低氧胁迫对西伯利亚鲟幼鱼生理状态的影响
2016-12-19王晓雯胡红霞马国庆
王晓雯,朱 华,胡红霞,马国庆
( 北京市水产科学研究所,渔业生物技术北京市重点实验室,北京 100068 )
低氧胁迫对西伯利亚鲟幼鱼生理状态的影响
王晓雯,朱 华,胡红霞,马国庆
( 北京市水产科学研究所,渔业生物技术北京市重点实验室,北京 100068 )
为探究低氧胁迫对西伯利亚鲟幼鱼的血液基础指标、抗氧化和能量代谢相关酶活力的影响,以体质量为(19.46±4.9) g的西伯利亚鲟幼鱼为研究对象,开展了低氧(2.3 mg/L)胁迫3 h和自高氧(18.5 mg/L)逐渐降至低氧(2.8 mg/L)又恢复到正常溶氧(7 mg/L),这两种不同情况下的低氧胁迫试验。检测了血液红细胞数、血红蛋白质量浓度,以及肝脏和肌肉的抗氧化酶活力和鳃组织三磷酸腺苷酶活力。结果显示,低氧胁迫下西伯利亚鲟幼鱼血红细胞数升高,血红蛋白质量浓度、血液pH、肝脏和肌肉的超氧化物歧化酶和过氧化氢酶活力以及碱性磷酸酶活力均低于正常溶氧组,而鳃组织钠/钾—三磷酸腺苷酶活力无明显变化。试验结果显示,急性低氧胁迫能较快使西伯利亚鲟幼鱼血液红细胞数目增加,并影响肝脏、肌肉的抗氧化应激及代谢相关酶活力,但是短时间内西伯利亚鲟幼鱼可能还未实现鳃部的能量调节。
西伯利亚鲟;血液指标;抗氧化酶;三磷酸腺苷酶;碱性磷酸酶
溶解氧是水产养殖的一个重要环境因子,直接关系到鱼类的生存、生长和代谢水平[1-3]。溶解氧水平降低会导致鱼类摄食减少,生长缓慢,影响其免疫机能,甚至引起死亡[4-7]。急性缺氧和溶解氧剧烈变化作为鱼类生活中不可避免的短期胁迫,对其生存产生重要影响。因此,鱼类通过对生存环境的长期适应,获得了很多耐受低氧的机制,如鱼体可以提高呼吸频率[8]、降低摄食量、增强血氧亲和力[9]和提高血液循环率[10]来适应低氧环境。
水体中溶解氧水平过低或过高均可导致鱼类发生强烈的应激反应。鱼体则利用自身酶类抗氧化剂清除多余的活性氧簇,包括超氧化物歧化酶、过氧化氢酶及谷胱甘肽过氧化物酶等。有研究发现,缺氧及溶解氧含量的恢复过程中会导致变温动物抗氧化酶活力的变化[11-13],低氧条件下鱼体进行无氧呼吸,产生的副产物可能会抑制过氧化氢酶活力[14],导致鱼体抗氧化机制相应发生变化[15]。
鱼体在对环境进行适应的过程中,需要进行包括神经调节、膜流动性调节以及能量代谢调节等方面[16]的调节。钠/钾—三磷酸腺苷酶在离子转运、能量代谢、物质运输、营养吸收[17]以及信息传递方面发挥重要作用。王春枝等[18]在对鲢鱼(Hypophthalmichthysmolitrix)组织急性低氧胁迫过程中发现低氧促进三磷酸腺苷酶活性升高,但当低氧超过自身调节范围,三磷酸腺苷酶活性呈现下降趋势。并有研究显示,多种应激条件下机体三磷酸腺苷酶活性均发生改变,对应激条件比较敏感。但尚未见鲟鱼的相关报道。
西伯利亚鲟(Acipenserbaerii),属鲟科、鲟属,介于软骨鱼类和硬骨鱼类之间的软骨硬鳞鱼类,是我国引自欧洲的鲟鱼种类,现已成为我国常见的鲟鱼养殖品种,具有较高的经济价值和研究价值。野生西伯利亚鲟生活在河流中,为半洄游性种群,耐低氧能力较弱。本研究以西伯利亚鲟幼鱼为对象,探讨其在急性低氧和由高溶解氧逐渐降低再恢复这两种情况下鱼体血液生化指标、抗氧化酶活力和能量代谢相关酶活力的变化情况,以丰富基础研究数据,为鲟鱼养殖生产提供理论依据。
1 材料与方法
1.1 试验鱼的来源与驯化
试验用西伯利亚鲟幼鱼购自北京市怀柔,于北京市水产科学研究所实验室自净化循环水槽中暂养7 d,水槽容量为直径1 m,高0.8 m,实际水容积500 L,水温(19.0±1.0) ℃,充气以保证溶解氧≥6 mg/L,日投饵2次。从暂养7 d的西伯利亚鲟鱼中挑选活力好、健康的个体进行试验。
1.2 试验方案
将挑选体质量为(19.46±4.9) g的试验鱼60尾随机平分为溶解氧质量浓度(7.8±0.3)、(2.3±0.2) mg/L这两个试验组,每组3个平行;低氧组通过充入氮气使溶解氧迅速地在1 h内降至(2.3±0.2) mg/L,并通过少量充入空气保持低氧处理3 h。设置溶解氧质量浓度(7.8±0.3) mg/L为对照组,连续充入空气,维持溶解氧质量浓度。
各试验平行组随机选取3尾鱼,尾静脉采血,置于含肝素钠的离心管中,混匀。采血后解剖,取出肝脏、肌肉和鳃组织用液氮速冻后置于-80 ℃冰箱保存备用。
1.2.2 溶解氧变化试验
于9:00,挑选体质量(15.9±4.7) g的试验鱼60尾平均放入3个试验水槽,连续充液氧1 h,溶解氧升至19 mg/L,停止充氧,并充入氮气使溶解氧缓慢降至4 mg/L后迅速降至2 mg/L,随后停止充氮气,继续充液氧,在溶解氧水平为18.5 mg/L、7.5 mg/L、4.2 mg/L、2.8 mg/L以及恢复至7 mg/L时刻各平行组随机取3尾鱼采血取样,整个溶解氧变化过程在8 h内完成。取样时具体操作同1.2.1。
1.3 血液基础指标的测定
取50 μL采集的抗凝血液,使用HANNA台式酸度测定仪进行pH的测定。
取150 μL 采集的抗凝血液,使用商品试剂盒测定血红蛋白质量浓度(微量游离血红蛋白测定试剂盒,南京建成生物工程研究所),单位为g/L。
取10 μL抗凝血,用0.9%生理盐水稀释150倍,使用25×16型血球计数板对红细胞进行计数。
式中,σy为地基中某点水平面上的垂直压力,假定等于该处单位面积上部土柱的重量为与σy相应的垂直有效压力,假定等于该处单位面积上部土柱的有效重量(浸润线以下采用土的浮容重);αmax为地面最大地震加速度(9度地震采用0.4 g,8度地震采用0.2 g,7度地震采用 0.1 g);(△τf/)cr为根据土层三轴震动试验资料求得发生液化时的震动剪应力比,取cr为40次。
红细胞数/个·L-1=80小格内细胞总数/80×400×107×稀释倍数
红细胞血红蛋白平均含量/g·个-1=血红蛋白质量浓度(g/L)/红细胞数(个/L)[19]
1.4 西伯利亚鲟肝脏和肌肉相关酶活力测定
总超氧化物歧化酶的测定采用黄嘌呤氧化酶法,其活力定义为每毫克组织蛋白在1 mL反应液中超氧化物歧化酶抑制率达50%时所对应的超氧化物歧化酶活力单位(U)。
过氧化氢酶活定义为每毫克组织蛋白每秒钟分解1 μmol的H2O2的量作为一个活力单位(U)。
碱性磷酸酶活力定义为1分钟内每克组织蛋白在37 ℃将1 μmol底物转化为酚的量为1个活力单位(U)。
钠/钾—三磷酸腺苷酶的活力单位定义为每小时每毫克蛋白的组织中三磷酸腺苷酶分解三磷酸腺苷产生1 μmol无机磷的量作为1个酶活力单位(U)。
肝和肌肉总超氧岐化酶活力、过氧化氢酶活力,碱性磷酸酶活力、鳃钠/钾—三磷酸腺苷酶活力均使用试剂盒(南京建成生物工程研究所),按照相关说明书进行,采取可见光法,用UV/Vis-4802S型双光束紫外可见分光光度计进行检测。
1.4 数据处理
利用EXCEL和SPSS 13.0软件对数据进行处理,所得数据用平均值±标准差表示,急性低氧试验结果进行t检验;溶解氧剧烈变化试验结果采用单因素方差分析进行组间显著性分析,将不同溶解氧含量下的数据进行Duncan多重比较,以P<0.05作为差异显著的标准。
2 结 果
2.1 低溶解氧对鲟鱼血液pH的影响
经过3 h的急性低氧处理,测得溶解氧为(2.3±0.2) mg/L处理组鲟鱼血液的pH为5.26±0.23,显著低于溶解氧为(7.8±0.3) mg/L对照组的pH 6.4±0.36(P<0.05)。
溶解氧由18.5 mg/L降至4.2 mg/L过程中,鲟鱼血液pH略微上升,而从4.2 mg/L继续降至2.8 mg/L的过程中,鲟鱼血液pH下降(图1)。
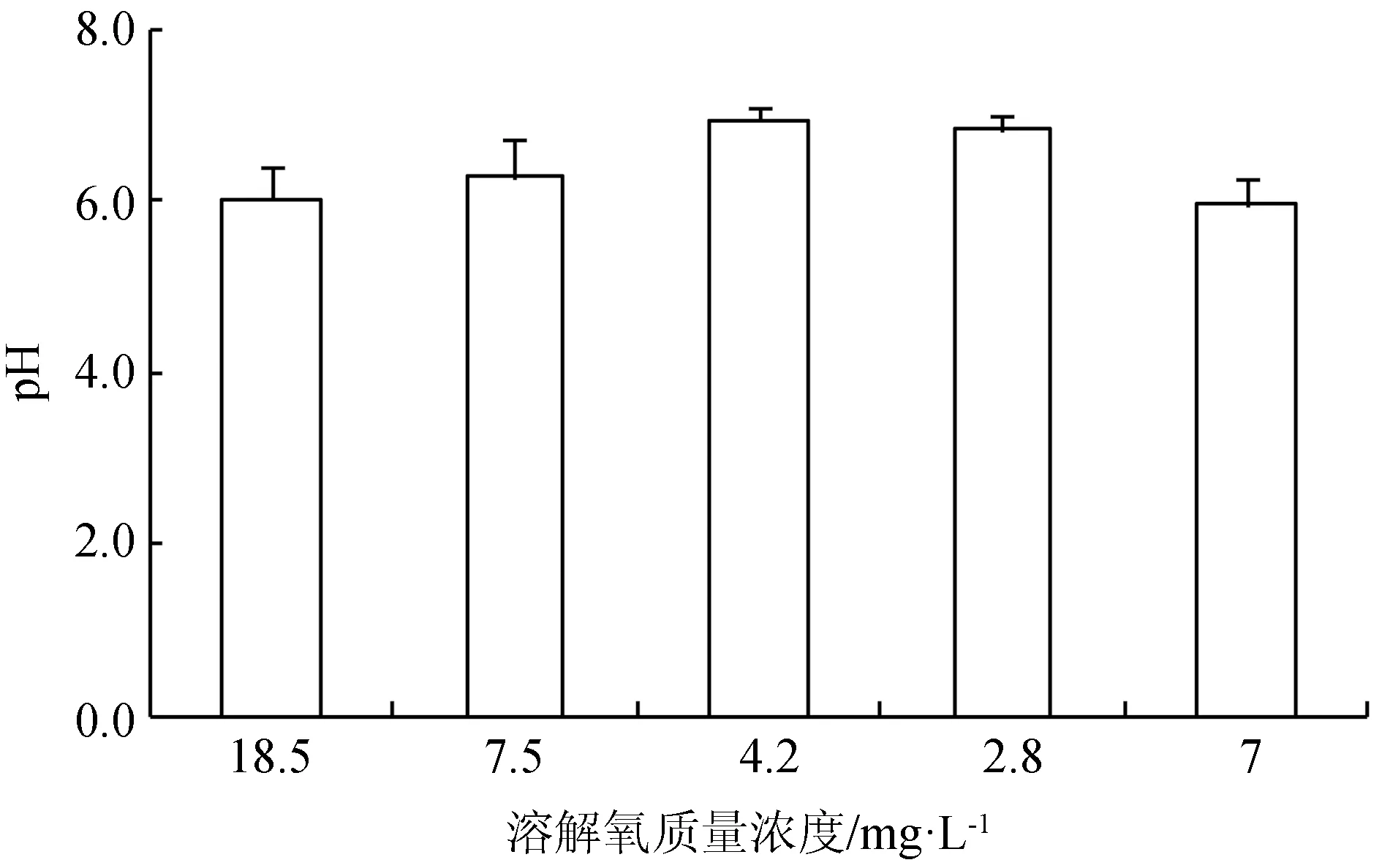
图1 不同溶氧条件下西伯利亚鲟幼鱼血液的pH
2.2 低溶解氧对鲟鱼血细胞数和血红蛋白质量浓度的影响
急性低氧处理3 h后,处理组红细胞数目显著高于对照组(P<0.05),而血红蛋白质量浓度低于对照组(P<0.05),红细胞血红蛋白平均含量显著低于对照组(P<0.05)。其中红细胞数高于对照组10.6%,血红蛋白质量浓度低于对照组31.3%(表1)。
在水体溶解氧质量浓度逐渐降低(由18.5 mg/L降至正常水平7.5 mg/L)的过程中,红细胞数目减少,血红蛋白质量浓度和红细胞血红蛋白平均含量无明显变化;当溶解氧继续降至4.2 mg/L 时,血细胞数目出现显著升高(P<0.05),溶解氧迅速降至2.8 mg/L时,血细胞数目发生略微的减少,并且血红蛋白质量浓度减小,此时鱼体内可能产生无氧代谢,溶解氧再次恢复到7 mg/L时,血细胞数目得到恢复(图2~图4)。
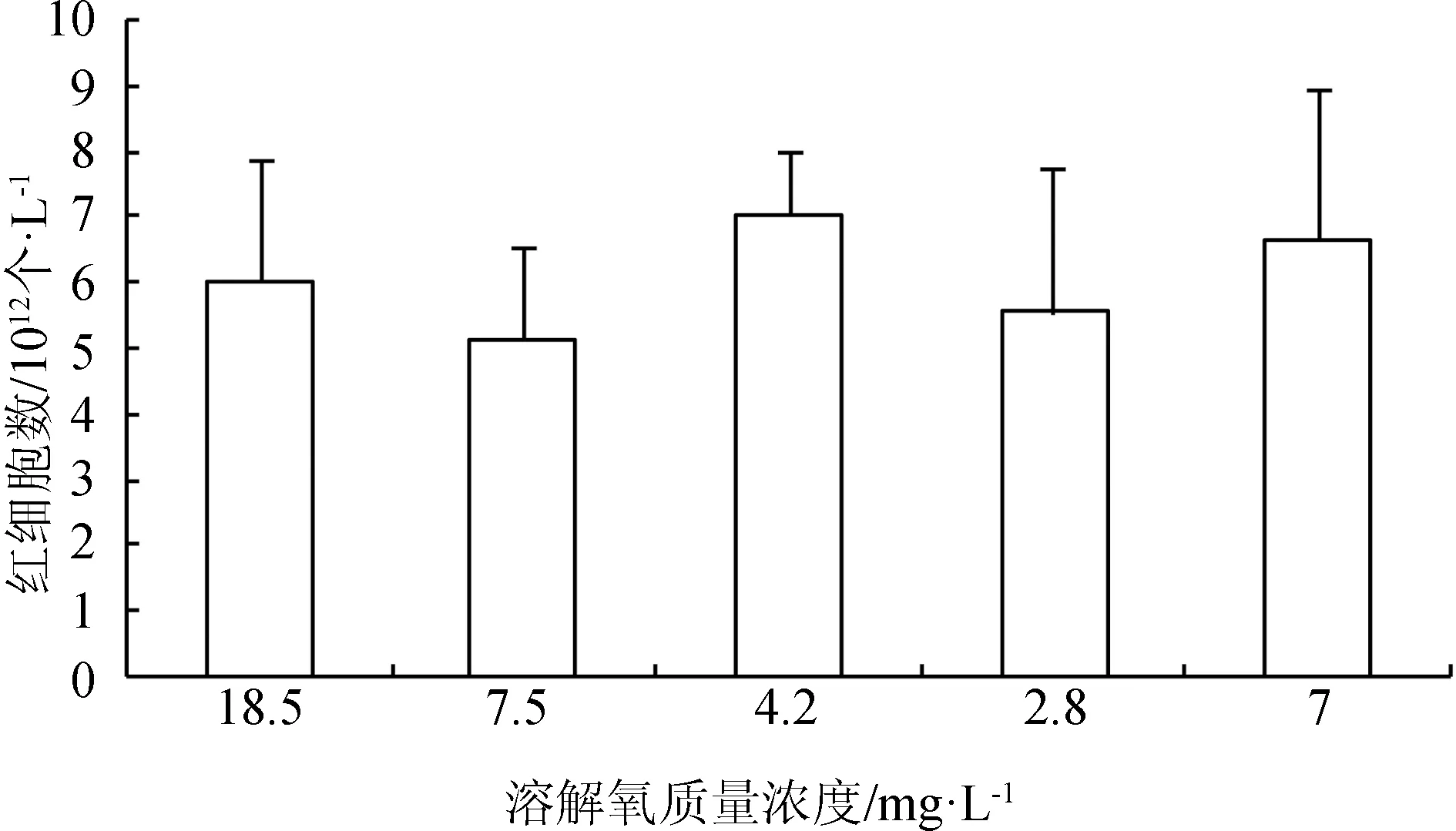
图2 不同溶解氧水平下西伯利亚鲟幼鱼血液红细胞数
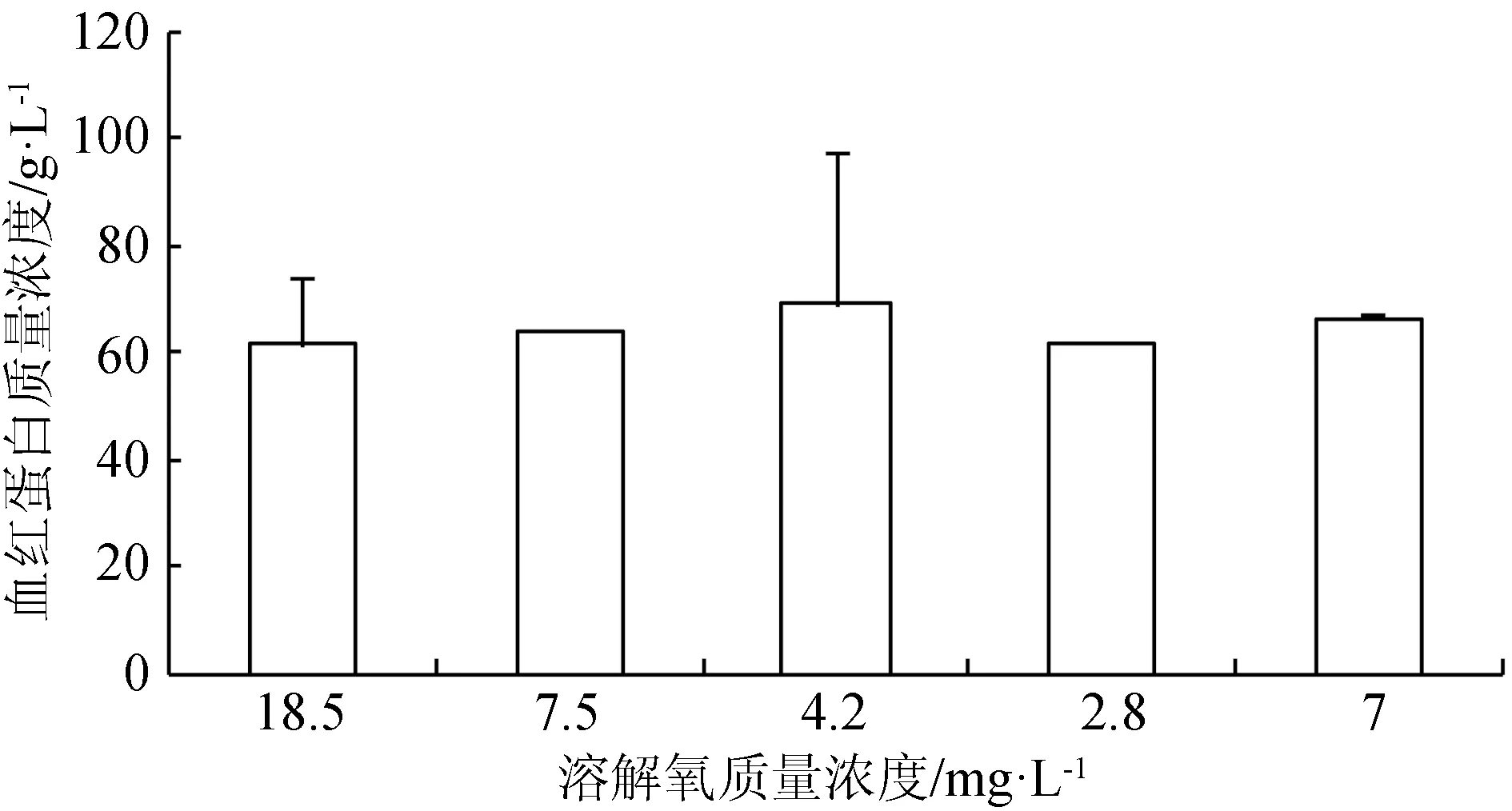
图3 不同溶解氧水平下西伯利亚鲟幼鱼血液血红蛋白质量浓度
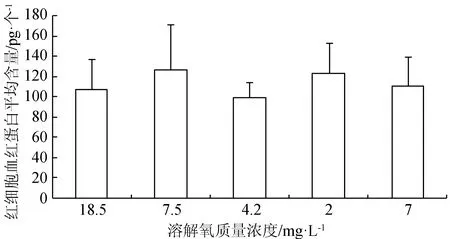
图4 不同溶解氧水平下西伯利亚鲟幼鱼血液红细胞血红蛋白平均含量
2.3 低溶解氧对鲟鱼抗氧化酶活力的影响
在急性低氧胁迫3 h后,低氧处理组肝脏和肌肉的超氧化物歧化酶活力,过氧化氢酶活力均显著低于正常溶解氧组(P<0.05)(表2)。
在溶解氧变化试验中,西伯利亚鲟幼鱼肝脏和肌肉组织的超氧化物歧化酶活力随着溶解氧质量浓度降低,其活力降低,待溶解氧质量浓度恢复到7 mg/L时,超氧化物歧化酶活力明显升高(图5)。肝脏过氧化氢酶的变化规律与超氧化物歧化酶一致。随着溶解氧质量浓度由18.5 mg/L降至7.5 mg/L时,肌肉的过氧化氢酶活力骤然升高,溶解氧质量浓度逐步降至2.8 mg/L的过程中,肌肉过氧化氢酶活力逐渐下降,当溶解氧质量浓度由2.8 mg/L恢复到7 mg/L时,肌肉过氧化氢酶活力显著升高(P<0.05)(图6)。

表1 急性低氧下西伯利亚鲟幼鱼红细胞数、血红蛋白质量浓度和红细胞血红蛋白平均含量
注:同一列上标字母不同的数据间差异显著(P<0.05).

表2 急性低氧试验中鲟鱼肝脏和肌肉的抗氧化酶和碱性磷酸酶活力
注:同一列上标注字母相同的数据间差异显著(P<0.05).

图5 不同溶解氧条件下西伯利亚鲟幼鱼肝脏和肌肉超氧化物歧化酶活力注:图中不同大写字母代表不同溶解氧质量浓度的数据具有显著性差异(P<0.05),以下各图同.
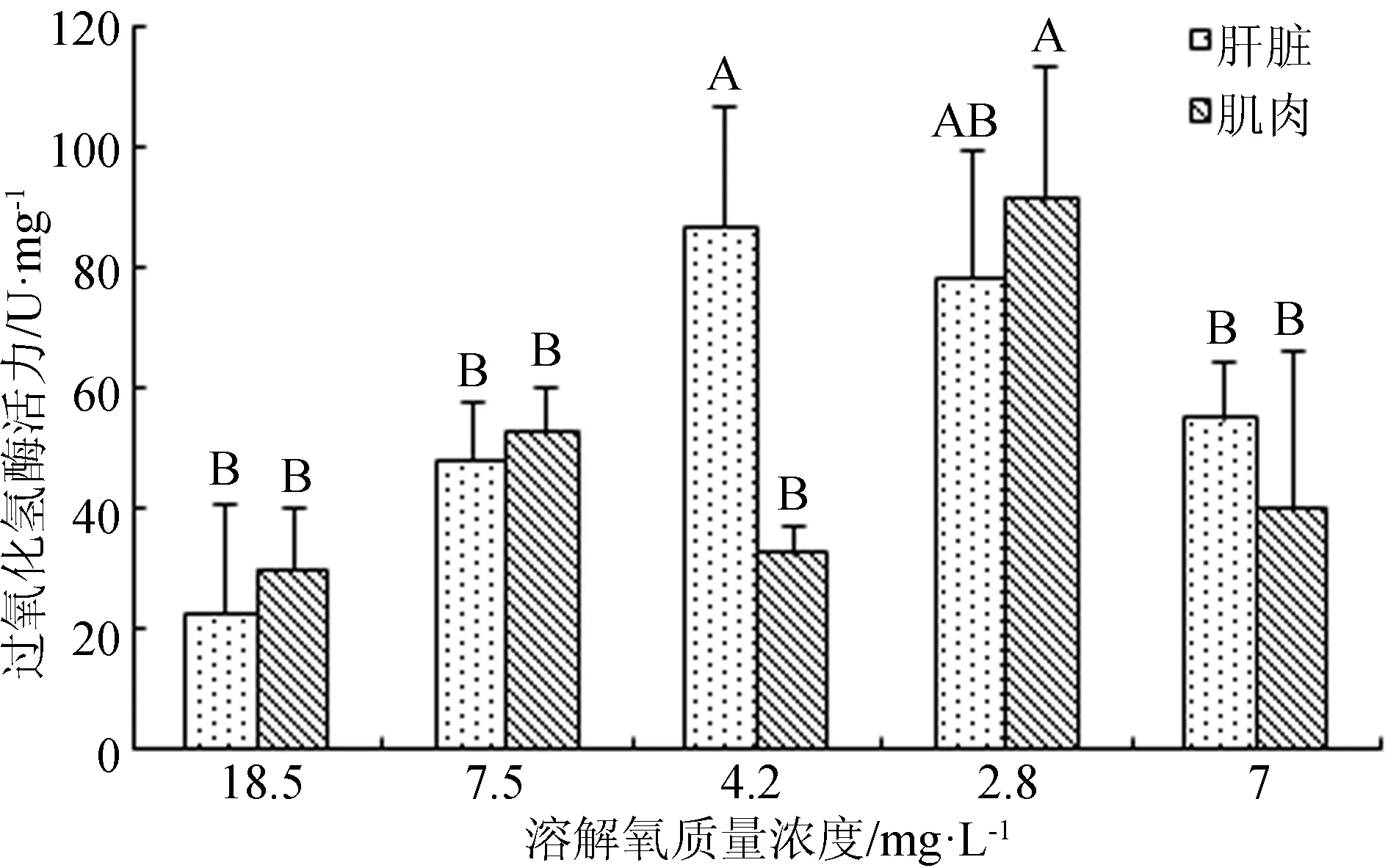
图6 不同溶解氧条件下西伯利亚鲟幼鱼肝脏和肌肉过氧化氢酶活力
2.4 低溶解氧对鲟鱼能量、代谢相关酶活力的影响
急性低氧处理3 h,西伯利亚鲟幼鱼鳃组织的三磷酸腺苷酶活力为(2.01±0.14) U/mg,对照组鳃部三磷酸腺苷酶活力为(2.78±1.07) U/mg,差异不显著(P>0.05)。低氧处理组肝脏的碱性磷酸酶活力显著低于正常溶氧组(P<0.05),而肌肉的碱性磷酸酶活力无显著差异(P>0.05)(表2)。
在溶解氧变化过程中,西伯利亚鲟幼鱼鳃组织三磷酸腺苷酶活力亦不存在显著差异(P>0.05)(图7)。而当溶解氧质量浓度由18.5 mg/L降至7.5 mg/L时,西伯利亚鲟幼鱼肝脏和肌肉组织的碱性磷酸酶活力出现略微升高,当溶解氧质量浓度继续降低,二者碱性磷酸酶活力逐渐下降,再次恢复到7 mg/L时,肝脏碱性磷酸酶活力显著上升(P<0.05)(图8)。
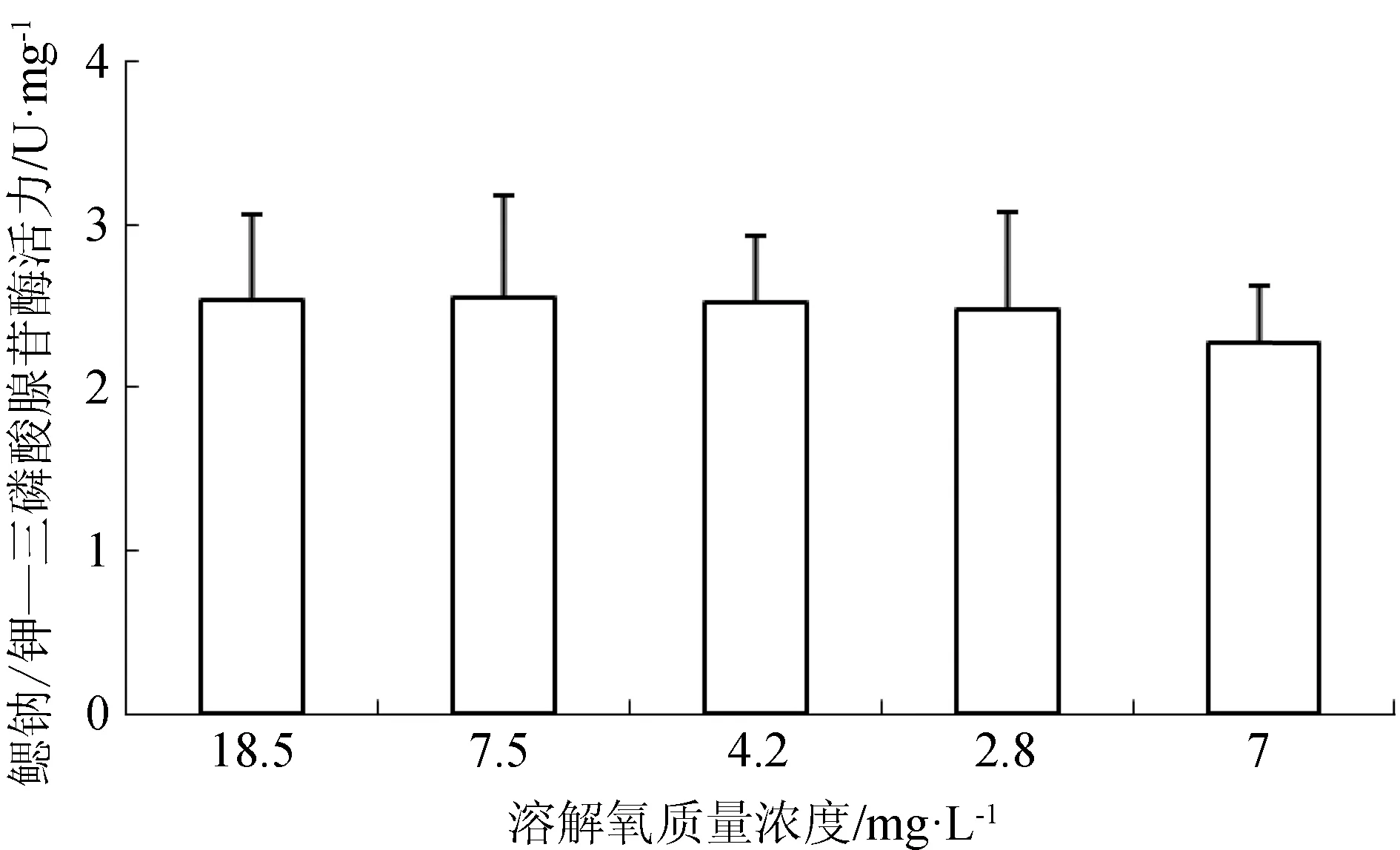
图7 不同溶解氧水平下西伯利亚鲟幼鱼鳃部三磷酸腺苷酶活力
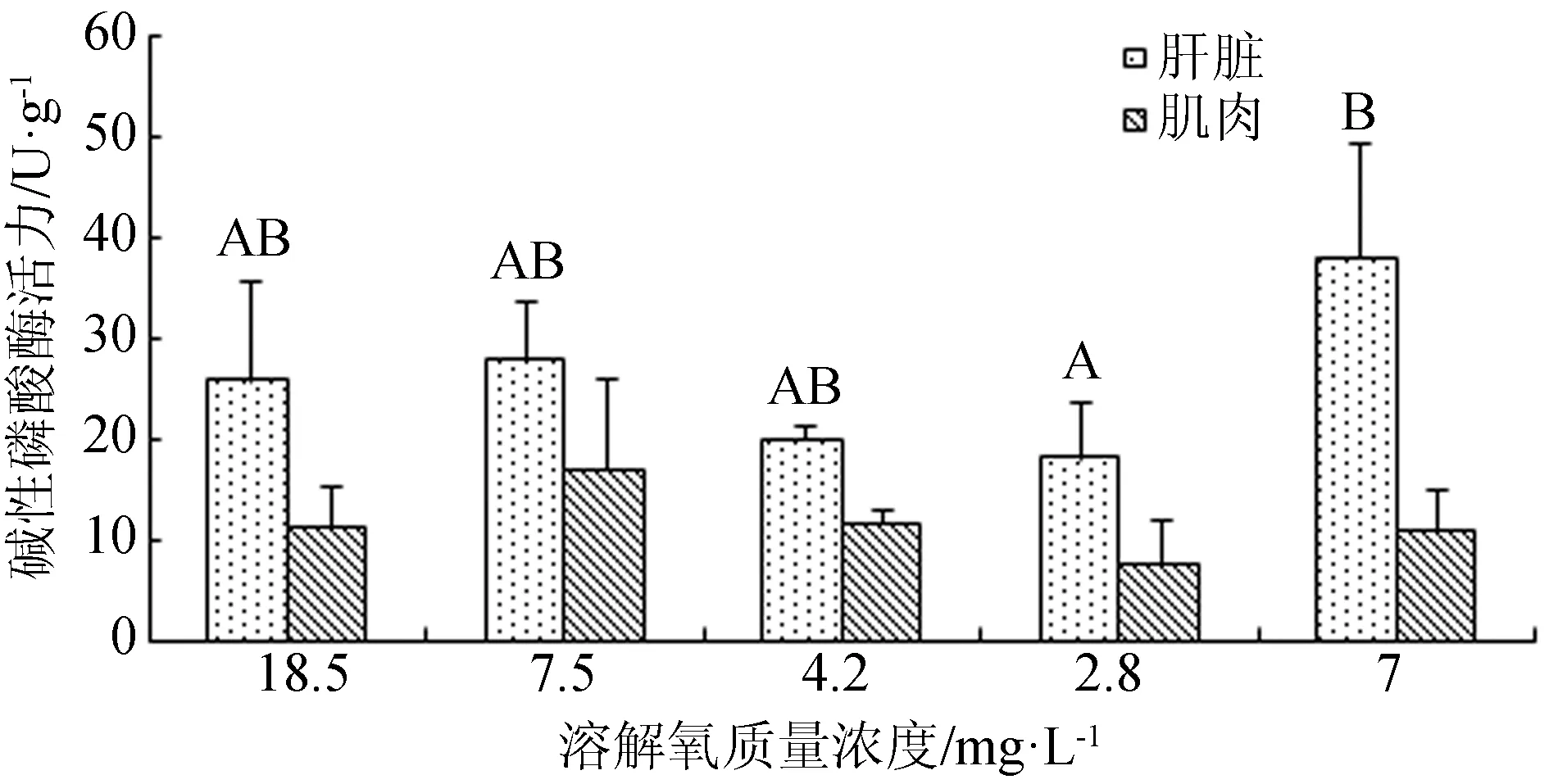
图8 不同溶解氧条件下鲟鱼肝脏和肌肉组织碱性磷酸酶活力
3 讨 论
3.1 低氧胁迫对西伯利亚鲟幼鱼血液基础指标的影响
水体过饱和溶解氧或者过低溶解氧均对鱼类的呼吸、代谢产生影响,导致鱼类生长[1]、消化[2]、抗氧化等多种生理活动变化,也会引起血氧亲和力[3]、血液中红细胞数目、血红蛋白含量[20]变化。有研究表明,经过30 d低氧驯化的鲤鱼(Cyprinuscarpio)红细胞数增加7%[21];经过12 h低氧驯化的大盖巨脂鲤(Colossomamacropomum)红细胞数增加53%,血红蛋白含量几乎增加1倍[22];鲫鱼(Cyprinusauratus)幼鱼经过2 h低氧处理,红细胞数增加约100%,血红蛋白增加约58%[23]。本研究结果显示,经过3 h的急性低氧处理,西伯利亚鲟幼鱼红细胞数目显著高于正常溶解氧状态下的红细胞数,超出10.8%,与以上结果一致,而血红蛋白含量低于正常溶解氧状态31.3%,这与鲫鱼的研究结果不同。相比鲫鱼,3 h的低氧驯化未引起西伯利亚鲟血液红细胞、血红蛋白含量发生较大程度变化,该结果提示鲟鱼的低氧适应过程反应速度较鲫鱼慢。当溶解氧质量浓度由18.5 mg/L降至7.5 mg/L时,血液红细胞数减少,待溶解氧继续降至4.2 mg/L时,红细胞数显著增加,溶解氧降至最低时,红细胞数反而减少,整个溶解氧变化过程血红蛋白含量无明显变化,可见溶解氧由18.5 mg/L逐渐下降过程中,西伯利亚鲟血液的红细胞数较血红蛋白含量变化更灵敏。血红细胞的氧亲和力还与血液pH有一定关系,pH升高通常血氧亲和力强,pH降低则会使血氧亲和力减弱[24]。本试验中,急性低氧处理3 h时,西伯利亚鲟血液pH显著低于正常溶解氧条件下pH,而在溶解氧变化过程中,低溶解氧质量浓度下的血液pH稍高,但无显著差异。溶解氧由18.5 mg/L降低过程中,溶解氧一定程度的降低可促进西伯利亚鲟幼鱼红细胞增加或从造血器官释放到循环系统,但当溶解氧质量浓度低于一定值(本试验中2.8 mg/L),可能超出鱼体调节范围,红细胞数没有增加,甚至有所减少。并且,短时间内溶解氧急剧变化,鱼体血红蛋白质量浓度和影响血氧亲和力的pH尚未得到调整,此时鱼体可能产生无氧呼吸。然而急性低氧处理3 h,西伯利亚鲟可能已实现自身调整以应对低氧胁迫,表现为增加血细胞数目,降低其在体内各组织的血氧亲和力,卸载更多氧气供给其他代谢反应,这与Wells等[22]对大盖巨脂鲤的低氧驯化研究结果一致。
3.2 低氧胁迫对西伯利亚鲟幼鱼抗氧化酶活力的影响
低氧环境下,鱼体的无氧代谢会导致乳酸和丙二醛等代谢产物的积累,低氧胁迫往往引起鱼体多种生理应激反应,并伴随过量活性氧分子的产生,这些都对生物体产生氧化胁迫,甚至引起氧化损伤[7,25]。Lushchak等[13]的研究显示鱼类在低氧胁迫时会提高某种抗氧化酶活力,以提高其应对恢复正常溶解氧环境带来的氧化应激能力。与之相反的是,本研究中,低氧处理3 h的鲟鱼肝脏和肌肉组织超氧化物歧化酶和过氧化氢酶活力均显著低于正常溶解氧组。而在溶解氧变化试验中,随着溶解氧质量浓度降低,肝脏超氧化物歧化酶和过氧化氢酶活力也随之降低,待恢复到正常溶解氧水平7.8 mg/L时,这两种抗氧化应激酶活力也得到恢复。然而肌肉超氧化物歧化酶活力在最低溶解氧处(2.8 mg/L)有所上升。综上,在溶解氧变化试验中,西伯利亚鲟幼鱼在低溶解氧水平时,肝脏的抗氧化能力有所降低,可能是在低氧状态下鱼体代谢率降低,并且通过降低相关酶活力来保护肝脏,而此时肌肉相应地有较高的抗氧化力,以保证机体内活性氧分子不过分积累,这与彭银辉等[26]对 鱼(Lizahaematocheila)的研究结果一致。而在急性低氧胁迫维持较长时间(3 h)的条件下,鱼体的肝脏和肌肉抗氧化功能可能受到影响,难以通过提高自身抗氧化酶来清除体内过量活性氧分子。
3.3 低氧胁迫对西伯利亚鲟幼鱼能量、代谢相关酶活力的影响
鱼类的各种生命活动需要机体能量的支持,三磷酸腺苷酶将三磷酸腺苷分解为二磷酸腺苷,并释放能量供给生命活动,而三磷酸腺苷酶的合成及活性均受到水体溶解氧变化的影响。王春枝等[18]对低氧胁迫下鲢鱼在心脏等组织中F1F0-三磷酸腺苷酶活性先升后降,认为在低氧应激条件下,通过自我调节促进鲤三磷酸腺苷酶活力升高,但当溶解氧降到一定程度,低氧造成鲢鱼线粒体内膜损伤而影响能量代谢。鳃组织鱼类的呼吸器官,是溶解氧进行气体交换的场所,本研究探讨了低氧胁迫下西伯利亚鲟鱼的鳃部三磷酸腺苷酶活力,结果显示急性低氧胁迫3 h以及在溶解氧变化过程中,鳃部三磷酸腺苷酶活力无明显变化,可能低氧胁迫未引起鱼体鳃部的能量调节。
碱性磷酸酶在鱼体内是一种参与代谢调控的非特异性磷酸水解酶,参与钙磷代谢,与机体的免疫密切相关[27-28],在机体的非特异性免疫反应中发挥重要作用。如果体内的碱性磷酸酶活力下降,则说明鱼体在环境胁迫下生命代谢能力减弱或免疫调节功能下降。本研究检测了在低氧胁迫下西伯利亚鲟肝脏和肌肉的碱性磷酸酶活力,结果显示,急性低氧处理3 h,西伯利亚鲟肝脏碱性磷酸酶活力显著低于正常溶解氧组。当溶解氧由18.5 mg/L降至7.5 mg/L时,西伯利亚鲟幼鱼肝脏和肌肉组织的碱性磷酸酶活力略微升高,随着溶解氧质量浓度继续下降,碱性磷酸酶活力逐渐降低,溶解氧恢复到7 mg/L时,碱性磷酸酶活力显著升高(P<0.05)。可见,低氧胁迫或者过高氧状态下,西伯利亚鲟碱性磷酸酶活力降低,此时鱼体代谢受到影响,非特异性免疫可能受到抑制,溶解氧质量浓度恢复后,鱼体代谢恢复正常。
3.4 小结
本研究系统分析了急性低氧(2.3 mg/L)和由高氧(18.5 mg/L)逐渐到低氧(2.8 mg/L)再恢复至7 mg/L这两种胁迫下,西伯利亚鲟幼鱼红细胞数、血红蛋白含量、抗氧化酶和能量代谢相关酶活力的变化情况。试验结果提示,西伯利亚鲟幼鱼在低氧胁迫时,可较快增加血液红细胞数量,并降低血红蛋白质量浓度,以降低血氧亲和力,来卸载更多氧气供组织使用;鱼体为保护肝脏,降低超氧化物歧化酶活力,而肌肉的超氧化物歧化酶活力在一定程度上有所升高;鳃部三磷酸腺苷酶活力并未增强,可能低氧胁迫未引起鱼体鳃部的能量调节;西伯利亚鲟幼鱼肝脏和肌肉碱性磷酸酶活力随溶解氧降低而受到抑制,自身代谢调控和非特异性免疫受到影响。但从低氧状态恢复到正常溶解氧状态时,各项指标均能恢复到正常状态,鱼体并未受到严重影响。
[1] Wei L Z, Zhang X M, Huang G Q. Effects of limited dissolved oxygen supply on the growth and energy allocation of juvenile Chinese shrimp,Fenneropenaeuschinensis[J]. Journal of World Aquaculture Society, 2009, 40(4):483-492.
[2] 张钊, 李荣, 郭惠, 等.不同溶解氧水平对工厂化养殖大菱鲆生长及肌肉生化组分的影响[J]. 水产科学, 2014, 33(8):520-523.
[3] 胡国宏,孙广华,朱世成,等.低溶氧量对怀头鲇呼吸代谢耗氧率的影响[J].动物学杂志, 2002, 37(2):46-48.
[4] 沈凡, 樊启学, 杨凯, 等. 不同溶氧条件下黄颡鱼免疫机能及抗病力的研究[J].淡水渔业, 2010, 40(4):44-49.
[5] Chabot D,Dutil D. Reduced growth of Atlantic cod in non-lethal hypoxia conditions [J]. Journal of Fish Biology, 1999, 55(3):472-491.
[6] Karim M R, Sekine M, Ukita M. Simulation of eutrophication and associated occurrence of hypoxia and anoxia condition in a coastal bay in Japan [J]. Marine Pollution Bulletin, 2002, 45(1/12):280-285.
[7] Wihelm F D, Torres M A, Zaniboni-filhoe, et al. Effects of different oxygen tensions on weight gain, feed conversion, and antioxidant status in piaparaLeporinuselongates(Valeneiennes, 1847) [J]. Aquaculture, 2005, 244(1/4):349-357.
[8] Randall D. The control of respiration and circulation in fish during exercise and hypoxia [J]. Journal of the Royal Society of Medicine, 1999, 92(4):186-188.
[9] Silkin Y A, Silkina E N. Effects of hypoxia on physiological biochemical blood parameters in some marine fish [J].Journal of Evolutionary Biochemistry and Physiology, 2005, 41(5):527-532.
[10] Gamperl A K, Farrell A P. Cardiac plasticity in fishes: environmental influences and intraspecific differences [J]. The Journal of Experimental Biology, 2004, 207(15):2539-2550.
[11] Bickler P E, Buck L T. Hypoxia tolerance in reptiles, amphibians, and fishes: life with variable oxygen availability [J]. Annual Review of Physiology, 2007, 69(69):145-170.
[12] Lushchak V I, Bagnyukov T V, Lushchak O V, et al.Hypoxia and recovery perturb free radical processes and antioxidant potential in common carp (Cyprinuscarpio) tissues [J]. International Journal of Biochemistry and Cell Biology, 2005, 37(6):1319-1330.
[13] Lushchak V I, Bagnyukov T V. Effects of different environmental oxygen levels on free radical processes in fish [J]. Comparative Biochemistry and Physiology Part B, 2006, 144(3):283-289.
[14] Wu R S. Hypoxia: from molecular responses to ecosystem responses [J].Marine Pollution Bulletin, 2002, 45(1/12):35-45.
[15] Cooper R U, Clough L M, Farwell M A, et al. Hypoxic-induced metabolic and antioxidant enzymatic activities in the estuarine fishLeiostomusxanthurus[J]. Journal of Experimental Marine Biology & Ecology, 2002, 279(1/2):1-20.
[16] Johnton I A. Calcium regulatory protein and temperature acclimation of actomyosin ATPase from a eurythermal teleost(Carassois) [J]. Comparative Biochemistry and Physiology Part B,1979, 129(2):163-167.
[17] 冯广朋, 庄平, 章龙珍, 等. 饥饿期间中华鲟幼鱼血液与肝脏酶活力的变化[J].海洋渔业, 2011, 33(2):165-171.
[18] 王春枝, 李忠, 梁宏伟, 等. 低氧胁迫对鲢线粒体ATP酶活性及F1-δ基因表达的影响[J]. 中国水产科学, 2014, 21(3):454-463.
[19] 金丽, 赵娜, 周传江,等.饥饿对胭脂鱼血液指标及造血的影响[J]. 水生生物学报, 2012, 36(4):665-673.
[20] Cameron J N. The influence of environmental variables on the haematology of pinfish (Lagodonrhomboides), and striped mullet (Mugilcephalus) [J]. Comparative Biochemistry and Physiology, 1970, 32(2):175-192.
[21] Weber R E, Lykkeboe G. Respiratory adaption in carp blood influences of hypoxia, red cell organic phosphates, divalent cations and CO2on hemoglobin-oxygen affinity [J]. Journal of Comparative Physiology, 1978, 128(2):127-137.
[22] Wells R M G,Grigg G C, Beard L A, et al. Hypoxia responses in a fish from a stable environment: blood oxygen transport in the Antarctic fishPagotheniaborchgrevinki[J]. The Journal of Experimental Biology, 1989(141):97-111.
[23] 张曦, 付世建, 彭姜岚, 等. 急性低氧对鲫鱼幼鱼血液基础指标的影响[J]. 重庆师范大学学报:自然科学版, 2011, 28(4):19-22.
[24] Tetents V, Wells R M G. Antarctic fish blood: respiratory properties and the effects of thermal acclimation [J]. The Journal of Experimental Biology, 1984, 109(3):265-279.
[25] 洪磊,张秀梅. 环境胁迫对鱼类生理机能的影响[J]. 海洋科学进展,2004, 22(1):114-121.
[26] 彭银辉, 黄国强, 李洁, 等.溶氧水平对梭鱼幼鱼能量代谢与氧化应激的影响[J]. 广西科学, 2013, 20(4):294- 298.
[27] 罗克. 动物粘膜免疫系统研究进展[J]. 福建畜牧兽医, 2000, 22(5):44-49.
[28] Cheng T C. The role of lysosomal hydrolases in molluscan cellular response to immunologic challenge[G]// Bulla L A J, Cheng T C. Invertebrate Models for Biomedical Research. New York: Springer US, 1978:59-71.
EffectsofHypoxiaonPhysiologicalStatusofSiberianSturgeonAcipenserbaeriJuveniles
WANG Xiaowen, ZHU Hua, HU Hongxia, MA Guoqing
( Beijing Key Laboratory of Fishery Biotechnology, Beijing Fisheries Research Institute, Beijing 100068, China )
Effects of acute hypoxia stress and drastic changes in dissolved oxygen levels from 18.5 mg/L to 2.8 mg/L then recovery to 7 mg/L on blood parameters, antioxidant and metabolism were studied in Siberian sturgeonAcipenserbaeriwith body weight of (19.46±4.9) g. Erythrocyte count, total hemoglobin concentration, activities of antioxidant enzymes in livers and muscle, and activities of Na+/K+-ATPase in gills were measured in the experiment. The results showed that erythrocytic number was elevated, while hemoglobin concentration and pH of whole blood under hypoxia stress were lower than those in the control group. Hypoxia stress also depressed the activities of superoxide dismutase (SOD), catalase (CAT) and alkaline phosphatase (AKP) in juvenile sturgeon. Nevertheless, there were no obvious changes in the activities of Na+/K+-ATPase in gills of Siberian sturgeon juveniles. Based on the results above, it is suggested that hypoxia stress could easily lead to the enhancement of erythrocyte amount, also reduce the activities of antioxidant and metabolism enzymes in livers or muscle of the juveniles. However, sturgeon juveniles could not achieve energy adjustment in gills itself within a short time.
Acipenserbaeri; blood parameter; antioxidant enzyme; ATPase; AKP enzyme
10.16378/j.cnki.1003-1111.2016.05.001
S965.199
A
1003-1111(2016)05-0459-07
2015-10-23;
2016-04-18.
国家科技支撑计划项目(2015BAD25B01,2012BAD26B05);现代农业产业技术体系北京市鲟鱼、鲑鳟鱼创新团队项目(SCGWZJ20161102,SCGWZJ20161104);北京市农林科学院冷水性鱼类科技创新团队项目(JNKST201611).
王晓雯(1989—),女,助理工程师;研究方向:水产免疫生理. E-mail:wxw211@126.com.通讯作者:朱华(1972—),男,研究员;研究方向:水产养殖与生理生态. E-mail:zhuhua@bjfishery.com.
