miR-30c调控PAI-1对血管内皮细胞活力和迁移的影响*
2017-01-03谭晓勇卢培林吴剑波
谭晓勇, 罗 茂, 卢培林, 吴剑波
(西南医科大学药物研究中心, 药学院心血管药理实验室, 四川 泸州 646000)
miR-30c调控PAI-1对血管内皮细胞活力和迁移的影响*
谭晓勇▲, 罗 茂▲, 卢培林, 吴剑波△
(西南医科大学药物研究中心, 药学院心血管药理实验室, 四川 泸州 646000)
目的: 深入研究微小RNA (miR)-30c靶向调控纤溶酶原激活物抑制物1(PAI-1)表达对血管内皮细胞活力和迁移能力的影响。方法:应用miR-30c模拟物(mimic)、抑制物(inhibitor)及阴性对照(NC)序列转染人脐静脉内皮细胞(HUVECs)后,RT-qPCR检测miR-30c水平及PAI-1的mRNA表达,Western blot法检测PAI-1蛋白的表达,CCK-8法和划痕实验分别检测细胞活力和迁移能力。生物信息学方法预测miR-30c与PAI-1的mRNA 3’-UTR结合位点,并用双萤光素酶报告基因验证miR-30c对PAI-1 mRNA的靶向作用。结果:miR-30c能够靶向调控PAI-1的mRNA和蛋白表达,与对照组和NC序列转染组比较,增加miR-30c表达可导致PAI-1的mRNA和蛋白表达降低,进而增强HUVECs的活力和迁移能力;相反,抑制miR-30c表达可导致PAI-1 的mRNA和蛋白表达升高,进而抑制HUVECs的活力和迁移能力。结论:高表达miR-30c可抑制PAI-1表达,导致HUVECs 活力和迁移能力明显增强,提示miR-30c可能参与调节内皮细胞功能。
微小RNA-30c; 人脐静脉内皮细胞; 纤溶酶原激活物抑制物1; 细胞活力; 细胞迁移
血管内皮细胞的激活、增殖、迁移是血管生成的关键[1-2]。研究显示,内皮细胞的损伤、激活和修复与部分心血管疾病如动脉粥样硬化、冠心病、高血压、糖尿病等的发生和发展密切相关[3]。微小RNA (microRNA, miRNA, miR)是一类内源性的、19~25个碱基长度的小分子非编码RNA,它可以通过与靶mRNA的3’-UTR互补导致其降解或抑制蛋白翻译,从而在人体生命活动中具有广泛的调控作用[4-5]。近来研究表明,部分miRNA广泛参与调节血管内皮细胞功能如增殖、迁移和凋亡等过程,参与血管生成的调节,与诸多心血管疾病的病理生理进程密切相关[6-7]。
已有研究表明,miR-30c与肿瘤发生发展关系密切[8],但miR-30c与心血管类疾病的研究尚少。其中,Duisters等[9]研究表明左心室肥厚病人和动物模型心肌组织中miR-30c表达下调,导致miR-30c靶标结缔组织生长因子(connective tissue growth factor,CTGF)表达升高,促进心室重构;Patel 等[10]研究发现人肺动脉内皮细胞的miR-30c能够直接靶向作用于组织型纤溶酶原激活物抑制物1(plasminogen activator inhibitor-1,PAI-1)表达,参与镰状细胞贫血,该过程受胎盘生长因子(placental growth factor,PIGF)诱导。PAI-1 是调节纤溶活性的重要因子,生理条件下是组织纤溶酶原激活物(tissue plasminogen activator,t-PA)和尿激酶型纤溶酶原激活物(urokinase-type plasminogen activator,u-PA)的特异性抑制物[11-13]。研究显示,PAI-1 能够通过影响细胞外基质的降解过程和细胞黏附功能从而参与调节血管生成[13]。综合提示,miR-30c可能通过靶向调控PAI-1表达,参与内皮细胞的增殖、迁移调节,进而调节内皮功能和血管生成。
材 料 和 方 法
1 材料
人脐静脉血管内皮细胞(human umbilical vein endothelial cells,HUVECs)购自ATCC;改良型RPMI-1640细胞培养基(HyClone);细胞转染试剂Lipofectamine 2000、TRIzol和DEPC水(Invitrogen);Mir-XTMmiRNA第1链合成试剂盒和SYBR®RT-qPCR试剂盒(Clontech);PrimeScriptTMRT reagent Kit with gDNA Eraser (Perfect Real Time)试剂盒(TaKaRa);E.colipoly (A) Polymerase和rATP (NEB);DNA Ladder Marker和2×Taq PCR Master Mix(北京天根生化科技公司);CCK-8试剂盒、胰蛋白酶、青霉素和链霉素(碧云天生物技术研究所);兔抗人β-actin、PAI-1单克隆Ⅰ抗和抗兔IgG Ⅱ抗(CST);双萤光素酶报告基因检测系统(Promega);其它试剂为进口产品或国产分析纯产品。高速冷冻离心机(Eppendorf);自动凝胶成像系统Gel DocXR+、蛋白电泳及转膜系统和CFX96 Touch 实时荧光定量PCR仪(Bio-Rad);ND-1000微量紫外可见分光光度计(NanoDrop);超净工作台(SW-CJ);-80 ℃超低温冰箱(Thermo)。吸头、离心管等经0.1% DEPC水浸泡24 h后高温灭菌,50 ℃烘干备用。PCR引物由Invitrogen公司合成,具体序列见表1。

表1 引物序列
F: forward; R: reverse.
2 方法
2.1 HUVECs的培养 使用含10%胎牛血清的改良型RPMI-1640细胞培养基(1×105U/L青霉素和100 mg/L链霉素)培养HUVECs,置于37 ℃、5% CO2、饱和湿度培养箱中常规培养。
2.2 miR-30c靶基因的预测 应用在线软件miRBase[14](http://www.mirbase.org/)获得miR-30c的基本信息,结合miRanda[15](http://www.microrna.org/)、TargetScan 6.2[16](http://www.targetscan. org/)和PicTar[17](http://www.pictar.org/)靶基因预测结果,分析其中交集结果并提交miTarbas[18](http://mirtarbase.mbc.nctu.edu.tw/)与已证实靶基因比对,结合已有文献报道[10],预测miR-30c与PAI-1 mRNA存在的靶向结合位点,用于后续实验分析。
2.3 miR-30c的转染 转染前24 h,取对数生长期HUVECs(1×105)铺于6孔板中,待生长至60%~70%汇合时,更换无血清培养基后同步进行miR-30c mimic (50 nmol/L)、miR-30c inhibitor (100 nmol/L)及对应阴性对照(negative control,NC)序列的转染。按照Lipofectamine 2000 转染试剂说明书进行,转染12 h 后更换新鲜培养基,24 h收集细胞提取总RNA,48 h收集细胞提取蛋白。
2.4 pMIR-REPORT萤光素酶报告基因检测 根据预测的靶基因结合位点设计并合成 miR-30c 与 PAI-1 mRNA 3’-UTR 结合的30~60 bp 寡核苷酸序列并插入双酶切位点,经酶切、PCR扩增鉴定、质粒抽提以及测序等步骤确定重组载体质粒pMIR-Luc-miR-30c-PAI-1 3’-UTR构建成功。进一步以构建的质粒为模板进行定点突变,构建突变型质粒。各步所用引物序列见表1。
将HEK 293细胞铺于48孔板,利用Lipofectamine 2000转染试剂,结合pMIR-Luc-miR-30c-PAI-1 3’-UTR重组质粒、对应的突变型质粒和miR-30c mimic (50 nmol/L)、miR-30c inhibitor (100 nmol/L)及其对应NC共转染HEK 293细胞72 h后,利用萤光素酶检测试剂盒(Promega)检测萤光素酶活性,验证作用靶点的真实性。
2.5 用RT-qPCR检测miR-30c的水平和靶标PAI-1的mRNA表达 TRIzol试剂盒提取各实验组的细胞总RNA,并分别应用Mir-XTMmiRNA 第1链合成试剂盒和SYBR®RT-qPCR 试剂盒和PrimeScriptTMRT reagent Kit with gDNA Eraser (Perfect Real Time)试剂盒制备cDNA模板。分别以U6 和 18S rRNA为内参照检测miR-30c的水平和靶标PAI-1的mRNA表达。应用SYBR Premix Ex Taq II (TaKaRa)进行qPCR检测。扩增条件为: 95 ℃ 10 min; 95 ℃ 10 s, 50 ℃~ 60 ℃ 20 s, 72 ℃ 30 s, 40个循环; 72 ℃ 3 min, 60 ℃~95 ℃绘制熔解曲线。应用2-ΔΔCt法计算miR-30c和靶标PAI-1 mRNA的表达量,引物见表1。
2.6 Western blot法检测PAI-1蛋白的表达 细胞转染48 h后加入适量预冷的RIPA buffer(含Roche蛋白酶抑制剂cocktail),超声裂解后12 000 r/min,4 ℃ 离心15 min,取上清。BCA法测定蛋白浓度后,样品蛋白:5×上样缓冲液比例为4∶1,100 ℃变性后,进行10% SDS-PAGE胶上样并转至PVDF膜。经封闭1 h后加入I抗4 ℃孵育过夜(PAI-1和内参照β-actin抗体的稀释比例均为1∶2 000)。TBST洗膜3次后,II抗孵育1 h,再次TBST漂洗3次后显影。
2.7 细胞活力和迁移能力的检测 HUVECs(1×104)接种于96孔板,待其基本融合时更换无血清培养基培养24 h后,每孔加入10 μL的CCK-8试剂37 ℃孵育1 h,全自动酶标仪测定450 nm波长处的吸光度(A),每组均取6复孔计算均值,统计学分析比较细胞活力的变化。HUVECs转染24 h后按1×105接种至6孔板,设置4个复孔,待细胞基本汇合后,在细胞表面划1条粗细均一的划痕,洗去悬浮的细胞并更换低血清培养基,于培养0 h、12 h和24 h时,倒置显微镜下观察拍照,使用ImageJ 软件分析迁移距离评估迁移能力的变化。
3 统计学处理
采用 SPSS 16.0 软件统计分析,所有实验数值以均数±标准差(mean±SD)表示,多组间比较采用单因素方差分析(one-way ANOVA)和方差齐性检验,两两比较用Bonferroni校正的t检验,以P<0.05为差异有统计学意义。
结 果
1 miR-30c对HUVECs活力和迁移能力的影响
应用划痕实验和CCK-8法分析HUVECs的迁移能力和细胞活力,结果显示与对照组和NC组比较,miR-30c mimic组的HUVECs迁移能力及细胞活力增高;反之,miR-30c inhibitor组的HUVECs迁移能力及细胞活力均下降(P<0.05)。上述结果说明过表达miR-30c可以显著促进内皮细胞的迁移和细胞活力;抑制miR-30c表达可明显抑制内皮细胞的迁移和细胞活力,见图1。
2 miR-30c靶基因PAI-1的预测及萤光素酶报告基因验证
在线软件预测、比对及文献分析结果显示,存在miR-30c与PAI-1 mRNA 3’-UTR区的靶向结合位点,可用于后续转录调控实验验证。根据预测的靶基因结合位点,我们成功构建pMIR-Luc-miR-30c-PAI-1 3’-UTR重组质粒及其突变型质粒。萤光素酶报告基因实验结果显示,与对照组和NC组比较,转染miR-30c mimic可显著抑制萤光素酶活性,转染miR-30c inhibitor抑制miR-30c后萤光素酶活性增强(P<0.05),而共转染突变型质粒组各组萤光素酶活性均无明显变化(数据未显示),提示miR-30c可直接结合于PAI-1 3’-UTR,靶向调控PAI-1的表达,见图2。

Figure 1.The effect of miR-30c on the migratory ability and viability of HUVECs. A: the representative images and quantitative analysis of the wound healing test (scale bar=1 mm;n=4); B: the result of CCK-8 assay (n=6). Mean±SD.*P<0.05vscontrol group;#P<0.05vsNC group.
图1 miR-30c对内皮细胞迁移能力和细胞活力的影响

Figure 2.Bioinformatic predictions and reporter gene assay of miR-30c targeting PAI-1. A: stem-and-loop structure of miR-30c; B: predictions of miR-30c binding to PAI-1; C: assessment of miR-30c binding to PAI-1 3’-UTR by reporter gene assays. Mean±SD.n=3.*P<0.05vscontrol group;#P<0.05vsNC group.
图2 miR-30c靶基因PAI-1的生物信息学预测及萤光素酶报告基因验证
3 miR-30c调控靶标PAI-1的mRNA表达
与对照组和NC组比较,过表达miR-30c能够显著抑制靶基因PAI-1 的mRNA表达;反之,抑制miR-30c表达可显著增强PAI-1的mRNA表达(P<0.05),见图3。

Figure 3.The expression levels of miR-30c (A) and its target PAI-1 mRNA (B) measured by RT-qPCR. Mean±SD.n=3.*P<0.05vscontrol group;#P<0.05vsNC group.
图3 miR-30c对靶标PAI-1 mRNA表达的影响
4 miR-30c调控靶标PAI-1蛋白表达
如图4所示,Western blot实验的结果表明,与对照组和NC组比较,过表达miR-30c能够明显抑制靶标PAI-1 蛋白的表达;相反,抑制miR-30c表达可明显增强PAI-1蛋白的表达(P<0.05)。
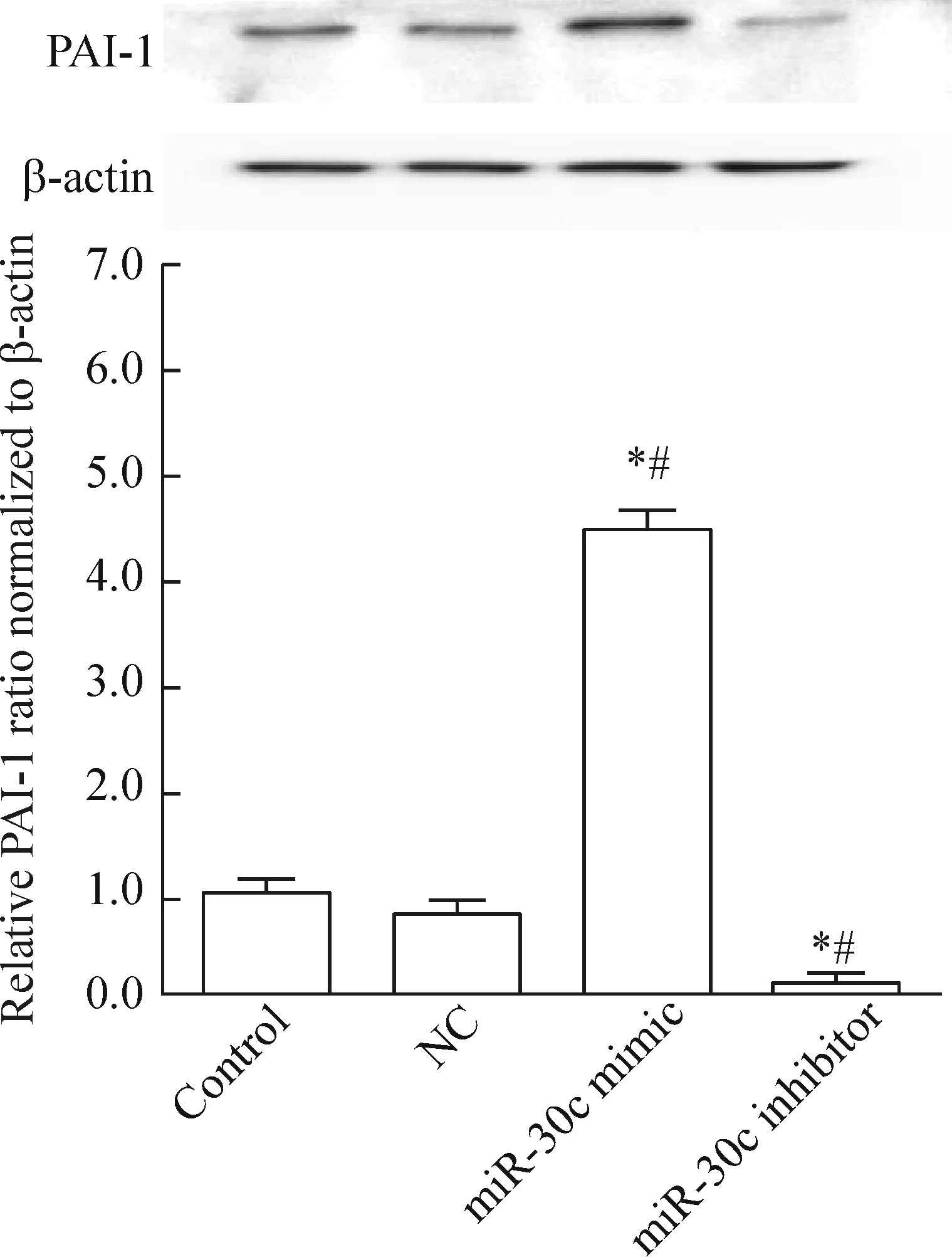
Figure 4.The effect of miR-30c on the protein levels of PAI-1. Mean±SD.n=3.*P<0.05vscontrol group;#P<0.05vsNC group.
图4 miR-30c对靶标PAI-1 蛋白表达的影响
讨 论
研究表明,血管生成过程中血管内皮细胞的功能激活是关键,血管内皮细胞的增殖、迁移等功能变化对维持血管壁稳定与循环系统稳态至关重要,与心血管系统疾病如动脉粥样硬化、冠心病、高血压等发生和发展密切相关。研究显示部分miRNA可以调节内皮细胞功能,参与内皮细胞的损伤、激活和修复等过程调节,进而影响内皮细胞介导的血管生成,在大量心血管疾病病理生理过程中发挥重要调控作用[6-7, 19]。例如,Bernstein 等[19]研究显示,大量miRNA通过Dicer通路加工形成,进而调控内皮毛细血管芽生和管腔形成,调控部分促血管生成的重要因子如血管内皮生长因子(vascular endothelial growth factor,VEGF)、血管生成素1/2(angiopoietin 1/2,Ang-1/2)等表达变化,进而调控血管的生成。
PAI-1是调节纤溶活性的重要因子,生理条件下是t-PA和u-PA的特异性抑制物[11-13]。研究显示,PAI-1能够通过影响细胞外基质的降解过程和细胞黏附功能从而参与调节血管生成[11]。进一步推测其机制,PAI-1活性升高,可抑制纤溶,导致细胞外基质(extracellular matrix,ECM)降解障碍;高水平PAI-1也通过阻止玻连蛋白(vitronectin)的精氨酸-甘氨酸-天冬氨酸(Arg-Gly-Asp,RGD)亲合力功能区从而破坏细胞的黏附和迁移功能。其中,玻连蛋白是细胞外基质中的主要黏附成分[12]。我们最近发现PAI-1与玻连蛋白结合可通过αvβ3整合素通路阻止VEGF诱导的VEGFR-2磷酸化,进一步研究显示通过调节纤维蛋白溶酶、尿激酶受体和αvβ3整合素受体,PAI-1 能够抑制VEGF诱导的血管形成[13]。
已有研究表明,miR-30c与脂肪干细胞分化、上皮细胞间质转化和肿瘤等相关[20-21]。最近研究揭示,miR-30c在心血管系统中同样扮演着重要角色[9-10]。例如,miR-30c通过靶向调控结缔组织生长因子表达,参与心室重构过程的调节[9];HPMVEC细胞内miR-30c经PIGF诱导后靶向调控PAI-1表达,参与镰状细胞贫血过程的调节[10];本文研究结果显示,miR-30c能够靶向结合PAI-1 3’-UTR种子序列,从而直接靶向负调控PAI-1的mRNA和蛋白表达,参与内皮细胞的迁移、细胞活力的调节,提示过表达miR-30c可抑制PAI-1表达,导致HUVECs 细胞活力和迁移能力的明显增强,推测miR-30c可能通过直接靶向调控PAI-1表达,影响细胞外基质的降解或黏附过程,进而参与血管生成过程。
Wu等[13]研究表明2 型糖尿病中应用PAI-1 抑制剂可明显修复缺血诱导的血管生成,增加下肢的血流灌注,提示PAI-1 可作为糖尿病微血管并发症血管生成治疗的一个潜在重要靶点,临床研发如miR-30c增强剂等新型PAI-1抑制剂将可能改善心血管类血管并发症的治疗及预后,如糖尿病下肢缺血血管生成治疗过程,增加血管重构,阻止血管渗漏,达到最理想治疗效果。本研究结果为进一步设计新型PAI-1抑制剂提供一定依据,其结果具有潜在的临床应用价值。
[1] Friis T, Engel AM, Bendiksen CD, et al. Influence of levamisole and other angiogenesis inhibitors on angiogenesis and endothelial cell morphologyinvitro[J]. Cancers (Basel), 2013, 5(3):762-785.
[2] Ju R, Cirone P, Lin S, et al. Activation of the planar cell polarity formin DAAM1 leads to inhibition of endothelial cell proliferation, migration, and angiogenesis[J]. Proc Natl Acad Sci U S A, 2010, 107(15): 6906-6911.
[3] 李乾豪, 仉红刚. 细胞微泡 miRNA 对内皮细胞的调控[J]. 中国病理生理杂志, 2015, 31(8):1531-1536.
[4] Gao Y, Peng J, Ren Z, et al. Functional regulatory roles of microRNAs in atherosclerosis[J]. Clin Chim Acta, 2016, 460:164-171.
[5] Samanta S, Balasubramanian S, Rajasingh S, et al. MicroRNA: a new therapeutic strategy for cardiovascular di-seases[J].Trends Cardiovasc Med, 2016, 26(5):407-419.
[6] 张文宇, 王 辉, 李玉媚, 等. miRNA-24对内皮型一氧化氮合酶表达调节及血管内皮细胞增殖的影响[J]. 中国病理生理杂志, 2014, 30(8):1415-1420.
[7] 郑志伟, 劳海燕, 余细勇, 等. 冠心病患者血浆循环miR-126的表达及其对血管内皮细胞的影响[J]. 中国病理生理杂志, 2011, 27(12):2313-2317.
[8] Irani S, Hussain MM. Role of microRNA-30c in lipid metabolism, adipogenesis, cardiac remodeling and cancer[J]. Curr Opin Lipidol, 2015, 26(2):139-146.
[9] Duisters RF, Tijsen AJ, Schroen B, et al. miR-133 and miR-30 regulate connective tissue growth factor: implications for a role of microRNAs in myocardial matrix remodeling[J]. Circ Res, 2009, 104(2):170-178.
[10]Patel N, Tahara SM, Malik P, et al. Involvement of miR-30c and miR-301a in immediate induction of plasminogen activator inhibitor-1 by placental growth factor in human pulmonary endothelial cells[J]. Biochem J, 2011, 434(3):473-482.
[11]Yasar Yildiz S, Kuru P, Toksoy Oner E, et al. Functional stability of plasminogen activator inhibitor-1[J]. Sci World J, 2014, 2014:858293.
[12]Czekay RP, Loskutoff DJ. Plasminogen activator inhibitors regulate cell adhesion through a uPAR-dependent mechanism[J]. J Cell Physiol, 2009, 220(5):655-663.
[13]Wu J, Strawn TL, Luo M, et al. Plasminogen activator inhibitor-1 inhibits angiogenic signaling by uncoupling vascular endothelial growth factor receptor-2-αVβ3 integrin cross talk[J]. Arterioscler Thromb Vasc Biol, 2015, 35(1):111-120.
[14]Griffiths-Jones S, Grocock RJ, van Dongen S, et al. miRBase: microRNA sequences, targets and gene nomenclature[J]. Nucleic Acids Res, 2006, 34(Database issue):D140-D144.
[15]John B, Enright AJ, Aravin A, et al. Human microRNA targets[J]. PLoS Biol, 2004, 2(11):e363.
[16]Lewis BP, Burge CB, Bartel DP, et al. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets[J]. Cell, 2005, 120(1):15-20.
[17]Chen K, Rajewsky N. Natural selection on human micro-RNA binding sites inferred from SNP data[J]. Nat Genet, 2006, 38(12):1452-1456.
[18]Chou CH, Chang NW, Shrestha S, et al. miRTarBase 2016: updates to the experimentally validated miRNA-target interactions database[J]. Nucleic Acids Res, 2016, 44(D1):D239-D247.
[19]Bernstein E, Kim SY, Carmell MA, et al. Dicer is essential for mouse development[J]. Nat Genet, 2003, 35(3):215-217.
[20]Irani S, Hussain MM. Role of microRNA-30c in lipid metabolism, adipogenesis, cardiac remodeling and cancer[J]. Curr Opin Lipidol, 2015, 26(2):139-146.
[21]Wen M, Men R, Liu X, et al. Involvement of miR-30c in hepatic stellate cell activation through the repression of plasminogen activator inhibitor-1[J]. Life Sci, 2016, 155:21-28.
(责任编辑: 卢 萍, 罗 森)
Effects of miR-30c on viability and migratory ability of HUVECs by targeting
PAI-1TAN Xiao-yong, LUO Mao, LU Pei-lin, WU Jian-bo
(DrugDiscoveryResearchCenter,LaboratoryforCardiovascularPharmacology,TheSchoolofPharmacy,SouthwestMedicalUniversity,Luzhou646000,China.E-mail:jbwucn@163.com)
AIM: To investigate the effect of microRNA (miR)-30c on the viability and migratory ability of human umbilical vein endothelial cells (HUVECs) by targeting plasminogen activator inhibitor-1 (PAI-1).METHODS: The HUVECs were transfected with miR-30c mimic and inhibitor or negative control (NC), and then the expression levels of miR-30c, PAI-1 mRNA and protein were detected by RT-qPCR and Western blot. The viability and migratory ability of HUVECs were measured by CCK-8 assay and wound healing test. After bioinformatic analysis, the assessment of miR-30c binding to PAI-1 3’-UTR was carried out using a luciferase reporter gene assay. RESULTS: miR-30c directly down-regulated PAI-1 levels by binding to the 3’ UTR seed sequence of PAI-1 mRNA. Furthermore, transfection of a miR-30c mimic down-regulated the expression of PAI-1 at mRNA and protein levels, leading to enhanced migratory ability and viability of the HUVECs. However, transfection of a miR-30c inhibitor up-regulated the expression of PAI-1 at mRNA and protein le-vels, leading to decreased migratory ability and viability. CONCLUSION: Regulation of miR-30c level changes the migratory ability and viability of HUVECs by affecting the PAI-1 expression, indicating the involvement of miR-30c in modulating endothelial function.
MicroRNA-30c; Human umbilical vein endothelial cells; Plasminogen activator inhibitor-1; Cell viability; Cell migration
1000- 4718(2016)12- 2199- 06
2016- 07- 15
2016- 08- 30
国家自然科学基金资助项目(No. 81172050; No. 81570263);四川省科技厅课题(No. 2014FZ0104);四川省教育厅课题(No.16ZA0178)
R363
A
10.3969/j.issn.1000- 4718.2016.12.012
杂志网址: http://www.cjpp.net
△通讯作者 Tel: 0830-3161673; E-mail: jbwucn@163.com
▲并列第1作者
