性判断任务中对谜题型歇后语的新颖语义联结过程进行探讨。
3 实验2 谜题型歇后语的语义关联性判断实验
3.1 实验目的
采用谜题型歇后语的语义关联性判断任务,考察汉语新颖语义联结形成的时间进程及其神经机制。
3.2 研究方法
3.2.1 被试
16名高校本科生, 其中男生 7人, 女生 9人,年龄介于20至24岁之间, 平均年龄为22.4岁。所有被试均无精神疾病和神经障碍病史, 身体健康,裸眼视力或校正视力正常, 右利手, 实验时精神状态良好。实验前均获得被试知情同意。实验结束后获得一定报酬。
3.2.2 实验设计
采用单因素的被试内设计, 自变量是歇后语前语境与后衬之间的关联类型:新颖语义关联条件和寻常语义关联条件, 因变量为被试进行语义关联性判断过程中相关ERP平均波幅。
3.2.3 实验材料
与实验1相似, 实验材料选取120条语义双关歇后语, 作为新颖语义关联条件的材料, 后衬均为双字词。对于每条歇后语, 另外编写一条与其后衬相同, 但前语境与后衬的凸显意义相关的“歇后语”,作为寻常语义关联条件的材料。比如, 新颖语义关联条件“写字不在行——出格”, 与之对应的寻常语义关联条件为“打扮不常规——出格”。请23名同学对这120对歇后语的新颖性进行7点评分, 1分代表“一点都不新颖”, 7分代表“非常新颖”。结果显示,新颖语义关联条件新颖性平均得分为 5.06 ± 0.72,寻常语义关联条件新颖性平均得分为 1.26 ± 0.40,二者差异极其显著, 配对t
(119) = 50.21,p
< 0.001。将120对歇后语交叉分为两组, 每组包含60条新颖语义关联歇后语和 60条寻常语义关联歇后语, 同一后衬的两条歇后语分属不同的材料组。另外每组添加120条前语境与后衬无语义关联的填充材料。每名被试只完成一组实验材料。3.2.4 实验程序
被试坐在灯光柔和、静音的屏蔽实验室完成实验。刺激由17寸纯平显示器呈现, 短语句文字为黑色, 后衬为绿色, 其余均相同:字体楷体, 字号30,背景为灰色。被试距屏幕约70 cm, 视角约为1°, 屏幕刷新率80 Hz。每个组材料包含240个trial, 分为 2个 block, 每个 block包含 120个 trial, 单个block完成后休息5 min, 然后进入下一个block。实验对语义关联性判断的反应手进行被试间平衡。正式实验开始前进行练习(材料与正式实验不同),练习中对被试反应的正确率和反应时进行反馈, 并对被试反应正确率达不到85%时延长练习次数。正式实验共持续约20 min (不包含中间休息时间)。
实验程序采用E-prime 2.0编制。首先呈现注视点500 ms, 间隔300 ms后呈现歇后语的前语境, 采取从左到右逐词呈现的方式, 每词呈现 400 ms后消失, 随后下一个词在右侧相邻位置呈现, 直至整个前语境呈现完毕。间隔400 ms空屏后, 呈现后衬700 ms, 空屏300 ms后, 屏幕中央呈现问号3000 ms,此时则让被试尽可能准确地按键判断后衬与前语境是否存在着语义关联性。语义相关按“1”键, 语义无关按“2”键。3000 ms内未作答则自动进入下一题,每个trial之间的时间间隔为1000 ms。具体如图3所示。
3.2.5 EEG记录和分析
采用Neuroscan脑电记录仪, 按国际10~20系统扩展的64导Ag/Agcl电极帽记录EEG。实验记录时以左侧乳突为参考电极, 离线处理时将所有电极的数据转化为双侧乳突平均作为参考。垂直眼电(VEOG)由位于左眼上下眶的电极记录, 水平眼电(HEOG)由双眼外侧1 cm处安置的电极记录。每个电极处的头皮电阻保持在5 kΩ以下。滤波带宽为0.05~100 Hz, 连续采样频率为500 Hz/导。完成连续记录 EEG 后离线处理数据, 采用30 Hz低通滤波, 自动校正VEOG和HEOG, 所有导联电压超过± 100 µV视作伪迹予以剔除。
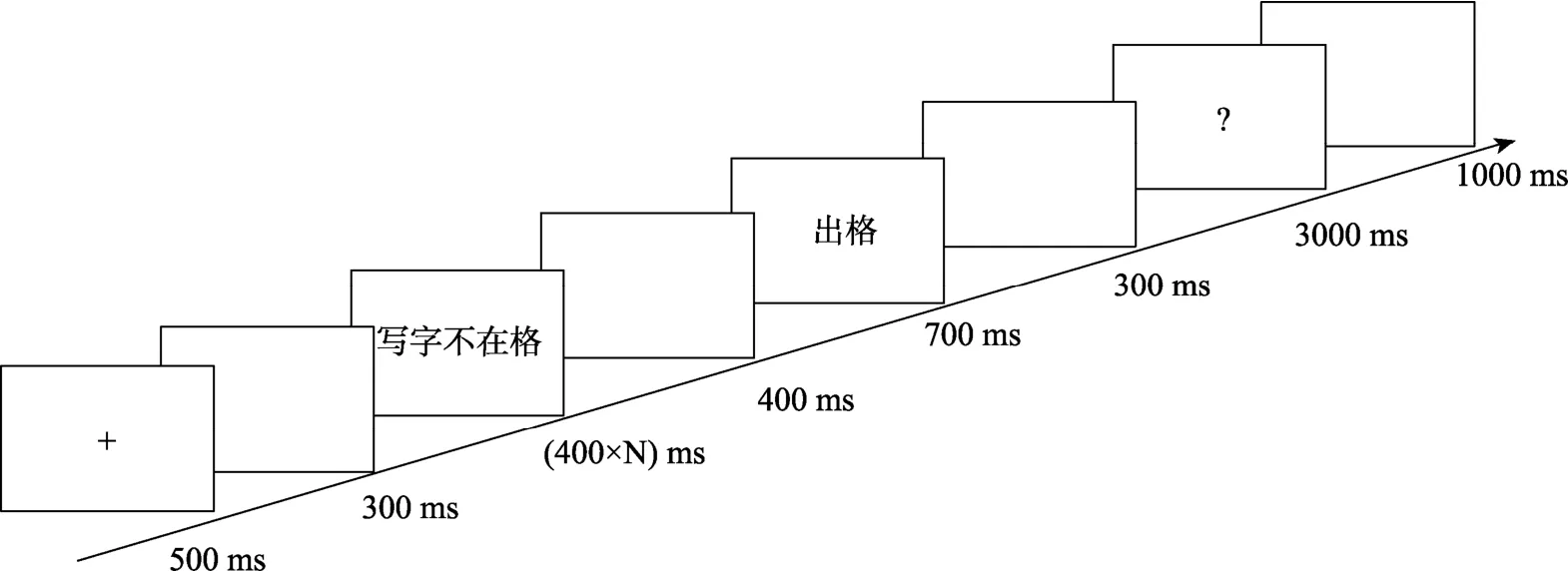
图3 歇后语语义关联性判断实验流程图
实验中所分析脑电的时程均为后衬呈现后的−200~1000 ms, 以−200~0 ms为基线。对新颖语义关联和寻常语义关联条件下引发的脑电分别进行叠加和平均, 发现两种条件在额中央区均诱发一个300~500 ms的负成分, 在右侧额区和颞区均诱发一个650~900 ms的晚期正成分(LPC)。根据波形和地形图, 对额中央区 N300~500的平均波幅进行2(条件:新颖语义关联, 寻常语义关联) × 3(半球:左[F5, FC5, C5], 中[Fz, FCz, Cz], 右[F6, FC6, C6])的重复测量方差分析, 对右侧颞区 LPC平均波幅进行2(条件:新颖语义关联, 寻常语义关联) × 3(电极:FT8, T8, TP8)的重复测量方差分析, 对右侧额区LPC的平均波幅进行了2(条件:新颖语义关联,寻常语义关联) × 4(电极:FP2, AF4, F4, FC4)的重复测量方差分析。方差分析的p
值均用 Greenhouse-Geisser法校正。3.3 结果
1名被试由于正确率低于 85%而被剔除, 剩余有效被试 15名, 正确率均高于 90%。由于实验采用延迟判断反应, 所以对被试进行语义关联性判断的反应时不作分析。
根据 ERP波形(见图 4), 新颖语义关联条件和寻常语义关联条件在额中央区均诱发一个 N300~500成分。用平均波幅法对该成分进行分析。两因素重复测量方差分析显示, 语义关联条件主效应不显著,F
(1, 14) = 0.959,p
= 0.344, η= 0.064; 半球主效应显著,F
(2, 28) = 24.908,p
< 0.001, η= 0.640,其中左半球的平均波幅(−3.067 ± 0.920 μV)比中线(−0.963 ± 1.274 μV)更负, 差异达到边缘显著p
=0.060, 中线平均波幅比右半球(3.285 ± 1.109 μV)显著更负,p
= 0.001; 语义关联条件和半球之间的交互作用不显著,F
(2, 28) = 2.649,p
= 0.104, η=0.159。根据ERP波形和地形图(见图5), 新颖语义关联条件和寻常语义关联条件在右侧额区和右侧颞区均诱发了一个650~900 ms的LPC。用平均波幅法对右侧颞区的LPC (650~900 ms)进行分析。两因素重复测量方差分析显示, 语义关联条件主效应显著,F
(1, 14) = 7.617,p
= 0.015, η= 0.352, 其中新颖语义关联条件的平均波幅(2.630 ± 0.765 μV)显著大于寻常语义关联条件(1.216 ± 0.677 μV); 电极主效应不显著,F
(2, 28) = 0.826,p
= 0.404, η= 0.056;电极与语义关联条件交互作用不显著,F
(2, 28) =0.528,p
= 0.595, η= 0.036。
图4 额中央区N400波形图
采用平均波幅法对右侧额区的LPC进行分析。两因素重复测量方差分析显示, 语义关联条件主效应显著,F
(1, 14) = 5.246,p
= 0.038, η= 0.273, 其中新颖语义关联条件的平均波幅(3.593 ± 0.784 μV)显著大于寻常语义关联条件(1.263 ± 1.140 μV); 电极主效应不显著,F
(3, 42) = 1.298,p
= 0.283, η= 0.085;电极与语义关联条件交互作用不显著,F
(3, 42) =0.882,p
= 0.458, η= 0.059.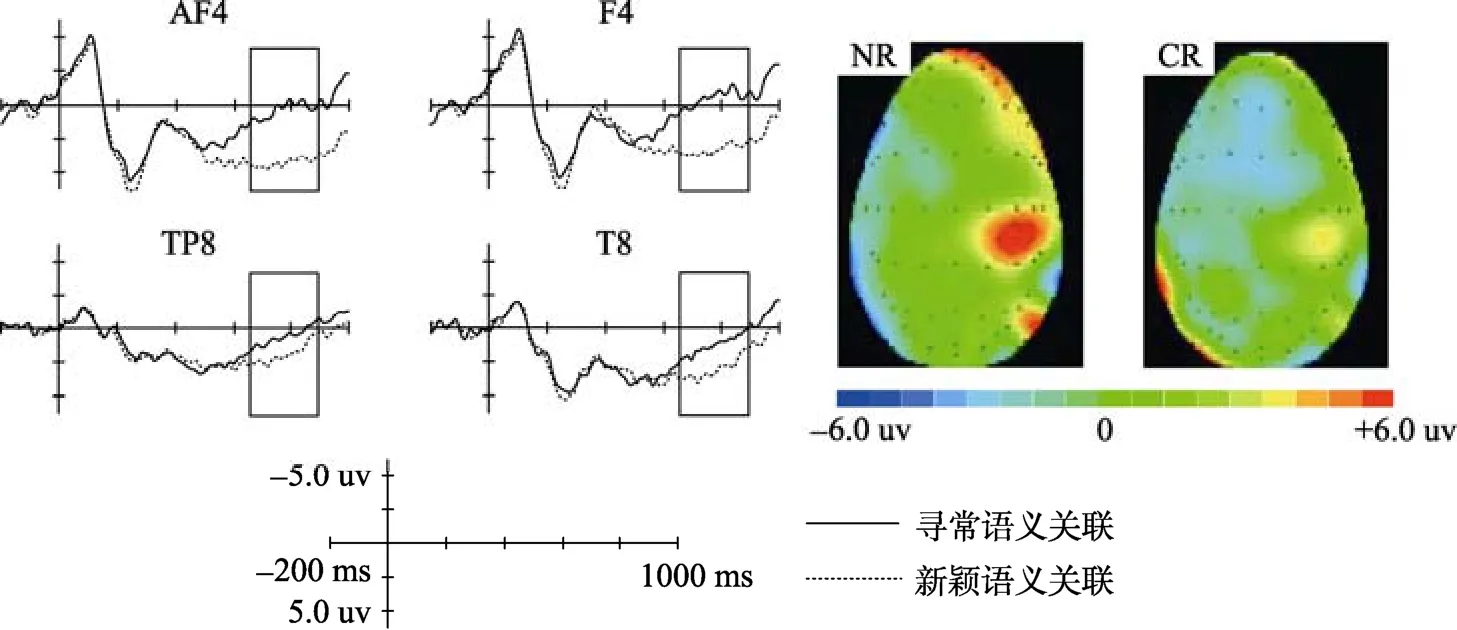
图5 右侧颞叶和额叶的LPC波形及地形图
3.4 讨论
实验2采用事件相关电位技术在谜题型歇后语的语义关联性判断任务中考察新颖语义联结形成的时间进程及神经机制。相对于实验1的阅读理解任务, 实验2要求被试对前语境与后衬词汇的关联性进行判断, 任务直接导向新颖语义联结过程; 而且, 实验采用延迟判断, 为被试提供充足的时间以对歇后语进行充分且深入理解, 同时延迟反应降低了任务按键压迫感, 削弱其对歇后语理解过程的影响。此外, 实验2 ERP分析的时间窗为后衬词汇呈现后的1000 ms, 这正是对谜题型歇后语后衬词汇进行组块破解, 整合单字非凸显含义形成词汇新义,并与前语境建立新颖语义联结的时间。因此, 实验2可以更为有效的探察新颖语义联结的加工机制。
ERP结果显示, 新颖语义关联条件和寻常语义关联条件在额中央区均诱发了一个300~500 ms的负成分。先前关于创造性问题解决的大部分研究均发现, 创造性解决比非创造性解决在 400 ms左右诱发了显著更负的 ERP成分, 该成分往往定位于前扣带回, 反映了创造性思维中认知冲突的监测(Mai, Luo, Wu, & Luo, 2004; Qiu et al., 2006)。不过也有研究并未发现创造性思维和非创造性思维在400 ms左右的负成分上的显著差异(沈汪兵等,2013; 王小潞, 2007)。本研究实验2中, 新颖语义关联和寻常语义关联条件在N300~500的平均波幅上并未表现出显著差异。究其原因可能是:创造性问题解决中400 ms的负成分反映的是认知冲突的监测, 先前研究大多要求被试进行有无顿悟感判断,相对于非顿悟条件, 顿悟条件存在较大的认知冲突,且顿悟感判断可能增强被试对冲突的主观意识, 因此在 400 ms诱发了更强的负成分; 而本研究只是要求被试进行语义关联性判断, 相对于先前研究,认知冲突相对较弱。
根据创造性思维加工理论, 新颖语义联结应该发生在思维加工的晚期。实验2 ERP结果显示, 相对于寻常语义关联条件, 新颖语义关联条件在右侧额区和颞区均诱发了一个更正的晚期正成分LPC。LPC被认为是语义信息整合的重要成分。在一般性的语义违反研究中, Brouwer等人(Brouwer, Fitz, &Hoeks, 2012; Brouwer & Hoeks, 2013)通过对文献总结梳理, 指出 N400反映的是记忆中语义信息的提取, 而 LPC/P600反映的是语义信息的整合。而在创造性言语(字谜、歇后语和网络语言等)加工中,研究者也发现创造性条件比非创造性条件诱发了一个更正的 LPC, 该成分或定位于海马/海马旁回或分布于右侧颞区, 因此被认为反映了新颖语义联结的形成过程(Qiu & Zhang, 2008; Zhao, Li et al.,2014; 赵庆柏等, 2017)。与先前文献一致, 本研究发现的右侧颞区 LPC可能反映了谜题型歇后语加工过程中后衬词汇非凸显新颖语义的激活与整合。对于右侧额区的LPC, 由于外侧前额皮层在保持认知目标并相应分配注意资源过程中发挥重要作用(Luks, Simpson, Feiwell, & Miller, 2002; MacDonald,Cohen, Stenger, & Carter, 2000), 因此本研究认为该成分反映了谜题型歇后语加工中对非凸显新颖语义信息的选择, 起到了执行控制的功能。
4 综合讨论
新颖信息联结形成是创造性思维的重要加工过程。狭义上来说, 新颖联结形成是指在看似不相关的远距离信息之间建立联系; 广义上来讲, 新颖联结形成还应包含新颖信息的选择过程。本研究在广义概念的基础上, 采用fMRI和ERP技术对谜题型歇后语加工的神经活动的时空特征进行记录, 旨在揭示汉语新颖语义联结形成的认知神经机制, 特别是对其右半球加工优势进行验证。实验1采用谜题型歇后语的阅读理解任务, 结果发现相对于寻常语义关联条件, 新颖语义联结条件在右侧颞上回表现出更强的激活; 实验2采用语义关联性判断任务,结果发现新颖语义关联条件在右侧额区和右侧颞区诱发了一个更正的晚期正成分LPC。两个实验结果均支持了汉语新颖语义联结形成的右半球加工优势。下面将结合本研究结果及先前文献, 从理论基础、脑区定位和时间进程三个方面对新颖语义信息形成的右半球加工优势进行讨论。
4.1 新颖语义联结形成的右半球加工优势的理论基础
对新颖语义联结形成过程的阐述, 主要存在于创造性思维相关理论中。早在 20世纪 60年代Mednick (1962)就提出, 创造性思维是对已有元素进行重新联结并形成新颖有用的组合的过程。随后,创造的生成−选择加工模型进一步明确, 创造活动需要从问题相关领域进行广泛的信息搜索, 并将这些信息片段形成临时组合, 而创造性则体现在对新颖且适当的组合的选择上(Bink & Marsh, 2000)。不过这两个理论针对的是更为广泛的创造活动, 其中的信息可能是语义的, 也可能是视觉空间的。具体到新颖语义联结的形成过程, 首先要广泛激活问题相关的各种语义信息, 包含凸显的寻常信息和非凸显的新颖信息。然而, 根据Giora (1997)的层级凸显假说, 凸显的寻常语义信息激活较强, 而非凸显的新颖语义信息则激活较弱。因此, 新颖语义联结的形成需要对凸显语义信息进行抑制, 对非凸显语义信息进行激活(赵庆柏等, 2015)。在创造性思维中,由凸显语义信息形成的组合往往是不符合任务要求的, 因此会产生负反馈, 导致其激活程度降低,根据重配原则, 此消彼长, 这会使得非凸显语义信息激活程度增加, 最终形成对任务解决有用的新颖语义联结(Ohlsson, 2011)。简而言之, 新颖语义联结形成就是选择新颖语义信息并在其间建立新颖联结的过程。
众所周知, 大脑左半球是语言加工的优势半球。然而, Beeman等人通过系列研究(Beeman et al.,1994; Jung-Beeman, 2005)发现, 右半球在语言加工中同样发挥了重要作用, 只是功能与左半球存在差异, 并由此提出右半球粗语义编码理论。该理论指出, 大脑左半球激活较小的语义范围, 主要是与输入词汇紧密相关的概念; 而大脑右半球则激活较大的语义范围, 包含了与输入词汇具有相对较远关联(新颖)的概念。该理论不仅得到了上述创造性语言加工神经成像研究的佐证, 也存在神经解剖学基础。在神经元层面, 右半球锥状神经元的树突比左半球延伸更远, 并产生更多的突触连接, 这有利于更多远距离源的信号输入(Hutsler & Galuske, 2003);在宏观层面, 右半球神经柱和脑区也比左半球具有更大的重叠和更紧密的相互连接(Galuske, Schlote,Bratzke, & Singer, 2000)。这些都支持了右半球的远距离粗语义编码。
创造性思维加工理论指出, 新颖语义联结形成的关键在于词汇多种意义的激活以及远距离新颖语义的选择与整合; 而右半球粗语义编码理论则表明, 大脑右半球负责了远距离新颖语义信息的激活与整合。因此, 两个理论共同支持了新颖语义联结形成的右半球加工优势。
4.2 新颖语义联结形成的关键脑区
新颖语义联结形成, 需要激活关键词汇的多重语义信息, 选择其新颖信息并进行整合, 最终在远距离概念间形成新颖联系。根据先前文献和本研究结果, 这一过程主要涉及右侧颞上/中回、右侧额下回以及海马三个关键脑区。
颞叶皮层是语义加工的关键脑区, 同时负责了文字和图片的语义信息加工(Vandenberghe, Price,Wise, Josephs, & Frackowiak, 1996), 其中右侧颞上/中回在新颖语义联结形成中发挥重要作用。复合远距离联想任务研究发现, 相对于非顿悟条件, 顿悟条件更强的激活了右侧颞上回前部, 认为该区域与远距离相关信息间的联结建立过程有关(Jung-Beeman et al., 2004); 汉语变位字研究则发现, 顿悟条件诱发了一个更正的LPC, 该成分定位于右侧颞上回后部, 可能反映了汉字重组过程新颖联结的形成(Zhang et al., 2011); 而惯用语研究结果显示,新颖的字面含义同时更强的激活了右侧颞中回后部和右侧颞上回前部(Mashal, Faust, Hendler, &Jung-Beeman, 2008)。对于颞叶皮层前部和后部区域的功能划分, Jung-Beeman (2005)提出颞叶皮层后部负责输入词汇的语义激活, 而前部区域则与词汇间的语义整合过程有关。然而, 右侧颞叶的这种功能划分并未得到创造性思维加工相关研究的支持, 特别是右侧颞上回前部的语义整合功能。这是因为, 创造性活动涉及相当复杂的思维加工过程,现有文献研究设计并不能将语义激活与整合过程剥离。而且, Jung-Beeman等人强调的右侧颞上回前部也较少有研究支持, 更多的创造性研究定位于颞叶上/中回后部(Luo & Niki, 2003; Zhao et al., 2013);甚至在同样的复合远距离联想任务中, 顿悟解决比分析解决更强的激活了右侧颞叶后部(尽管文献正文中作者提到的是右侧颞上回前部, 但是从结果表格中的BA分区和顶点坐标数据来看, 该脑区应位于右侧颞上回后部, 本文引用以结果数据为准)(Subramaniam, Kounios, Parrish, & Jung-Beeman,2009)。此外, 单词语义与通用知识整合的研究显示,语义整合可以分为两类, 一类是小的信息块组合成已有的记忆表征, 是原有记忆表征的重现, 主要由颞叶负责; 另一类是将不同信息组合成新的、原有记忆中不存在的意义表征, 主要由额叶负责(Hagoort,Hald, Bastiaansen, & Petersson, 2004)。复合远距离联想任务要求报告与三个问题词汇相关的词汇, 产生的三个复合词或者短语是被试记忆中已存储的,其顿悟体现的是新颖性, 而不是全新的。因此, 相关研究所发现的右侧颞上回前部应该反映的是词组或短语水平的新颖语义的激活, 而右侧颞叶后部则反映的是单词水平的新颖语义激活。本研究实验1结果显示, 新颖语义关联条件比寻常语义关联条件更强的激活了右侧颞上回前部, 这可能反映了后衬词汇重组后新颖语义的激活。
尽管右侧颞叶皮层负责了新颖语义信息的激活, 但是新颖语义联结形成需要双侧颞叶的共同参与(Luo & Niki, 2003; Subramaniam et al., 2009; Zhao et al., 2013)。因为创造并非孤立的特殊活动, 需要分析性思维的广泛参与(Ohlsson, 2011); 同样, 新颖语义联结也并非是只在新颖语义信息之间建立联系, 也需要寻常语义的参与。而且, 根据层级凸显假说(Giora, 1997), 当词汇的非凸显新颖语义与句子情景进行整合之后, 其非凸显性(新颖性)会降低, 相应的神经加工也会从右半球向左半球转移。
不过颞叶只是负责了语义信息的激活, 新颖语义联结形成还要对广泛激活的多重语义信息进行选择与整合, 这一过程需要大脑执行控制系统进行参与。额叶是大脑的执行控制中枢。先前研究发现,外侧前额皮层, 特别是额下回普遍参与了创造性思维加工过程(Abraham et al., 2012; Mashal et al.,2007; Mayseless & Shamay-Tsoory, 2015; Qiu et al.,2010; Zhao, Li et al., 2014)。由于额下回在定向语义检索和执行控制中发挥重要作用(Bookheimer, 2002;Wagner, Paré-Blagoev, Clark, & Poldrack, 2001), 因此被认为是负责语义选择和整合的关键脑区(Jung-Beeman, 2005; Zhu et al., 2013)。不过, 如前所述,尽管大脑左右半球对应脑区功能相似, 但也存在差异。研究发现, 左侧额下回的激活程度与任务对认知表征快速选择的需求程度有关(Miller & Cohen,2001)。在语义加工中, 凸显的精细语义激活较强,往往可以实现自动化快速加工, 因此左侧额下回更可能负责凸显精细语义的选择与整合。而对于右侧颞下回, 当需要选择的信息在右半球激活更强时,该脑区将发挥重要作用(Knutson, Wood, & Grafman,2004); 而且功能连通分析显示, 在创造性思维加工过程中, 脑区之间表现出更强的同侧协同(Zhao,Zhou et al., 2014)。既然右侧颞叶负责了非凸显新颖语义的加工, 那么右侧额下回则更可能负责了新颖语义信息的选择与整合。本研究实验2中, 谜题型歇后语同时在右侧额区和右侧颞区诱发了 650~900 ms的LPC, 这一结果也支持了右侧额区在新颖语义选择和整合中的重要作用。
同样需要说明的是, 本研究强调右侧额下回,是因为相对于左侧额下回在各种语义整合任务的普遍激活, 右侧额下回在新颖语义联结形成中更具有特异性; 而在真实的情境下, 新颖语义联结形成需要双侧额下回的共同参与, 特别是两个脑区的平衡激活(Mayseless & Shamay-Tsoory, 2015)。
新颖语义联结形成不仅需要新颖语义信息的激活、选择与整合, 还要形成有效的语义联结。海马因为在路径巡航和关联记忆中的作用(Luo &Niki, 2005; Redish, 2001), 被认为是新颖语义联结形成的关键脑区(Luo & Niki, 2003; Zhao et al.,2013)。然而, 这一结论主要受到脑筋急转弯、谜语等相关研究的支持, 复合远距离联想相关研究并未发现海马的激活。其原因可能在于, 复合远距离联想任务是让被试找出与三个问题词汇相关的目标词汇, 顿悟性解决主要体现在远距离的联结上, 但是这个任务是人造的研究范式, 问题的解决并未产生有效实用的产品; 而海马的激活在有效性相关加工中发挥重要作用(Huang, Fan, & Luo, 2015)。对于左、右侧海马在新颖语义联结中的作用是否存在差异, 现有研究并未给出明确结论, 有的研究报告的是右侧海马(Luo & Niki, 2003; Qiu & Zhang, 2008),有的是左侧(赵庆柏等, 2017), 还有的是双侧(Zhao et al., 2013)。不过, 从创造性思维加工同侧脑区功能连通优势来推测, 右侧海马可能在新颖语义联结形成中发挥更重要的作用(Zhao, Zhou et al., 2014)。本研究结果并未发现海马的激活, 其可能原因是:海马激活并非贯穿始终, 而是在新颖语义联结形成的时刻, 实验 1采用的组块设计可能淹没了海马的激活; 而实验 2采用的 ERP技术, 其脑电地形图反映的是皮层电位分布, 无法反映大脑深层的海马活动。
4.3 新颖语义联结形成的时间进程
根据创造性思维加工理论、层级凸显理论以及右半球粗语义编码理论, 在创造性问题解决或创造性语言加工初期, 输入词汇的凸显语义应优先激活,并进行整合, 此时左半球处于加工优势; 然而凸显寻常语义的加工并不能满足创造性任务的需求, 因此会产生认知冲突, 从而导致凸显语义的抑制和非凸显新颖语义的激活, 此刻右半球开始逐渐发挥作用; 随后新颖语义信息进行整合, 并最终形成新颖语义联结, 右半球在这一阶段中处于优势加工地位。简而言之, 右半球加工优势始于认知冲突的产生, 止于新颖语义联结的最终形成。
先前关于创造性思维加工的 ERP研究支持了这一推论。首先, 创造性思维加工通常会在关键词汇呈现后300~500 ms时间段内诱发一个更负ERP成分(对于该差异波, 不同的文献报告的有 N320、N380或者N400), 反映了认知冲突的监控, 溯源分析结果显示该成分定位于右侧前扣带回 (Mai et al.,2004; Qiu et al., 2006)。也有研究发现, 该成分定位于右侧额中回, 可能反映的是冲突产生之后的定势转换过程(Shen et al., 2013)。不过本研究中, 新颖语义关联条件与寻常语义关联条件在300~500 ms时间段的 ERP成分并未表现出显著差异, 如前所述,这可能是任务要求导致的认知冲突降低所知。其次,创造性思维加工会在N300~500之后诱发一个晚期正成分LPC。由于材料和任务要求的不同, 该成分脑区定位也并不相同。在某些研究中该成分定位于右侧颞叶皮层, 反映了新颖语义信息的激活与整合(Zhang et al., 2011); 也有研究将该成分定位于右侧海马旁回, 反映了新颖语义联结的形成(Qiu &Zhang, 2008)。此外, 反应锁时分析的研究也发现,在新颖语义联结形成的前一刻, 右侧颞叶皮层表现出更强的激活(Jung-Beeman et al., 2004; Zhao, Li et al., 2014)。
5 结论
本研究采用fMRI和ERP技术, 对谜题型歇后语加工的神经活动的时空特征进行探究, 旨在揭示新颖语义联结形成的神经机制, 特别是右半球加工优势。结果显示, 右侧颞区和右侧额区在谜题型歇后语加工的中晚期发挥重要作用, 分别负责了新颖语义的激活以及选择和整合。研究结果支持了新颖语义联结形成的右半球加工优势。
Abraham, A., Pieritz, K., Thybusch, K., Rutter, B., Kröger, S.,Schweckendiek, J., ... Hermann, C. (2012). Creativity and the brain: Uncovering the neural signature of conceptual expansion.Neuropsychologia, 50
, 1906–1917.Ahrens, K., Liu, H. L., Lee, C. Y., Gong, S. P., Fang, S. Y., &Hsu, Y. Y. (2007). Functional MRI of conventional and anomalous metaphors in Mandarin Chinese.Brain and Language, 100
, 163–171.Aziz-Zadeh, L., Liew, S. L., & Dandekar, F. (2013). Exploring the neural correlates of visual creativity.Social Cognitive and Affective Neuroscience, 8
, 475–480.Beeman, M., Friedman, R. B., Grafman, J., Perez, E.,Diamond, S., & Lindsay, M. B. (1994). Summation priming and coarse semantic coding in the right hemisphere.Journal of Cognitive Neuroscience, 6
, 26–45.Beeman, M. J., & Bowden, E. M. (2000). The right hemisphere maintains solution-related activation for yet-tobe-solved problems.Memory & Cognition, 28
, 1231–1241.Benedek, M., Beaty, R., Jauk, E., Koschutnig, K., Fink, A.,Silvia, P. J., ... Neubauer, A. C. (2014). Creating metaphors:The neural basis of figurative language production.NeuroImage, 90
, 99–106.Bink, M. L., & Marsh, R. L. (2000). Cognitive regularities in creative activity.Review of General Psychology, 4
, 59–78.Bookheimer, S. (2002). Functional MRI of language: New approaches to understanding the cortical organization of semantic processing.Annual Review of Neuroscience, 25
,151–188.Brouwer, H., Fitz, H., & Hoeks, J. (2012). Getting real about semantic illusions: Rethinking the functional role of the P600 in language comprehension.Brain Research, 1446
,127–143.Brouwer, H., & Hoeks, J. C. J. (2013). A time and place for language comprehension: Mapping the N400 and the P600 to a minimal cortical network.Frontiers in Human Neuroscience, 7
, 758Galuske, R. A. W., Schlote, W., Bratzke, H., & Singer, W.(2000). Interhemispheric asymmetries of the modular structure in human temporal cortex.Science, 289
, 1946–1949.Giora, R. (1997). Understanding figurative and literal language: The graded salience hypothesis.Cognitive Linguistics, 8
, 183–206.Hagoort, P., Hald, L., Bastiaansen, M., & Petersson, K. M.(2004). Integration of word meaning and world knowledge in language comprehension.Science, 304
, 438–441.Huang, F. R., Fan, J., & Luo, J. (2015). The neural basis of novelty and appropriateness in processing of creative chunk decomposition.NeuroImage, 113
, 122–132.Hutsler, J., & Galuske, R. A. W. (2003). Hemispheric asymmetries in cerebral cortical networks.Trends Neurosciences, 26
, 429–435.Jung-Beeman, M. (2005). Bilateral brain processes for comprehending natural language.Trends in Cognitive Sciences, 9
, 512–518.Jung-Beeman, M., Bowden, E. M., Haberman, J., Frymiare, J.L., Arambel-Liu, S., Greenblatt, R., ... Kounios, J. (2004).Neural activity when people solve verbal problems with insight.PLoS Biology, 2
, e97.Kaping, D., Vinck, M., Hutchison, R. M., Everling, S., &Womelsdorf, T. (2011). Specific contributions of ventromedial, anterior cingulate, and lateral prefrontal cortex for attentional selection and stimulus valuation.PLoS Biology, 9
, e1001224.Kenett, Y. N., Anaki, D., & Faust, M. (2015). Processing of unconventional stimuli requires the recruitment of the nonspecialized hemisphere.Frontiers in Human Neuroscience,9
, 32.Knutson, K. M., Wood, J. N., & Grafman, J. (2004). Brain activation in processing temporal sequence: An fMRI study.NeuroImage, 23
, 1299–1307.Kröger, S., Rutter, B., Stark, R., Windmann, S., Hermann, C.,& Abraham, A. (2012). Using a shoe as a plant pot: Neural correlates of passive conceptual expansion.Brain Research,1430
, 52–61.Luks, T. L., Simpson, G. V., Feiwell, R. J., & Miller, W. L.(2002). Evidence for anterior cingulate cortex involvement in monitoring preparatory attentional set.NeuroImage, 17
,792–802.Luo, J., & Niki, K. (2003). Function of hippocampus in"insight" of problem solving.Hippocampus, 13
, 316–323.Luo, J., & Niki, K. (2005). Does hippocampus associate discontiguous events? Evidence from event-related fMRI.Hippocampus, 15
, 141–148.MacDonald, A. W., Cohen, J. D., Stenger, V. A., & Carter, C. S.(2000). Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control.Science,288
, 1835–1838.Mai, X. Q., Luo, J., Wu, J. H., & Luo, Y. J. (2004). "Aha!"effects in a guessing riddle task: An event-related potential study.Human Brain Mapping, 22
, 261–270.Mashal, N., Faust, M., Hendler, T., & Jung-Beeman, M. (2007).An fMRI investigation of the neural correlates underlying the processing of novel metaphoric expressions.Brain and Language, 100
, 115–126.Mashal, N., Faust, M., Hendler, T., & Jung-Beeman, M. (2008).Hemispheric differences in processing the literal interpretation of idioms: Converging evidence from behavioral and fMRI studies.Cortex, 44
, 848–860.Mayseless, N., & Shamay-Tsoory, S. G. (2015). Enhancing verbal creativity: Modulating creativity by altering the balance between right and left inferior frontal gyrus with tDCS.Neuroscience, 291
, 167–176.Mednick, S. (1962). The associative basis of the creative process.Psychological Review, 69
, 220–232.Miller, E. K., & Cohen, J. D. (2001). An integrative theory of prefrontal cortex function.Annual Review of Neuroscience,24
, 167–202.Ohlsson, S. (2011).Deep learning: How the mind overrides experience
. New York: Cambridge University Press.Pobric, G., Mashal, N., Faust, M., & Lavidor, M. (2008). The role of the right cerebral hemisphere in processing novel metaphoric expressions: A transcranial magnetic stimulation study.Journal of Cognitive Neuroscience, 20
, 170–181.Qiu, J., Li, H., Jou, J., Liu, J., Luo, Y. J., Feng, T. Y., ... Zhang,Q. L. (2010). Neural correlates of the "Aha" experiences:Evidence from an fMRI study of insight problem solving.Cortex, 46
, 397–403.Qiu, J., Li, H., Luo, Y. J., Chen, A. T., Zhang, F. H., Zhang, J.M., ... Zhang, Q. L. (2006). Brain mechanism of cognitive conflict in a guessing Chinese logogriph task.Neuroreport,17
, 679–682.Qiu, J., & Zhang, Q. L. (2008). "Aha!" effects in a guessing Chinese logogriph task: An event-related potential study.Chinese Science Bulletin, 53
, 384–391.Ragland, J. D., Ranganath, C., Phillips, J., Boudewyn, M. A.,Kring, A. M., Lesh, T. A., ... Carter, C. S. (2015).Cognitive control of episodic memory in Schizophrenia:Differential role of dorsolateral and ventrolateral prefrontal cortex.Frontiers in Human Neuroscience, 9
, 604.Redish, A. D. (2001). The hippocampal debate: Are we asking the right questions?Behavioural Brain Research, 127
, 81–98.Shen, W. B., Liu, C., Yuan, Y., Zhang, X. J., & Luo, J. (2013).Temporal dynamics of mental impasses underlying insightlike problem solving.Science China Life Sciences, 56
,284–290.[沈汪兵, 刘昌, 袁媛, 张小将, 罗劲. (2013). 顿悟类问题解决中思维僵局的动态时间特性.中国科学: 生命科学, 43
,254–262.]Shen, W. B., Liu, C., Zhang, X. J., & Chen, Y. L. (2011). The time course and hemispheric effect of “insight” in threecharacter Chinese riddles task: An ERP study.Acta Psychologica Sinica, 43
, 229–240.[沈汪兵, 刘昌, 张小将, 陈亚林. (2011). 三字字谜顿悟的时间进程和半球效应: 一项 ERP研究.心理学报, 43
,229–240.]Shen, W. B., Liu, C., Zhang, X. J., Zhao, X. J., Zhang, J., Yuan,Y., & Chen, Y. L. (2013). Right hemispheric dominance of creative insight: An event-related potential study.Creativity Research Journal, 25
, 48–58.Shen, W. B., Yuan, Y., Liu, C., Zhang, X. J., Luo, J., & Gong,Z. (2016). Is creative insight task-specific? A coordinatebased meta-analysis of neuroimaging studies on insightful problem solving.International Journal of Psychophysiology,110
, 81–90.Subramaniam, K., Kounios, J., Parrish, T. B., & Jung-Beeman,M. (2009). A brain mechanism for facilitation of insight by positive affect.Journal of Cognitive Neuroscience, 21
,415–432.Tang, X. M., Qi, S. Q., Wang, B. T., Jia, X. J., & Ren, W.(2017). The temporal dynamics underlying the comprehension of scientific metaphors and poetic metaphors.Brain Research,1655
, 33–40.Vandenberghe, R., Price, C., Wise, R., Josephs, O., &Frackowiak, R. S. J. (1996). Functional anatomy of a common semantic system for words and pictures.Nature,383
, 254–256.Wagner, A. D., Paré-Blagoev, E. J., Clark, J., & Poldrack, R. A.(2001). Recovering meaning: Left prefrontal cortex guides controlled semantic retrieval.Neuron, 31
, 329–338.Wang, X. L. (2007).Neural mechanism of Chinese metaphorial cognition
(Unpublished doctorial dissertation). Zhejiang University.[王小潞. (2007).汉语隐喻认知的神经机制研究
(博士学位论文). 浙江大学.]Whitman, R. D., Holcomb, E., & Zanes, J. (2010).Hemispheric collaboration in creative subjects: Crosshemisphere priming in a lexical decision task.Creativity Research Journal, 22
, 109–118.Wu, L., Knoblich, G., & Luo, J. (2013). The role of chunk tightness and chunk familiarity in problem solving:Evidence from ERPs and fMRI.Human Brain Mapping, 34
,1173–1186.Xing, Q., Zhang, Z. L., Wang, M. R., Zhang, J. L., Wang, J.,Yao, Y. F., & Zhan, D. L. (2013). The electrophysiological mechanisms of constraint relaxation studied with a Chinese logogriph task.Acta Psychologica Sinica, 45
, 508–516.[邢强, 张忠炉, 王梦偌, 张金莲, 王菁, 姚艳芬, 占丹玲.(2013). 汉字字谜任务中限制解除的电生理机制.心理学报, 45
, 508–516.]Yang, J. (2014). The role of the right hemisphere in metaphor comprehension: A meta-analysis of functional magnetic resonance imaging studies.Human Brain Mapping, 35
,107–122.Zhang, M., Tian, F., Wu, X., Liao, S., & Qiu, J. (2011). The neural correlates of insight in Chinese verbal problems: An event related-potential study.Brain Research Bulletin, 84
,210–214.Zhang, Q. L., Qiu, J., & Cao, G. K. (2004). A review and hypothesis about the cognitive mechanism of insight.Psychological Science, 27
, 1435–1437.[张庆林, 邱江, 曹贵康. (2004). 顿悟认知机制的研究述评与理论构想.心理科学, 27
, 1435–1437.]Zhao, Q. B., Ke, W., Tong, B., Zhou, Z. J., & Zhou, Z. K.(2017). Creative processing of internet language: Novel N400 and LPC.Acta Psychologica Sinica, 49
, 143–154.[赵庆柏, 柯娓, 童彪, 周治金, 周宗奎. (2017). 网络语言的创造性加工过程: 新颖N400与LPC.心理学报, 49
, 143–154.]Zhao, Q. B., Li, S. Q., Chen, S., Zhou, Z. J., & Cheng, L.(2015). Dynamic neural processing mode of creative problem solving.Advances in Psychological Science, 23
,375–384.[赵庆柏, 李松清, 陈石, 周治金, 成良. (2015). 创造性问题解决的动态神经加工模式.心理科学进展, 23
, 375–384.]Zhao, Q. B., Li, Y., Shang, X. L., Zhou, Z. J., & Han, L.(2014). Uniformity and nonuniformity of neural activities correlated to different insight problem solving.Neuroscience,270
, 203–211.Zhao, Q. B., Zhou, Z. J., Xu, H. B., Chen, S., Xu, F., Fan, W.L., & Han, L. (2013). Dynamic neural network of insight: A functional magnetic resonance imaging study on solving Chinese ‘chengyu’ riddles.PLoS One, 8
, e59351.Zhao, Q. B., Zhou, Z. J., Xu, H. B., Fan, W. L., & Han, L.(2014). Neural pathway in the right hemisphere underlies verbal insight problem solving.Neuroscience, 256
,334–341.Zhu, Z. D., Feng, G. Y., Zhang, J. X., Li, G. C., Li, H., & Wang,S. P. (2013). The role of the left prefrontal cortex in sentence-level semantic integration.NeuroImage, 76
, 325–331.