獐子岛近岸人工鱼礁区浮游动物群落结构特征
2016-12-19张皓宇刘汉超刘永虎
张皓宇,刘汉超,刘永虎,田 涛,陈 勇,杨 军
( 大连海洋大学,辽宁省海洋牧场工程技术研究中心,辽宁 大连 116023 )
獐子岛近岸人工鱼礁区浮游动物群落结构特征
张皓宇,刘汉超,刘永虎,田 涛,陈 勇,杨 军
( 大连海洋大学,辽宁省海洋牧场工程技术研究中心,辽宁 大连 116023 )
为研究獐子岛近岸人工鱼礁区浮游动物群落结构特征,利用2014年6月、10月和2015年6月、10月獐子岛近岸人工鱼礁区4个航次的调查资料,对该礁区浮游动物的物种种类、密度、生物量、多样性指数、与环境因子的关系等方面进行分析。调查结果表明,共发现29种浮游动物(包括6类浮游幼虫),2014年6月16种,2014年10月12种,2015年6月28种,2015年10月24种;优势种9种(包括浮游幼虫4种),拟长腹剑水蚤、中华哲水蚤、桡足类六肢幼体为4个航次共同优势种;浮游动物生物量和密度分布情况基本一致,最高值出现在2015年6月份的站位2,节肢动物门生物量和密度占绝对优势;聚类分析中2014年6月、10月和2015年6月浮游动物群落均分为4个聚类组,2015年10月分为3个聚类组;水温和盐度是影响浮游动物群落关系十分重要的环境因子,与浮游动物密度和生物量呈显著相关关系。
浮游动物群落;生物量;密度;聚类分析;相关分析
浮游动物是海洋生态系统的重要组成部分,作为次级生产者,在物质循环和能量流动过程中有着非常重要的地位,起着承上启下的作用[1-4]。一方面,浮游动物与浮游植物之间的摄食关系可以适当地调控浮游植物群落的密度和生物量变化;另一方面,浮游动物为海域中的鱼虾蟹以及其他游泳动物提供重要食物来源,在海洋生态系统的食物网中占据重要位置[5-8]。浮游动物群落结构会随着周围水域环境的变化而发生变化,能够及时对水质的优劣状态做出反应,浮游动物的多样性是水域生态系统服务功能的重要指标之一[9-11]。因此对于浮游动物的调查研究具有非常重要的现实意义,有利于海洋生态系统的修复和完善。
建设人工鱼礁区是改善海洋生态环境的重要途径,礁体能够为海洋生物提供摄食、产卵繁殖、生长、避难、休养生息的场所。人工鱼礁能够通过改变周围海域的非生物环境因子来影响生物环境[12]。投礁后的海流形态会发生很大变化,礁体的迎流方向形成上升流,背流方向形成涡流,不同流态使海域中营养盐不断地交替补充,非常有利于浮游生物群落的生长,由于摄食关系最终能够诱集更多的海洋生物,使人工鱼礁区生物资源量不断扩大[13-14]。
獐子岛海域位于黄海北部,长山群岛南部,海岸线曲折绵延,渔业资源丰富,拥有全国最大的虾夷扇贝(Patinopectenyessoensis)养殖基地[15]。在獐子岛近岸海域投放了大量海珍品增殖礁,人工鱼礁对改善海域生态环境,维护海洋生物资源起至关重要作用[16]。国内对于獐子岛海域相关的调查和研究已有诸多报道,包括水体中环境因子的研究,生态环境的评估,鱼类资源、底栖藻类、大型底栖动物的调查等方面[15-18],但对于獐子岛海域特别是围绕其人工鱼礁区浮游动物方面的相关研究却很少,仅见尹洁慧[19]在2009—2010年对獐子岛海域的浮游动物进行了周年调查。
本文通过对獐子岛近岸人工鱼礁区浮游动物的调查研究,分析浮游动物的群落结构,以期为海洋牧场的合理建设,渔业资源的可持续发展提供基础资料和理论依据。
1 材料与方法
1.1 采样地点与站位设置
于2014年6、10月和2015年6、10月在獐子岛近岸人工鱼礁区进行了4次调查,共设置8个站位(图1)。
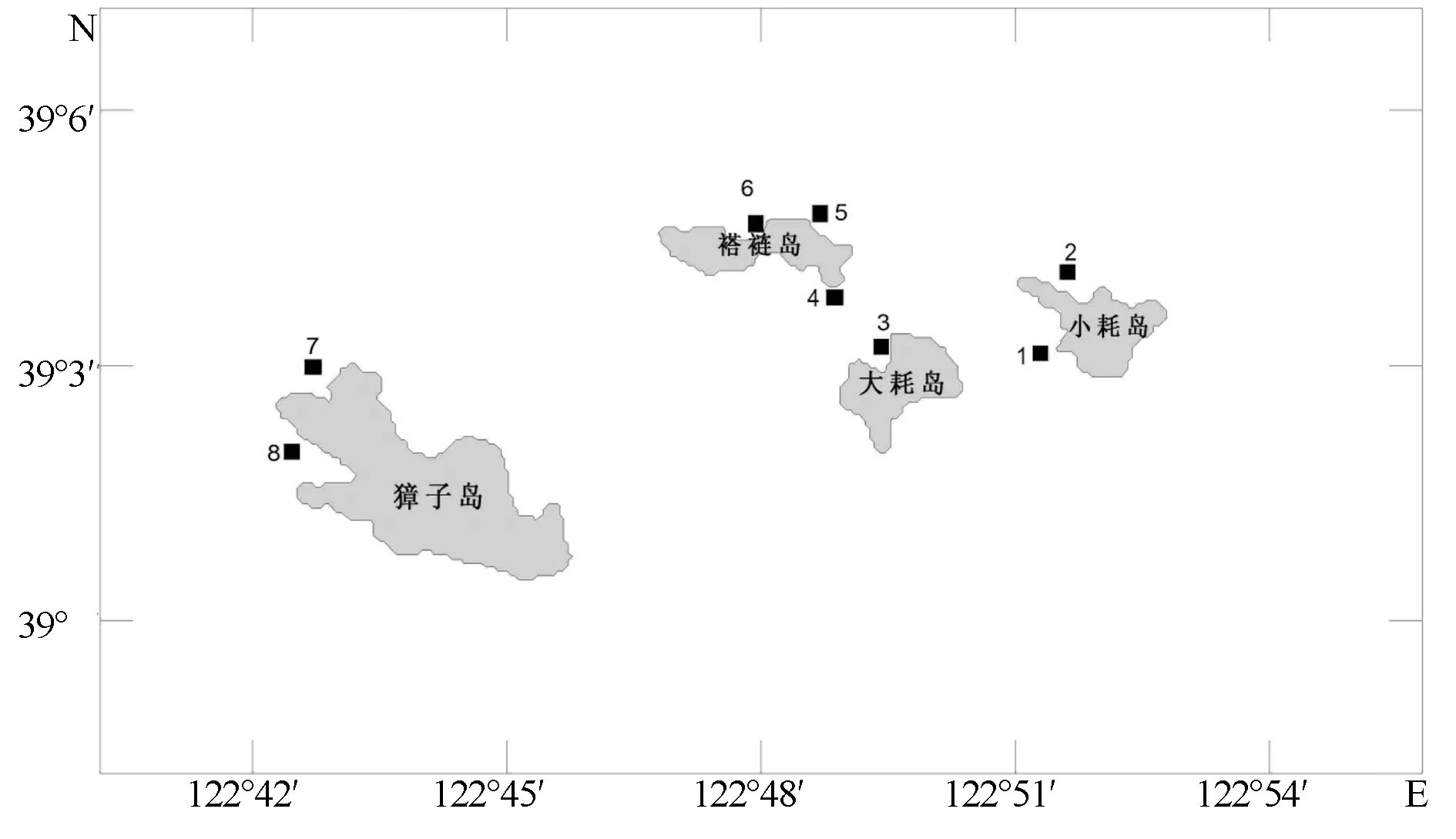
图1 调查站位分布
1.2 样品采集与分析
在4个航次的浮游动物调查取样过程中,使用浅水Ⅲ型浮游生物网由底层至表层进行垂直拖网,样品放入样品瓶中,用5%甲醛溶液进行保存。在实验室内利用显微镜进行镜检分析,计数和鉴定浮游动物种类。具体方法参照《国家海洋调查规范 第6部分:海洋生物调查》[20]进行。
1.3 数据处理
浮游动物的多样性用Shannon-Weaver多样性指数(H′)表示,群落的均匀度用Pielou均匀度指数(J′)表示,群落的丰富度用物种丰富度指数(D)表示,群落的优势种类用优势度(Y)表示,各指数计算公式如下[21-23]:
H′=-∑PilnPi
J′=H′/lnS
D=(S-1)/lnN
Y=ni/Nfi
式中,Pi为i种种类的数量比例,S为附着动物的种类数量;N为附着动物的总个数;f为某种类出现频率。
运用SPSS 19.0和Excel 2010软件进行相关数据分析。
2 结果与分析
2.1 种类组成
4个航次调查共发现浮游动物6大类29种,其中,节肢动物13种,占总物种数的44.83%,刺胞动物2种,占总物种数的6.90%,原生动物3种,占10.34%,毛额动物和脊索动物各1种,分别占3.45%,浮游幼虫8类,占27.29%。其中,2014年6月份共16种,2014年10月份12种,2015年6月份共28种,2015年10月份24种。
4个航次调查中均发现的有拟长腹剑水蚤(Oithonasimilis)、小拟哲水蚤(Paracalanusparvus)、中华哲水蚤(Calanussinicus)、短角长腹剑水蚤(O.brevicornis)、克氏纺锤水蚤(Acartiaclause)、强壮箭虫(Sagittacrassa)、桡足类六肢幼体、短尾类蚤状幼虫、双壳类幼体、多毛类幼体、长腕幼虫(表1)。
4个航次调查中节肢动物、毛颚动物和浮游幼虫均有出现,其中节肢动物种数量在4次调查中均占优势,仅在2014年10月少于浮游幼虫(少1种),2015年6月出现最多(14种),2014年10月出现最少(5种);毛额动物4个航次调查均出现1种;浮游幼虫物种数变化不明显,2014年6月出现最少(5种),2015年10月出现最多(8种);原生动物只出现在2015年的2个航次调查中,且物种数无明显变化;脊索动物出现在2014年6月和2015年的2个航次调查中,均为1种;刺胞动物只在2015年6月出现2种(图2)。
综上所述,2015年2个航次调查发现的浮游动物种类数(29种)高于2014年调查所发现的浮游动物种类数(18种),年际变化比较明显。同一年的6月调查比10月调查发现的种类数要多,但相差种类数不多,说明同一年份的种类的两季节差异不明显。
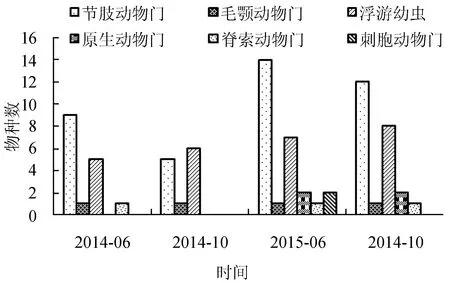
图2 浮游动物不同类群的季节分布
2.2 浮游动物的生物量与密度分布情况
浮游动物生物量和密度分布情况见图3。4个航次獐子岛近岸人工鱼礁区海域浮游动物生物量之间存在一定差异(F=0.9465,P=0.4332)。生物量依次为2015年6月>2015年10月>2014年6月>2014年10月。节肢动物门在生物量上占有绝对优势,生物量为0.2396~1.4220 g/m3,最高生物量出现在2015年6月;浮游幼虫生物量次之,为0.0160~0.1881 g/m3,最高生物量出现在2015年6月;毛颚动物门生物量为0.0018~1.0171 g/m3,最高生物量出现在2014年6月;脊索动物门生物量为0.0010~0.0015 g/m3。
浮游动物密度季节之间存在一定差异(F=0.9782,P=0.4269)。浮游动物密度分布情况和生物量分布情况基本一致(图4)。
综合4个航次浮游动物的生物量和密度数据,节肢动物门所占比例最高,刺胞动物门最低。
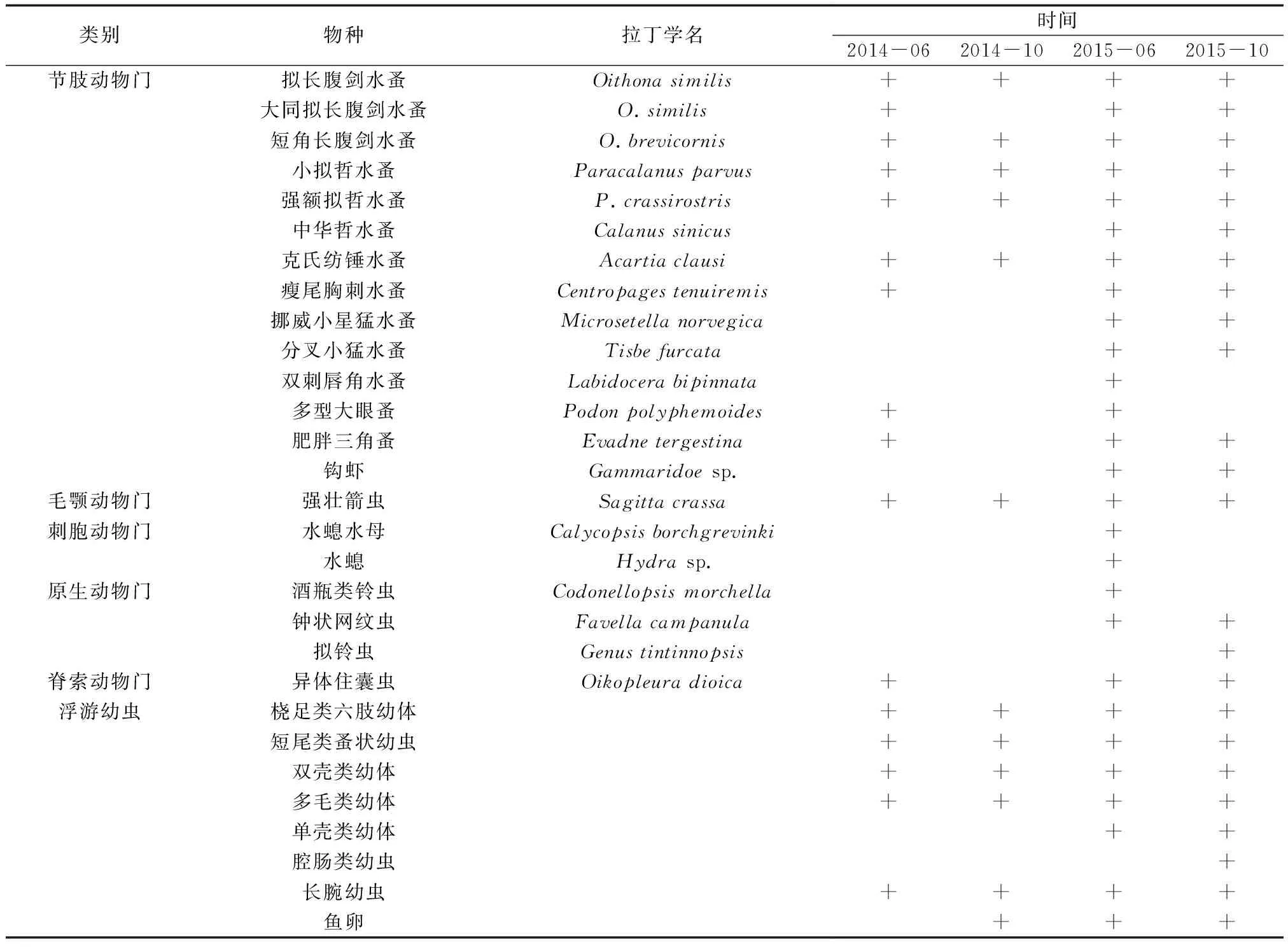
表1 獐子岛人工鱼礁区浮游动物的种类组成
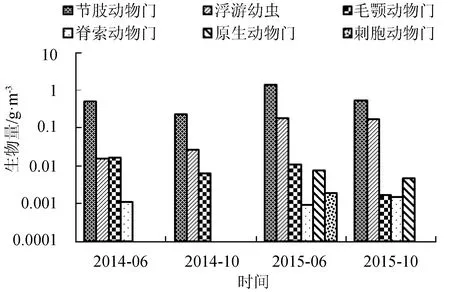
图3 各门类浮游动物生物量的分布情况

图4 各门类浮游动物密度的分布情况
浮游动物2014年6月平均生物量为0.0689 g/m3。生物量最高点出现在站位5(0.1048 g/m3),次高点出现在站位1(0.0958 g/m3),即褡裢岛人工鱼礁区北部海域和小耗岛人工鱼礁区西部海域,生物量由这两处海域逐渐向四周降低,生物量最低点出现在站位7(0.0423 g/m3),位于獐子岛西北部海域(图5)。2014年10月平均生物量为0.0338 g/m3,生物量最高点出现在站位5(0.0661 g/m3),高值区出现在褡裢岛北部海域,生物量最低点出现在站位1(0.0036 g/m3),生物量由北部向南部逐渐降低。2015年6月平均生物量0.2041 g/m3,生物量最高点出现在站位2(0.5149 g/m3),位于小耗岛人工鱼礁区北部海域,生物量最低点为站位7(0.0969 g/m3),位于獐子岛人工鱼礁区的西北部海域。2015年10月平均生物量为0.0949 g/m3,生物量最高点出现在站位4(0.2006 g/m3),位于褡裢岛人工鱼礁区北部海域,生物量最低点为站位7(0.0261 g/m3),低值区主要为调查海域的东南部海域。
浮游动物2014年6月平均密度为7232.196个/m3,2014年10月平均密度为3618.111个/m3,2015年6月平均密度25358.5820个/m3,2015年10月平均密度为9974.539个/m3。 浮游动物密度高低值分布情况与生物量分布情况基本一致(图6)。
4个航次调查中,浮游动物生物量和密度高值区主要分布在褡裢岛和小耗岛人工鱼礁区附近海域,低值区主要分布在獐子岛人工鱼礁区附近。
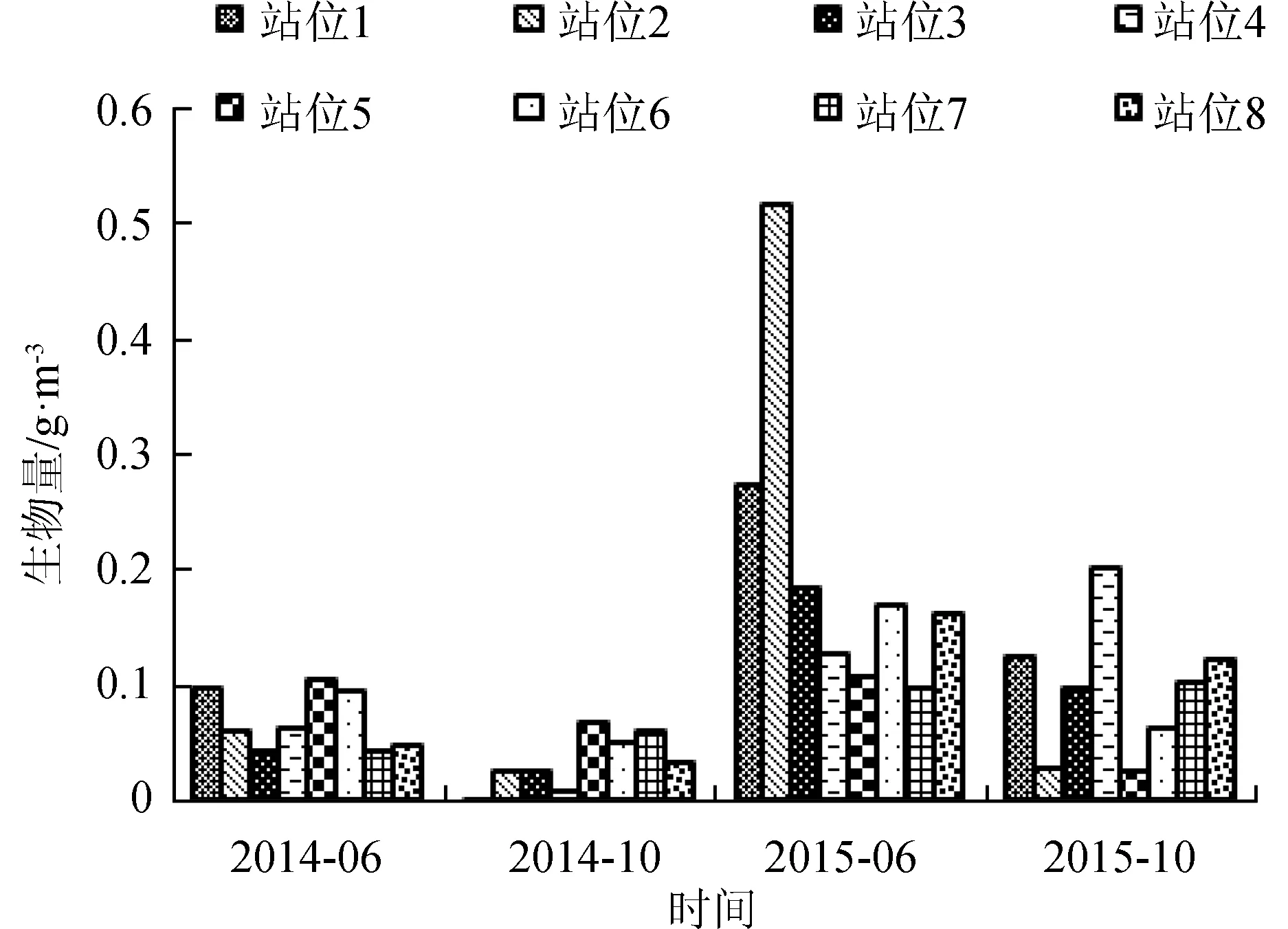
图5 浮游动物生物量的水平分布情况

图6 浮游动物密度的水平分布情况
2.3 浮游动物的优势种及优势度
獐子岛近岸人工鱼礁区4个航次共发现优势种9种,其中包括桡足类5种,浮游幼虫4种。2014年发现优势种7种,2015年发现优势种8种;拟长腹剑水蚤、中华哲水蚤、桡足类六肢幼体为4个航次共同优势种,其中2015年6月的拟长腹剑水蚤优势度最高(0.6798);克氏纺锤水蚤、长腕幼虫、双壳类幼体均为季节单一优势种,其余6种优势种在两个季节均有出现(表2)。
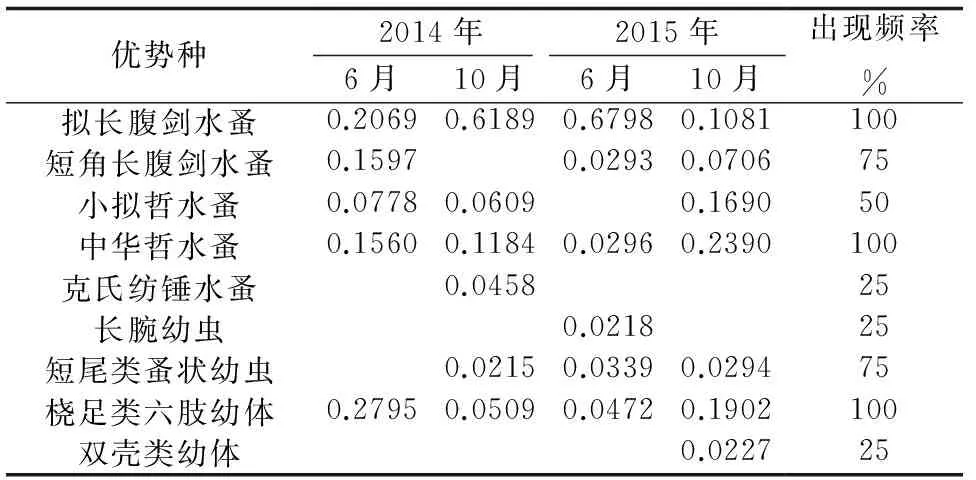
表2 浮游动物优势种及其优势度
2.4 浮游动物的生物多样性
浮游动物的Shannon-Weaver多样性指数最高值出现在2015年10月(2.2263),最低值出现在2014年10月(1.3993),在4个航次调查中,2015年10月的浮游动物的群落多样性和群落的复杂程度最好;Pielou均匀度指数最高值出现在2014年6月(0.7170),最低值出现在2015年6月(0.4465),2014年6月的种类数虽然在4个航次中不是最多,但其物种在群落结构上分布的均匀度最高;丰富度指数最高值出现在2015年6月(2.240),最低值出现在2014年10月(1.0893),2015年6月浮游动物种类数在4个航次中最多(28种),其种类最为丰富(表3)。
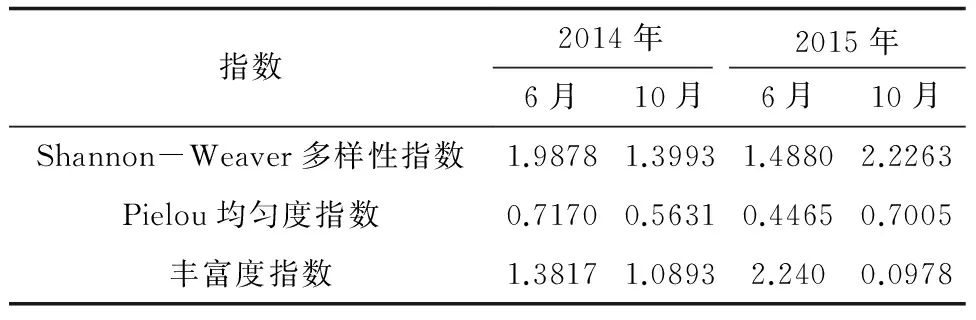
表3 浮游动物群落的物种多样性指数
2.5 浮游动物的聚类分析
通过聚类分析可知(图7):
(1) 2014年6月的浮游动物群落分为4个聚类组:物种种类、密度和生物量较少的站位8和密度和生物量较高的站位6各单独一类;站位2和站位7组成一个聚类组,这两个站位生物量和密度也比较少,是拟长腹剑水蚤的主要聚集区之一;其余站位为一聚类组。
(2) 2014年10月的浮游动物群落分为4个聚类组:中华哲水蚤密度最高的站位7,物种数量多、双壳类幼体聚集区的站位2,以及密度最低、物种数量最少的站位1,3个站位分别各自为一个聚类组;其余站位为一聚类组。
(3) 2015年6月的浮游动物群落分为4个聚类组:小拟哲水蚤主要聚集区的站位8,拟长腹剑水蚤主要聚集区域的站位2各自为一个聚类组;物种数量较少的站位6和站位7组成一个聚类组;其余站位为一个聚类组。
(4) 2015年10月的浮游动物群落分为3个聚类组:浮游动物密度和生物量较低的站位2、站位5和站位6一个聚类组;站位1和站位4为一个聚类组,两个站位密度和生物量均较高,也是浮游幼虫主要聚集地;其余站位为一聚类组。

图7 浮游动物群落聚类分析
2.6 浮游动物群落与环境因子的关系
通过对浮游动物的密度和生物量与各环境因子的关系进行分析可知(表4、表5),2014年6月密度和生物量与水温均为极显著正相关,相关系数分别为0.905、0.907;密度和生物量与盐度均显著负相关,相关系数分别为-0.764、-0.751。2014年10月份密度和生物量与盐度均极显著负相关,相关系数分别为-0.873、-0.871。2015年6月密度与水温显著正相关,相关系数为0.707;密度和生物量与盐度均显著负相关,相关系数分别为-0.739、-0.741。2015年10月密度与盐度极显著负相关,相关系数为-0.909;生物量与盐度显著负相关,相关系数为-0.899。由此看出水温和盐度是影响浮游动物群落结构的重要环境因子。

表4 浮游动物密度与环境因子的相关分析
注:*表示0.01<α≤0.5,显著相关(双侧检验);**表示α≤0.01,极显著相关(双侧检验).下同.

表5 浮游动物生物量与环境因子的相关分析
3 讨 论
3.1 浮游动物群落的密度、生物量和优势种
4个航次调查共发现浮游动物29种(包括8种浮游幼虫),其中2014年18种,2015年29种,包括节肢动物、毛颚动物、刺胞动物、脊索动物、原生动物、浮游幼虫6大类。尹洁慧[19]2009—2010年在獐子岛海域对浮游动物进行调查,共发现浮游动物36种(包括浮游幼虫7种),涉及到节肢动物、毛颚动物、刺胞动物、脊索动物、原生动物、栉水母动物6大类(包括浮游幼虫)。由于2009—2010年调查为逐月调查,调查海域为养殖区和非养殖区,调查面积大,站位多,而本次调查主要针对夏秋季节的浮游动物群落,相较于2009—2010年的调查,本次调查物种数量相对较少。在2009—2010年优势种包括中华哲水蚤、双刺唇角水蚤、鸟喙尖头蚤(Penetiaavirostris)、细长脚(Themistogracilipes)、克氏纺锤水蚤、长腕类幼虫、夜光虫(Noctilucascintillans)、强壮箭虫,共8种,本次调查中,中华哲水蚤、克氏纺锤水蚤、长腕类幼虫仍保持优势种地位,其余优势种已被拟长腹剑水蚤、短角长腹剑水蚤、小拟哲水蚤、短尾类蚤状幼虫、桡足类六肢幼体、双壳类幼体取代。优势种群在浮游动物群落中占据最大比例,控制着浮游动物密度和生物量的大小,而生物量/密度比值越大,浮游动物个体越大,比值越小,浮游动物个体越小[24],本次调查中的9种优势种中包括4类浮游幼虫,其余优势种主要为中型浮游动物[25],浮游动物个体比较小,这与此次调查浮游动物密度高,生物量相对较低的结果相对应。
依据2009—2010年逐月多样性数据对比,其中各月份丰富度指数最高值为1.56,多样性指数最高值为1.81,均匀度指数最高值为0.67,本次调查航次中丰富度指数最高值为2.24,多样性指数最高值为2.23,均匀度指数最高值为0.72;2009—2010年逐月调查中浮游幼虫7种,优势种8种,本次调查中浮游幼虫增至8种,优势种增至9种。浮游动物各物种多样性指数、浮游幼虫和优势物种数均有不同程度的增加。本次调查主要围绕人工鱼礁区进行,人工鱼礁对浮游动物的诱集作用可能是重要原因,促进了浮游动物的增殖和结构优化。但具体原因有待进一步深入研究分析。
4个航次调查发现的浮游动物主要生态类型为广温广盐型,代表种类有拟长腹剑水蚤、小拟哲水蚤;近岸低盐型,代表种类有克氏纺锤水蚤;低温高盐型,代表种类有中华哲水蚤。这也是优势种群的主要生态类型。这是因为该浮游动物群落主要生态类型的形成与周围海域形成的环流有关,獐子岛近岸受到北黄海海域沿岸流和暖流所组成的气旋式环流的影响,形成黄海冷水团和沿岸水团两大基本水团,而在这两大水团的影响下的浮游动物以低温高盐性和广温广盐性为主[10]。
3.2 浮游动物群落与环境因子的关系
浮游动物群落结构与周围环境因子有非常重要的关系。水温的变化影响浮游动物的种类、生物量和密度的变化,进而影响到浮游动物群落的变化[26]。4个航次调查中6月份密度和生物量均高于10月,6月至10月水温趋势变化是由上升到最高水温之后逐渐下降,浮游动物与水温的相关性分析显示,6月浮游动物密度和生物量与水温呈显著正相关;2014年10月和2015年10月浮游动物密度与水温之间的相关系数分别为0.559、0.628,生物量与水温之间的相关系数分别为0.559、0.597,说明水温是影响浮游动物群落结构的重要参数。这与尹洁慧[19]对獐子岛与环境因子的关系的分析结果一致。4个航次调查中发现的浮游动物群落物种生态类型中包括近岸低盐类,且存在优势种(克氏纺锤水蚤),这可能是造成与盐度呈负相关关系的重要原因[27]。2014年6月份叶绿素与浮游动物密度和生物量呈正相关,其他3个航次调查呈负相关,说明在合适的范围内,叶绿素含量的增加会促进浮游动物群落的生长,但若含量过高又会对浮游动物起到抑制作用;营养盐对浮游动物的影响也是如此,一方面适当的营养盐为浮游动物提供养分,对浮游动物起积极作用,另一方面过高的营养盐含量会破坏群落结构[28]。
[1] 徐晓群,曾江宁,寿鹿,等.兴化湾浮游动物群落季节变化和水平分布[J].生态学报,2010,30(3):734-744.
[2] 孙军,刘东艳,王宗灵,等. 浮游动物摄食在赤潮生消过程中的作用[J].生态学报,2004, 24(7):1514-1522.
[3] 孙松,李超伦,张光涛,等. 胶州湾浮游动物群落长期变化[J]. 海洋与湖沼,2011, 42(5):625-631.
[4] 吴利,冯伟松,张堂林,等. 湖北省西凉湖浮游动物群落周年动态变化及其与环境因子的关系[J]. 湖泊科学,2011,23(4):619-625.
[5] 郭沛涌,沈焕庭,刘阿成,等. 长江河口浮游动物的种类组成、群落结构及多样性[J].生态学报,2003, 23(5):892-900.
[6] Newton G M. Estuarine ichthyoplankton ecology in relation to hydrology and zooplankton dynamics in a salt-wedge estuary[J]. Marine and Freshwater Research,1996, 47(2):99-111.
[7] Merrell J R,Stoecker D K. Differential grazing on protozoan microplankton by developmental stages of the calanoid copepodEurytemoraaffinisPoppe[J]. Journal of Plankton Research, 1998, 20(2):289-304.
[8] Strom S L,Brainard M A, Holmes J L, et al. Phytoplankton blooms are strongly impacted by microzooplankton grazing in coastal North Pacific waters[J]. Marine Biology, 2001, 138(2):355-368.
[9] 纪焕红,叶属峰,刘星,等. 南麂列岛海洋自然保护区浮游动物的物种组成及其多样性[J].生物多样性,2006,14(3):206-215.
[10] 杨青,王真良,樊景凤,等. 北黄海秋、冬季浮游动物多样性及年间变化[J]. 生态学报,2012, 32(21):6747-6754.
[11] 姜作发,唐富江,董崇智,等. 黑龙江水系主要江河浮游植物种群结构特征[J]. 吉林农业大学学报,2007, 29(1):53-57.
[12] 田丽斯,李莹,张明,等. 獐子岛潮间带底栖海藻资源及其季节性变化[J].水产科学,2009, 28(3):142-145.
[13] 盛化香. 崂山湾人工鱼礁区底栖大型海藻群落的季节性和区域性变化[D].青岛:中国海洋大学,2012.
[14] 陈涛,王云龙,廖勇. 象山港人工鱼礁区浮游动物种类组成及群落结构特征[J].海洋科学,2014(7):41-46.
[15] 王全超,韩庆喜,李宝泉. 辽宁獐子岛马牙滩潮间带及近岸海区大型底栖动物群落特征[J]. 生物多样性,2013,21(1):11-18.
[16] 田涛,陈勇,陈辰,等. 獐子岛海洋牧场海域人工鱼礁区投礁前的生态环境调查与评估[J]. 大连海洋大学学报,2014, 29(1):75-81.
[17] 段丽琴,宋金明,袁华茂,等. 獐子岛附近海域沉积物中氮和磷的分布、来源及污染风险评价[J].广西科学院学报,2015,31(3):173-178.
[18] 陈勇,杨军,田涛,等. 獐子岛海洋牧场人工鱼礁区鱼类资源养护效果的初步研究[J]. 大连海洋大学学报,2014,29(2):183-187.
[19] 尹洁慧. 獐子岛海域浮游动物群落时空变化特征及其与环境因子之间的关系[D].青岛:中国科学院大学,2013.
[20] 国家海洋局. GB/T 12763.6—2007,海洋调查规范第6部分:海洋生物调查[S].北京:中国标准出版社,2007.
[21] Pielou E C. An introduction to mathematical ecology[J]. Bioscience, 1969, 78(1):7-12.
[22] Wilhm J L.Use of Biomass Units in Shannon′s Formula[J]. Ecology, 1968, 49(1):153-156.
[23] 吴建新,李强,张晴. 吕泗大洋港浮游植物群落与环境因子关系[J].生态学杂志,2013, 32(2):396-400.
[24] 方良,李纯厚,杜飞雁,等.大亚湾海域浮游动物生态特征[J].生态学报,2010, 30(11):2981-2991.
[25] 林青, 由文辉, 徐凤洁,等. 滴水湖浮游动物群落结构及其与环境因子的关系[J].生态学报, 2014, 34(23):6918-6929.
[26] 陈光荣,钟萍,张修峰,等. 惠州西湖浮游动物及其与水质的关系[J].湖泊科学,2008, 20(3):351-356.
[27] 马玉艳,韩华,卞正和,等.大小长山岛周围海域浮游动物群落分布特征[J].海洋环境科学,2012,31(3):364-369.
[28] 张武昌, 张翠霞, 肖天. 海洋浮游生态系统中小型浮游动物的生态功能[J].地球科学进展,2009,24(11):1195-1201.
DistributionCharacteristicsofZooplanktonCommunityinArtificialReefAreainZhangziIsland
ZHANG Haoyu, LIU Hanchao, LIU Yonghu, TIAN Tao, CHEN Yong, YANG Jun
( Center for Marine Ranching Engineering Science Research of Liaoning, Dalian Ocean University, Dalian 116023,China )
The zooplankton species composition,dominant species,biomass, abundance distribution,and species diversity were studied in artificial reef area of Zhangzi Island in Dalian based on the data surveyed in June and Octorber of 2014 and 2015. The results showed that 29 zooplankton species were identified during the survey,most being offshore lower-salinity group,wide temperature salinity group and offshore lower-temperature group. In total,16 species were observed in June of 2014,12 species in October of 2014, 28 species in June of 2015,and 24 species in October of 2015.There were 9 dominant species, almost the Arthropod, and the main dominant species wereCalanussinicus, Copepodite larvae, andOithonasimiles. The distribution of biomass and abundance of zooplankton were consistent basically, and correlated with both water temperature and salinity. Community cluster analysis indicated that in October and June of 2014 and in June of 2015, the zooplankton was divided into 4 cluster groups, and 3 cluster groups in October of 2015.
zooplankton community; biomass; abundance; cluster analysis; correlation analysis
10.16378/j.cnki.1003-1111.2016.05.003
S932.2
A
1003-1111(2016)05-0473-07
2015-12-13;
2016-03-22.
国家科技支撑计划项目(2012BAD18B03,2013BAD23B01).
张皓宇(1988—),女,硕士研究生;研究方向:浮游动物及大型藻类附着生物.E-mail:709103847@qq.com. 通讯作者:刘永虎(1984—),男,实验师,硕士;研究方向:海洋牧场.E-mail:t695@163.com.
