7个黑木耳菌株遗传差异性分析*
2017-01-11路新彦郑巧平宋小亚丽水市农业科学研究院浙江丽水323000
路新彦,刘 昆,蒋 俊,郑巧平,宋小亚(丽水市农业科学研究院,浙江 丽水 323000)
7个黑木耳菌株遗传差异性分析*
路新彦,刘 昆,蒋 俊,郑巧平**,宋小亚
(丽水市农业科学研究院,浙江 丽水 323000)
采用拮抗试验、酯酶同工酶和分子标记对引进7个黑龙江主栽黑木耳菌株进行遗传差异分析。结果表明,在黑木耳遗传差异性分析中,3种方法具有一致性。通过酯酶同工酶与分子标记聚类分析,均能将来自于黑龙江同一地域的黑木耳分为4个类别:黑威9号与981遗传距离最近为第一类,黑威10号、8808与第一类2个菌株遗传距离较近为第二类,黑29与第一类、第二类4个菌株遗传距离较远为第三类,而9809、H916与以上5个菌株遗传距离最远为第四类。同时在拮抗试验中黑29、9809、H916三个遗传距离较远的菌株也表现出明显的拮抗现象。研究结果为黑木耳新品种引选以及种质资源评价分析提供技术支撑。
黑木耳;拮抗;酯酶同工酶;分子标记
黑木耳(Auricularia auricula) 是我国广泛人工栽培的食用菌种类之一,种质资源丰富。目前栽培菌株主要通过野生菌株驯化而来,生产用种来源很多,各地相互引种频繁,菌株命名混乱。采用传统的生物学形态进行鉴别,周期长,易受环境因素和生理状况的影响,尤其对形态差异较小的菌株很难鉴别。如何简便、快速、准确地将黑木耳菌株区分开来,已成为广大科研工作者研究的主要内容。分别用酯酶同工酶和分子标记方法进行黑木耳菌株遗传差异性分析的文章已有报道[1-3],但同时采用3种方法比较分析的文章尚未报道。本文通过拮抗、酯酶同工酶和分子标记对7个黑龙江黑木耳菌株进行遗传差异性分析,旨在为黑木耳品种引选和种质资源评价的研究提供理论依据。
1 材料与方法
1.1 试验材料
供试的黑木耳7个菌株分别为黑威9号、黑威10号、黑29、8808、981、9809和H916,均由黑龙江省科学院应用微生物研究所提供。
1.2 试剂与仪器
1.2.1 培养基
固体PDA培养基和液体PDA培养基。
1.2.2 试剂与仪器
本试验所用的RAPD、ISSR引物由上海生工生物工程公司合成。移液器、离心机 (Eppendorf5424)、PCR仪(Eppendorf,Mastercycler)、电泳仪(北京六一仪器厂,DYY-8C型)、凝胶成像系统(BIO-RAD,Gel Doc XR+)。
1.3 拮抗试验
将供试菌株两两接种于PDA平板培养基上,26℃下培养25 d,观察不同菌株间是否有拮抗线的存在和拮抗线是否明显。
1.4 DNA提取
1.4.1 菌丝培养收集
用接种耙将PDA斜面菌种耙碎,接种于PDA液体培养基中,每个菌株各2瓶,于25℃下,摇床120 r·min-1培养至菌丝球生长较密,收集菌丝0.5 g,于-20℃冰箱中保存备用。
1.4.2 DNA提取
采用CTAB法提取DNA。
1.5 差异性检测
PCR扩增反应体系和程序参照农业部相关标准,分别为NYT 1743-2009食用菌菌种真实性鉴定RAPD法和NYT 1730-2009食用菌菌种真实性鉴定ISSR法。
1.6 酯酶同工酶电泳
制胶、电泳、染色方法参照中华人民共和国农业行业标准NY/T1097-2006食用菌菌种真实性鉴定酯酶同工酶电泳法。
1.7 聚类分析
用NTYSYpc2.1软件计算相似系数,得相似性系数矩阵,用UPGMA法进行聚类分析,生成聚类图。
2 结果及分析
2.1 菌丝拮抗试验结果
拮抗试验得到21个组合,存在拮抗现象:黑29、9809、H916三个菌株拮抗明显,9809和8808、 981两两拮抗明显,说明它们彼此不亲和。拮抗试验情况见图1。

图1 不同菌株间拮抗现象Fig.1 Antibiosis among different strains
拮抗现象不明显的有:黑威9号与黑29、8808、981、9809、H916;黑威10号与9808;H916与黑威10号、8808、981。说明它们有一定的亲和力,但不能确定它们的遗传关系。无拮抗现象的有:黑威10号与黑威9号、黑29、8808、981;981与黑29、8808。说明它们彼此亲和力强,遗传较相近,具体见表1。试验结果分析发现981、黑威10号与黑29三者两两没有拮抗现象,亲和性强,表明他们的遗传距离接近。
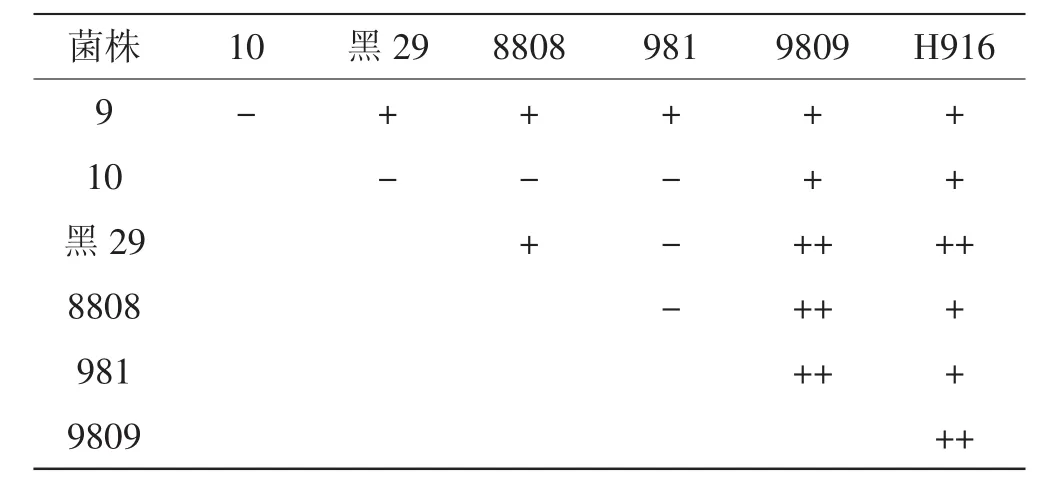
表1 7个黑木耳菌株拮抗试验结果Tab.1 Anatagonistic effect of 7 Auricularia auricula strains
2.2 酯酶同工酶试验结果
2.2.1 酯酶同工酶酶谱及其迁移率
酯酶同工酶对7个不同黑木耳菌株的扩增图谱见图2。

图2 酯酶同工酶对7个黑木耳菌株的扩增图谱Fig.2 Esterase isozyme patterns of 7 Auricularia auricula strains
由图2可知,在整个酶带分布区间内各菌株产生酶带的数量和酶带强弱有一定的差异,这表明各菌株之间还是存在一定的遗传距离。7个黑木耳菌株分别出现8条~11条酶带,迁移率(Rf)在0.286~0.792。当Rf在623与0.688处,酶谱上均有活性很强的酶带,可能为黑木耳的基础酶谱带。当Rf在0.455、0.539、0.623、0.688处,7个菌株分别有出现酶带,且为多态性特征带。当Rf在0.773和0.792处,9809和H916两个菌株没有出现酶带,而其他5个菌株都有明显酶带。
2.2.2 菌株间相似系数
按Nei或Li(1997)公式计算菌株间的相似系数Sc[4],见表2。
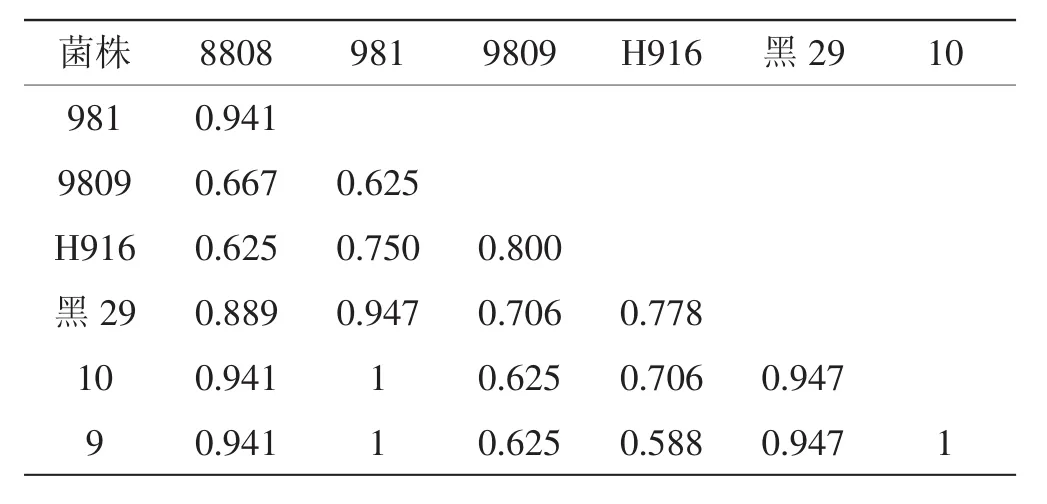
表2 7个黑木耳菌株酯酶同工酶酶谱的相似系数Tab.2 Ester ase isoenzyme similarity coefficient of 7 Auricularia auricula strains
由表2可知,7个黑木耳菌株之间的相似系数(Sc)在0.588~1,相似系数都较大,说明菌株亲缘关系都很近[5],也许因为它们来自于相同的地理区域。其中黑威9、黑威10号与981菌株的相似系数Sc为1,说明这三个菌株亲缘关系非常近,有相同或者相近的遗传基础。
2.2.3 7个黑木耳菌株的酯酶同工酶条带聚类分析
通过NTYSYpc2.1软件分析得到黑龙江7个黑木耳菌株的聚类分析树状图,见图3。

图3 7个黑木耳的酯酶同工酶聚类分析图Fig.3 Dendrogram based on esterase isozyme cluster analysis of 7 Auricularia auricula strains
由图3可知,当相似系数Sc为0.889时,可将7个黑木耳菌株分为4个类别:H916为第一类,9809为第二类,黑29为第三类,8808和981、黑威9号及黑威10号为第四类。
2.3 分子标记试验结果
2.3.1 引物筛选和PCR扩增结果
通过使用检测标准中所提供的22个RAPD引物和28个ISSR引物,对7个供试黑木耳菌株进行分析,筛选得到8个RAPD引物和10个ISSR引物,扩增获得分辨率高、反应稳定性较好和多态性丰富的条带,具体见表3。
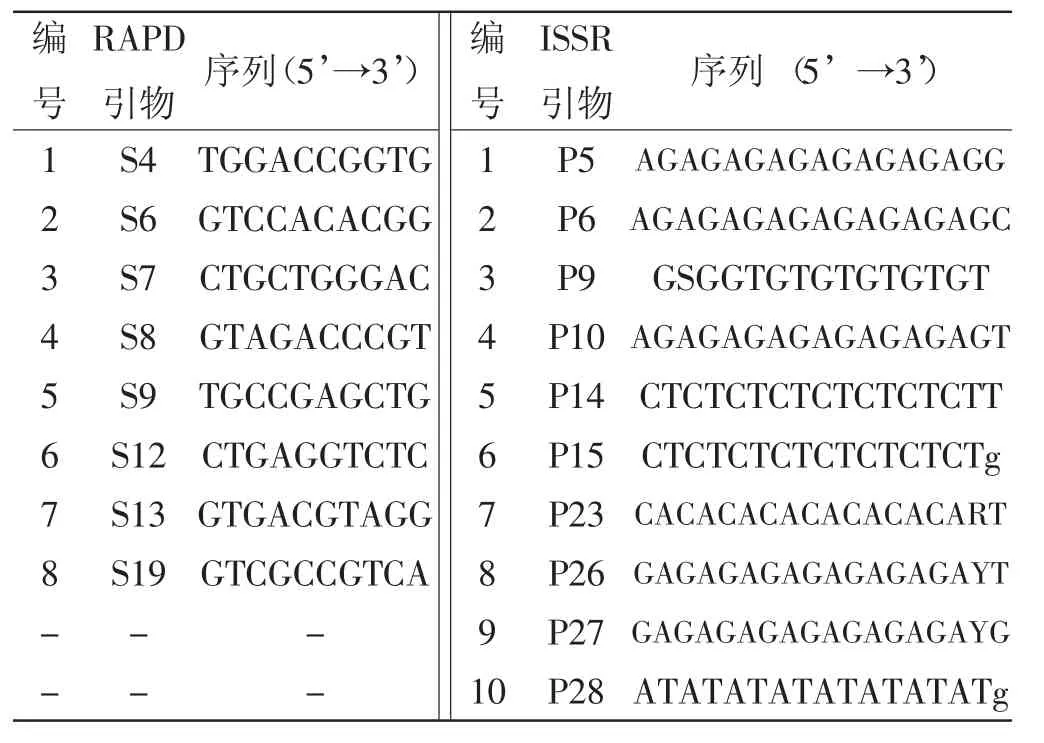
表3 试验引物信息Tab.3 Information of primers
试验共扩增出条带185条,大小分布在500 bp~2 000 bp,其中多态性条带132条,多态性比率为71.35%,部分图谱结果见图4、图5。
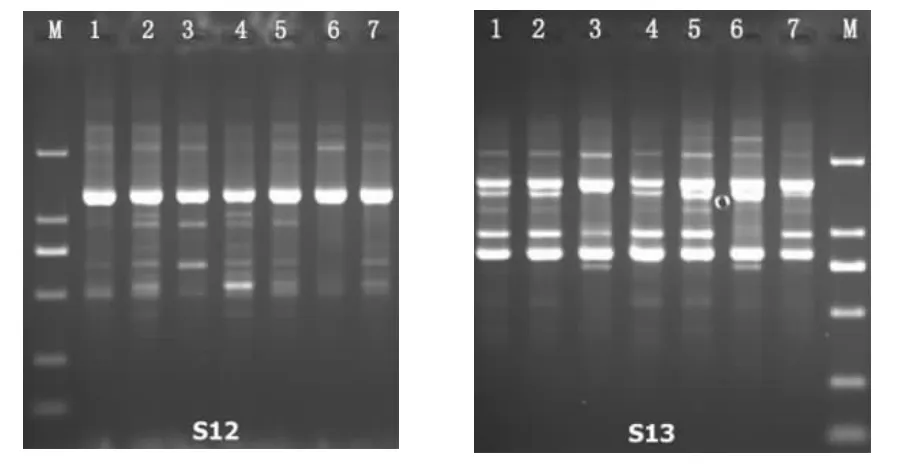
图4 RAPD引物S12、S13对7个黑木耳菌株的扩增图Fig.4 RAPD amplification patterns of primers S12 and S13
2.3.2 聚类分析
使用NTSYSpc-2.1软件计算菌株间的Nei遗传距离为0.61~0.89,应用UPGMA方法进行的遗传距离聚类分析,得到7个菌株亲缘关系树状图见图6。
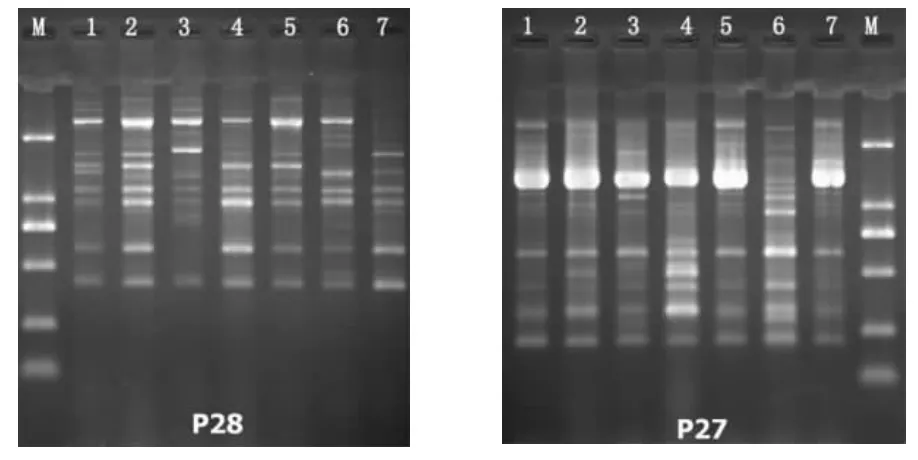
图5 ISSR引物P28、P29对7个黑木耳菌株的扩增图Fig.5 ISSR amplification patterns of primers P28 and P29

图6 7个黑木耳的UPGMA聚类图Fig.6 UPGMA tree of 7 Auricularia auricula strains
从图6中可以看出,相似系数在0.825水平将7个菌株分为4个类群:981与黑威9号为第一类;黑威10号与8808为第二类;H916为第三类;9809为第四类。
3 讨论
从本试验结果可以发现在黑木耳遗传差异性分析中,3种方法具有一致性。酯酶同工酶与分子标记的聚类结果比较,均能发现黑威9号与981遗传距离最近,这两个菌株与黑威10号、8808遗传距离较近,黑29与以上4个菌株较远,而9809、H916与以上5个菌株遗传距离最远。同时在拮抗试验中黑29、9809、H916三个菌株拮抗明显,9809和8808、981两两拮抗明显,它们彼此不亲和,在此对上述结论进行了更充分的验证。
但试验中发现由于酯酶活性易受到外界因素的影响,酯酶同工酶的稳定性以及试验的可重复性不高。分子标记是直接从DNA水平上来检测基因组的多态性[6],不受环境条件和发育阶段的影响,重复性好,适合用于黑木耳生产菌株快速准确鉴定。菌丝拮抗有着其直观性的优点,但需要进一步的生物化学方法的辅助和补充,对于试验结果的准确性有促进作用。
本试验通过形态学和生物化学方法,快速、准确地分析比较7个黑木耳菌株遗传差异性,但在新品种引选和种质资源评价中还需要进一步考察其栽培性状和产量等特性,以指导实际应用。进一步探明不同基因条带与对应特定性状的关系,比如菌丝活力强弱、耳片颜色深浅、耳脉多少等栽培特性,将是未来菌株遗传差异性分析的发展方向。
[1]郭兴,李滇华,任广明,等.ISSR分子标记鉴别黑龙江地区黑木耳主栽菌株的研究[J].中国林副特产,2010(6):14-16.
[2]唐利华,郭倩,王瑞娟,等.中国黑木耳主要栽培菌株酯酶同工酶的研究[J].食用菌学报,2007,14(4):37-40.
[3]张介驰,马庆芳,张丕奇,等.用RAPD分子标记鉴别黑木耳菌种的研究[J].菌物研究,2006,4(4):54-56.
[4]高娃,张丕奇,党阿丽,等.东北黑木耳野生菌株比较[J].食药用菌,2013,21(4):230-232.
[5]何彦,曾晓丽,汪思迪,等.黑木耳菌株酯酶同工酶酶谱多样性研究[J].生物技术,2009,19(6):8-10.
[6]刘春勇,张文成,任改新.苏芸金芽胞杆菌与蜡状芽胞杆菌基因组DNA同源性及多态性的研究[J].南开大学学报:自然科学版,1999,32(2):98-102,102.
Study on Genetic Polymorphism of 7 Auricularia auricula Strains
LU Xin-yan,LIU Kun,JIANG Jun,ZHENG Qiao-ping,SONG Xiao-ya
(Lishui Academy of Agricultural Sciences,Lishui 323000,China)
Genetic diversity of seven strains of Auricularia auricular from Heilongjiang province were analyzed by antagonistic reaction,esterase isoenzyme and DNA marker respectively.Both of the data from RAPDISSR and antagonistic effect indicated that seven strains could be devided into four distinct groups,such as the first group included Heiwei 9 and 981,the second group included Heiwei 10 and 8808,the third group included single strain Hei29,the 9809 and H916 were classified to the last group.Meanwhile,the antagonistic reaction showed that the three isolates with longer genetic distance of Hei 29,9809 and H916 also showed obvious antagonistic effect.The present study indicated that results obtained by the three methods were consistent,which could provide technical support for new species selection and germplasm resources evaluation of A.auricula.
Auricularia auricula;antagonistic effect;ester ase isoenzyme;molecular marker
S646.6
A
1003-8310(2017)01-0052-04
10.13629/j.cnki.53-1054.2017.01.012
现代农业产业技术体系建设专项(CARS-24);浙江省农业新品种选育重大科技专项(2016c02057-2)。
路新彦(1980-),女,硕士,助理研究员,主要从事食用菌栽培育种及菌种检测研究。E-mail:luxy1108@163.com
**通信作者:郑巧平(1968-),男,本科,高级农艺师,主要从事食(药)用菌栽培育种及废菌糠资源化利用。E-mail:lsnks@163.com
2016-11-20
