黑曲霉产β-葡萄糖苷酶工艺的神经网络优化
2016-12-17秦小明钟赛意陈建平
高 倩,秦小明,钟赛意,陈建平
(广东海洋大学食品科技学院,广东 湛江 524088)
黑曲霉产β-葡萄糖苷酶工艺的神经网络优化
高 倩,秦小明,钟赛意,陈建平
(广东海洋大学食品科技学院,广东 湛江 524088)
在单因素试验研究的基础上,采用Box-Behnken中心组合设计,利用JMP 7.0中的神经网络平台,在接种量、初始pH、发酵时间和装液量等4个方面对黑曲霉CⅠCC 2475发酵产β-葡萄糖苷酶工艺条件进行优化,以期获得高酶活力的β-葡萄糖苷酶。结果表明:接种量约11.0%、初始pH值5.6、发酵时间130 h以及装液量70.0mL条件下,黑曲霉发酵产β-葡萄糖苷酶的酶活力达到最大值118.73 U/mL,与单因素优化前61.01 U/mL相比,酶活力提高了48.61%。
黑曲霉CⅠCC2475;β-葡萄糖苷酶;神经网络
β-葡萄糖苷酶(β-glucosidase,EC 3.2.1.21)广泛存在于各类生物体中,属于纤维素酶类水解酶,作用于纤维寡糖中的 β-1,4糖苷键及糖苷类物质中的羟基或芳香基团与糖基之间的糖苷键,在食品[1]、医药[2]、化工[3]等领域具有重要的作用,在糖苷合成过程中也发挥着重要作用[4]。近年来,国内研究学者不断研究β-葡萄糖苷酶的分离纯化[5-6],但结果仍不理想,分离纯化出的β-葡萄糖苷酶的酶活性与国外研究相比较低,因此仍需要加大对β-葡萄糖苷酶分离纯化的研究。
研究结果发现,从微生物中提取的β-葡萄糖苷酶活性远高于植物中提取的β-葡萄糖苷酶[7]。黑曲霉生长速度较快,发酵周期较短,其发酵生产的酶制剂安全可靠,不含有毒物质,其所产β-葡萄糖苷酶的安全性是国际公认的[8]。在黑曲霉产β-葡萄糖苷酶的研究中,常用的优化分析方法是响应面分析,但是响应面分析有一定的局限性,仅适用于二次函数模型的模拟[9]。神经网络是以已知分类的数据作为输入,在初始阈值和权值的作用下计算输出结果,再与实际结果比较得到实验误差;通过不断调整阈值和权值减少误差,使输出结果近似实际值。神经网络具有强大的分类和预测能力,对于非线性系统具有较好的优化处理能力,可保存已训练好的模型,输入数据后可准确快速预测输出结果[10]。基于此优势,许多国内外研究学者将神经网络优化应用于微生物发酵过程中[11-12]。笔者在单因素试验研究的基础上,利用JMP 7.0中的神经网络,在接种量、初始pH、发酵时间和装液量等4个方面对黑曲霉CⅠCC2475发酵产β-葡萄糖苷酶工艺条件进行优化,以期获得较高酶活力的 β-葡萄糖苷酶。
1 材料与方法
1.1 材料与试剂
1.1.1 菌株 黑曲霉(Aspergillus niger) CⅠCC 2475,购于中国工业微生物菌种保藏管理中心。将菌种接入PDA斜面培养基中,25℃ 活化5 d。
1.1.2 培养基 种子培养基(g/L):可溶性淀粉20、葡萄糖 10、MgSO4·7H2O 1、KH2PO41、酵母膏 5、自然pH(不加调整),121℃灭菌20min;
液体发酵培养基(g/L)[13]:麸皮 20、硫酸铵1.5、KH2PO42、MgSO4·7H2O 1、吐温-80 2mL,pH 5.0,121℃灭菌20min。
1.1.3 其他试剂 对硝基苯β-D-葡萄糖苷(pNPG)购于上海源叶生物科技有限公司,柠檬酸,柠檬酸钠,无水碳酸钠均为国产分析纯。
1.2 仪器与设备
净化工作台(SW-CJ-270,上海博讯实业有限公司医疗设备厂),恒温培养箱(SPX-250B-Z,上海博讯实业有限公司医疗设备厂),紫外可见分光光度计(UⅤ-3200PC,上海精密科学仪器有限公司),全温培养摇床(HZQ-A,苏州威尔实验用品有限公司),立式压力蒸汽灭菌器(LDZX-30KBS,上海申安医疗器械厂),高速冷冻离心机(CR22GⅢ,HⅠTACHⅠ公司)
1.3 方法
1.3.1 发酵产酶实验 在 PDA斜面培养基中取一环菌种,接种于装有200mL种子培养液的500mL摇瓶中,28℃下培养48 h。按8%的接种量(4mL)接种于装有50mL发酵培养液的250mL摇瓶中,28℃、170 r/min下培养4 d。
1.3.2 β-葡萄糖苷酶粗酶液的提取 从摇瓶中取适量发酵液,4 500 r/min条件下离心10min,取上清液,柠檬酸-柠檬酸钠缓冲液(pH 5.0)稀释5倍,用于酶活性的测定。
1.3.3 β-葡萄糖苷酶活性测定[14]1.8mL、0.1 mol/L柠檬酸-柠檬酸钠缓冲液(pH 5.0)中加入0.1mL、10 mmol/L对硝基苯β-D-葡萄糖苷(pNPG)作为反应底物,加入0.1mL酶液,45℃反应30min后加入2mL、1 mol/L Na2CO3终止反应并显色,在410 nm处测定吸光值。45℃时,每min每mL酶液酶解1 μmol pNPG的酶活性浓度为一个酶活力单位(U/mL)。
1.3.5 单因素试验
1.3.5.1 接种量对产酶的影响 以体积分数计,分别以4%、6%、8%、10%和12%的接种量接种于装液量为50mL的250mL摇瓶中,其他因素不变。
1.3.5.2 初始pH对产酶的影响 用1 mol/L HCl及1 mol/L KOH调节发酵培养基初始pH,使发酵培养基的初始pH分别为4.0、4.5、5.0、5.5、6和6.5,其他因素不变。
1.3.5.3 发酵时间对产酶的影响 将发酵培养时间分别设定为 24、48、72、96、120、144、168和192 h,其他因素不变。
1.3.5.4 装液量对产酶的影响 在250mL摇瓶中分别加入50、75、100、125和150mL培养液,其他因素不变。
1.3.6 神经网络模型建立及产酶工艺条件优化本研究以接种量(X1)、初始pH(X2)、发酵时间(X3)、装液量(X4)4个试验因素为主,以酶活力(U/mL)为响应因素。采用Box-Behnken设计因素水平编码表。采用JMP 7.0数据处理软件中的神经网络平台,建立神经网络模型及对产酶工艺条件的优化分析。
2 结果与讨论
2.1 接种量对产酶的影响
理论上,当接种量在一定范围内增加时,酶活力会随之增大,但接种量超过饱和量时,菌种的存活空间变小,产酶能力变低。从图1可见,在一定范围内酶活力随着接种量的增加而增大,当接种量达到10%时,酶活力达到最大。

图1 接种量对黑曲霉发酵生产β-葡萄糖苷酶的影响Fig.1 Effect of inoculation amount on production of β-glucosidase from Aspergillus niger
2.2 初始pH对产酶的影响
在适合pH的条件下,菌种的产酶能力会提高。从图2可见,随着pH值的增大,菌种的产酶能力先提高后降低,pH在5.5时酶活力相对较高。

图2 初始pH对黑曲霉发酵生产β-葡萄糖苷酶的影响Fig.2 Effect of fermentation pH on production of β-glucosidase from Aspergillus niger
2.3 发酵时间对产酶的影响
发酵时间也是影响菌种发酵产酶的重要因素之一。由图3可见,黑曲霉产酶活力随着发酵时间的增加而增加,在发酵120 h后酶活力逐渐趋于平稳。
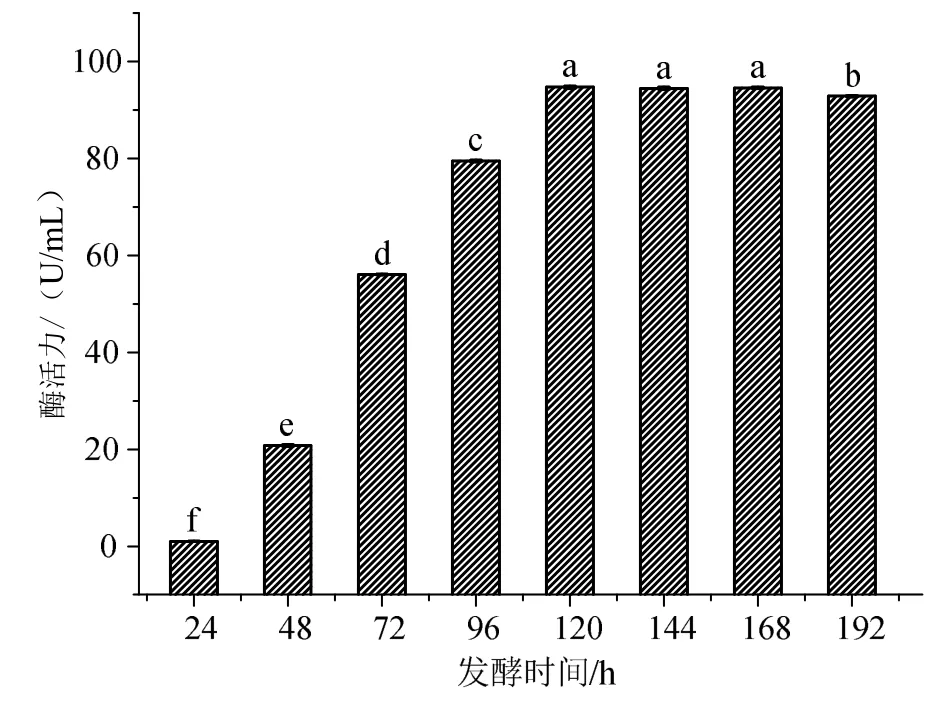
图3 发酵时间对黑曲霉发酵生产β-葡萄糖苷酶的影响Fig.3 Effect of fermentation time on production of β-glucosidase from Aspergillus niger
2.4 装液量对产酶的影响
装液量即为菌种的生长及产酶空间,同样影响酶的活力。从图4可见,装液量在25mL到75mL范围内,酶活力增加的比较缓慢,当装液量为75mL时,酶活力达到最大值。
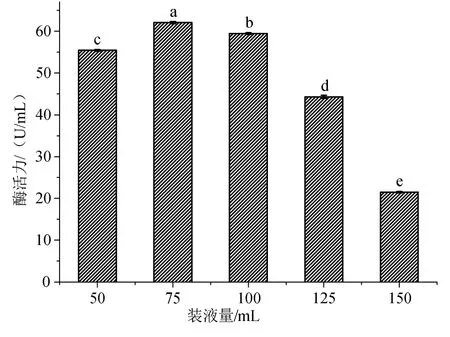
图4 装液量对黑曲霉发酵生产β-葡萄糖苷酶的影响Fig.4 Effect of broth content on production of β-glucosidase from Aspergillus niger
2.5 神经网络模型建立及产酶工艺条件优化
2.5.1 实验设计 在单因素实验结果基础上,采用Box-Behnken 设计因素水平编码表(表1)。试验结果如表2。
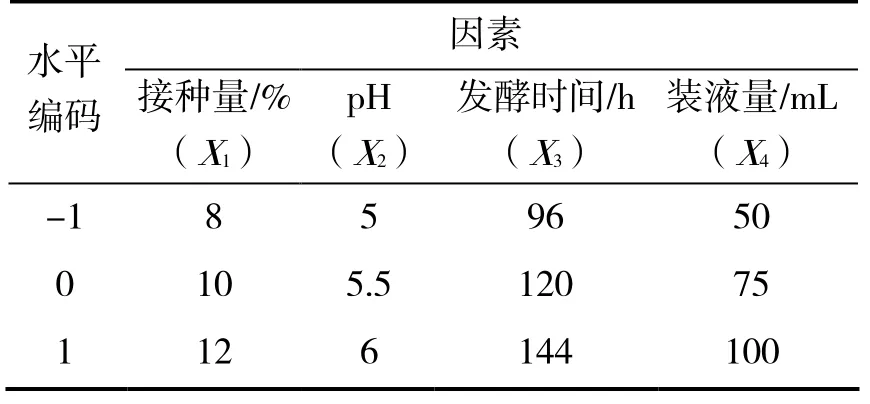
表 1 因素水平编码Table 1 Code of factors and levels
2.5.2 神经网络模型的建立 根据试验结果,选用“K折叠”交叉验证的方法建立神经网络模型。
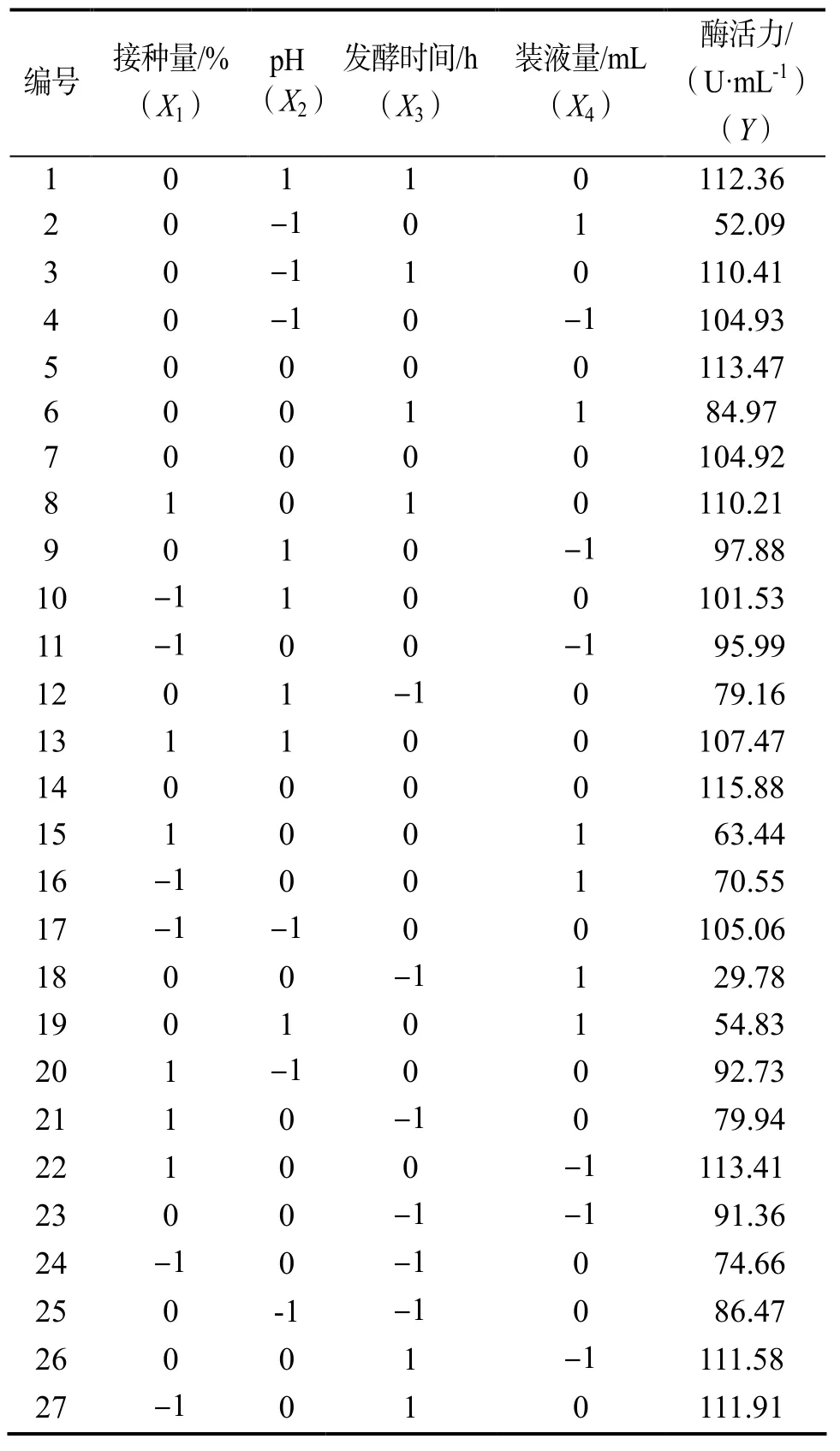
表2 Box-Behnken设计及结果Table 2 Design and results by Box-Behnken
在经过多次对交叉验证组数K及隐藏节点的神经网络训练之后,确定交叉验证组数K值为5,采用4×5×1结构的三层神经网络(图5),即4个输入神经元,分别为接种量(X1)、初始pH(X2)、发酵时间(X3)和装液量(X4);5个隐含层神经元;1个输出神经元,代表响应指标酶活力(Y)。设置各参数值,隐藏节点数5,过拟合罚项0.001,历程数16,最大迭代数50,收敛准则0.000 01,交叉验证组数K为5,执行神经网络模型的拟合迭代过程,拟合决定系数r2值为0.99以上(图6),说明4×5×1结构的三层神经网络模型具有较好的预测能力。
2.5.3 产酶工艺条件优化 固定接种量(X1)、初始 pH(X2)、发酵时间(X3)和装液量(X4)4个因素中的两个因素为中间水平,作三维曲面图,将得到4个因素对黑曲霉发酵产β-葡萄糖苷酶的酶活力的影响规律进行预测分析,结果见图7。

图5 神经网络结构Fig.5 Structure of artificial neural network

图6 实际值与预测值相关性Fig.6 The correlation of actual values and the predicted values
从图7a、7b、7c中可见,在pH值约低于5.5时,酶活力逐渐增加;而高于5.5后,酶活力逐渐降低,可见环境的酸碱程度会明显影响酶的活力。有相关报道称,酶活力降低的原因可能是菌体的生长及产物的合成都有一个最适的酸碱环境,偏酸或偏碱的环境会抑制菌体内的代谢,影响黑曲霉的正常生长及产物的合成,进而导致产酶量及酶活力降低[15-16]。
图7a、7d和7e中显示,随着发酵时间的延长,酶活力明显增加,在发酵时间达到约130 h后逐渐平稳。这可能因为在一定的发酵时间内,菌体量的增加使产酶量逐渐增加,随着发酵时间的延长,培养基内的营养减少,代谢产物积累,使菌体停止产酶[16]。
图7b、7d和7f中显示,酶活力随着接种量的增加,呈现出先增加后降低的趋势。接种量过低,产酶活力则低;接种量过大,可能会使菌体的浓度增大,培养液对菌体的供养不足,则产酶降低[17]。图中显示在约小于11%时,酶活力逐渐增加;当约大于11%后,酶活力逐渐下降,这说明接种量存在一个最适值。
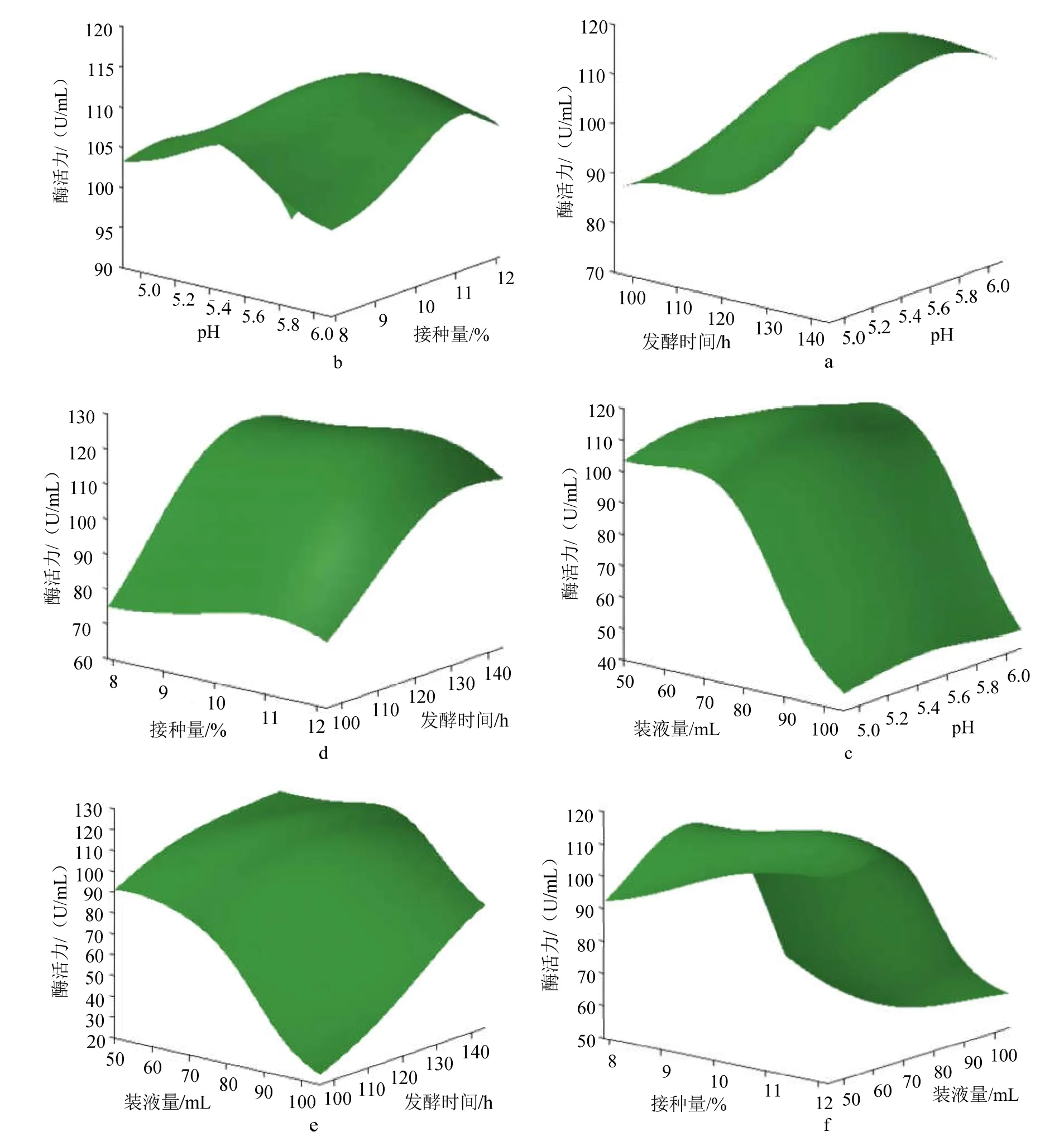
图7 接种量、初始pH、发酵时间和装液量对产酶的影响Fig.7 Effect of inoculation amount、fermentation pH、fermentation time and broth content on production of enzymes
从图7c、7e和7f中可见,装液量对黑曲霉产酶的影响显著,在装液量大于约70mL时,酶活力有显著下降的趋势。这可能因为黑曲霉为好氧微生物,装液量过大,则溶氧量减少,致使菌体的生长缓慢,产酶随之降低[18]。
根据接种量、初始 pH、发酵时间和装液量四个因素对黑曲霉产β-葡萄糖苷酶的影响规律,采用JMP 7.0软件中的预测刻画器,优化黑曲霉产酶的条件,预测刻画见图8。
从图8中预测得出,当接种量为11.09%,初始pH为5.65,发酵时间为130.91 h,装液量为70.38mL时,黑曲霉发酵生产β-葡萄糖苷酶的酶活力达到最大(120.33 U/mL)。考虑实验条件的可行性及易控性,确定黑曲霉发酵生产β-葡萄糖苷酶最佳的工艺参数为:接种量为11.0%,初始pH为5.6,发酵时间为130 h,装液量为70mL。
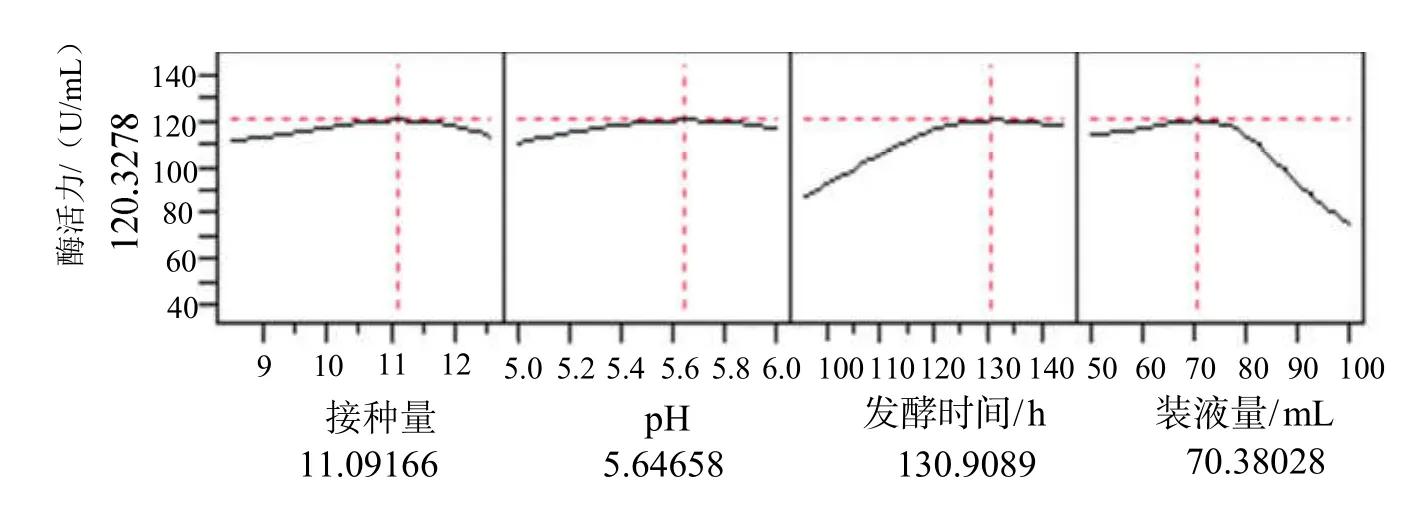
图8 神经网络模型的预测刻画器Fig.8 Prediction plot of the neural network
2.5.4 神经网络模型预测结果的验证 为验证使用神经网络建立的模型是否符合实际情况,随机选取3组新的水平因素组合来测试该模型(表3)[19],并对模型得出的最优值进行结果验证。
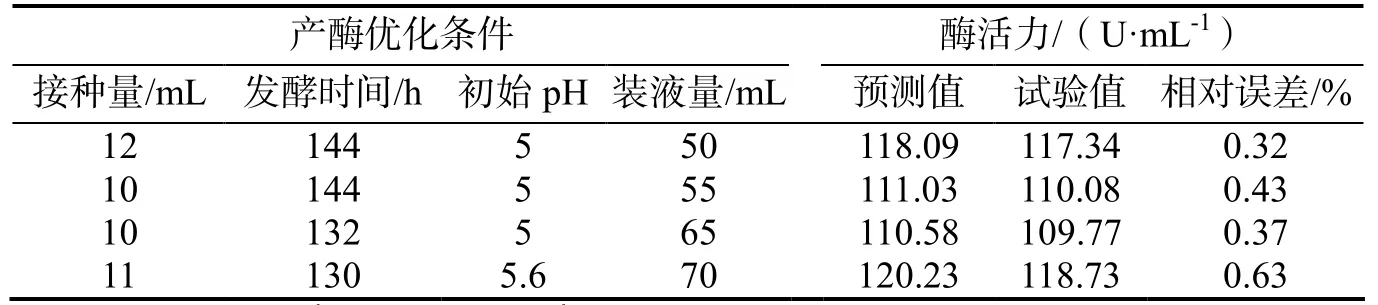
表3 神经网络模型预测值与验证值比较Table 3 Predicted values of neural network model and experimental values
从表3的结果可看,建立的神经网络模型对试验结果预测较为准确,说明使用该神经网络建立的模型可对黑曲霉产β-糖苷酶具有很好的预测能力。经过单因素及神经网络优化,酶活力提高48.61%。孙海彦[20]等通过响应面优化黑曲霉产 β-葡萄糖苷酶,使酶活力提高了46%,凌宏志[21]等通过响应面优化黑曲霉HDF05产β-葡萄糖苷酶,仅使酶活力提高了23.9%,说明本研究的结果处于较优水平。
3 结论
在单因素试验研究的基础上,采用Box-Behnken中心组合设计,利用JMP 7.0软件建立了黑曲霉发酵生产 β-葡萄糖苷酶的神经网络模型,从接种量、初始 pH、发酵时间及装液量等 4个方面探讨了其交互作用,以及对黑曲霉生产β-葡萄糖苷酶酶活力的影响,得到最佳发酵工艺条件为接种量11.0%、初始pH 5.6发酵时间130 h、装液量70.0mL,制备的β-葡萄糖苷酶的最大酶活力为118.73 U/mL,与单因素优化前(61.01 U/mL)相比提高了48.61%。
[1]GOPAL S,ⅤERMA A K,ⅤⅠNOD K.Catalytic properties,functional attributes and industrial applications of β-glucosidases[J].3 Biotech,2016,6(1):1-14.
[2]LⅠMA F S D,ⅠDA EⅠ.Optimisation of soybean hydrothermal treatment for the conversion of β-glucoside isoflavones to aglycones[J].Food Science and Technology,2014,56(2):232-239.
[3]LⅠ D,LⅠ X,DANG W,et al.Characterization and application of an acidophilic and thermostable β-glucosidase from Thermofilum pendens [J].Journal of Bioscience and Bioengineering.2013,115(5):490-496.
[4]GUO D,XU Y,KANG Y,et al.Synthesis of octyl-β-D-glucopyranoside catalyzed by Thai rosewood β-glucosidase-displaying Pichia pastoris in an aqueous/organic two-phase system[J].Enzyme & Microbial Technology,2016,85:90-97.
[5]ŞENER A.Extraction, partial purification and determination of some biochemical properties of β-glucosidase from Tea Leaves(Camellia sinensis L.)[J].Journal of Food Science & Technology,2015,52(12):8322-8328.
[6]AŠⅠĆ A,BEŠⅠĆ L,MUHOⅤⅠĆⅠ,et al.Purification and Characterization of β-Glucosidase from Agaricus bisporus(White Button Mushroom)[J].Protein Journal,2015,34(6):1-9.
[7]李晓东,周明,杨丽娜,等.黑曲霉β-葡萄糖苷酶的酶学特性研究[J].微生物学杂志,2014(3):14-18.
[8]先天敏,陈介南,张林.黑曲霉改良株C112产β-葡萄糖苷酶的诱导及条件优化[J].中南林业科技大学学报,2013,33(11):154-161.
[9]DESAⅠ K M,SURⅤASE S A,SAUDAGAR P S,et al.Comparison of artificial neural network(ANN)and response surface methodology(RSM)in fermentation media optimization: Case study of fermentative production of scleroglucan[J].Biochemical Engineering Journal,2008,41(3):266-273.
[10]蔡强,王君君,李海生,等.基于神经网络的食品安全评价模型构建研究[J].食品科学技术学报,2014,32(1):69-76.
[11]吴丹,杨基先,李昂,等.基于 BP神经网络的产絮菌发酵参数的优化[J].哈尔滨工业大学学报,2013(10):30-35.
[12]FRANCO-LARA E,LⅠNK H,WEUSTER-BOTZ D.Evaluation of artificial neural networks for modelling and optimization of medium composition with a genetic algorithm[J].Process Biochemistry,2006,41(10):2200-2206.
[13]徐星,肖华,黄琳琳,等.黑曲霉胞外β-葡萄糖苷酶的纯化及酶学性质的研究[J].天津科技大学学报,2015(5):15-19.
[14]ORTEGA N,BUSTO M D,PEREZ-MATEOS M.Optimisation of β-glucosidase entrapment in alginate and polyacrylamide gels[J].Bioresource Technology,1998,64(2):105-111.
[15]ONCU S,TARⅠ C,UNLUTURK S.Effect of various process parameters on morphology,rheology,and polygalacturonase production by Aspergillus sojae in a batch bioreactor[J].Biotechnology Progress,2007,23(23):836-45.
[16]赵林果,王平,宗雅冬,等.黑曲霉分泌β-葡萄糖苷酶过程中 pH 值的调控[J].化学与生物工程,2008(12):48-51.
[17]马旭光,张宗舟,霍建泰,等.航天诱变黑曲霉 ZM-8发酵玉米秸秆粉产 β-葡萄糖苷酶的培养基组分优化[J].食品工业科技,2012,33(6):236-239,243.
[18]王蕊,王林风,闫德冉,等.毕赤酵母表达 β-葡萄糖苷酶中试条件优化[J].食品科技,2015(12):15-19.
[19]张良,刘书成,章超桦,等.神经网络优化牡蛎的高密度 CO2杀菌工艺[J].农业工程学报,2011(12):369-373.
[20]孙海彦,王茜,刘恩世,等.响应面方法优化黑曲霉产 β-葡萄糖苷酶培养基成分的研究[J].轻工科技,2012,167(10): 8-9,20.
[21]凌宏志,葛菁萍,平文祥,等.响应面法优化黑曲霉HDF05产β-葡萄糖苷酶过程参数[J].生物工程学报,2011(3): 419-426.
(责任编辑:陈庄)
Optimization of Conditions for Producing β-Glucosidase by Aspergillus niger Based on Neural Network
GAO Qian,QⅠN Xiao-ming,ZHONG Sai-yi,CHEN Jian-ping
(College of Food Science and Technology,Guangdong Ocean University,Zhanjiang 524088,China)
Based on the single factor experiments,this research obtained high enzyme activity of β-glucosidase from Aspergillus niger CⅠCC 2475 by using Box-Behnken center-united design and JMP 7.0 neural network,the conditions of β-glucosidase were optimized by the control of inoculation amount,fermentation pH,fermentation time and broth content.The results showed that: the inoculation amount was about 11.0%,the initial pH of 5.6,fermentation time was 130 hours and broth content 70.0mL,the enzyme activity of Aspergillus niger fermentation β-glucosidase reached the maximum(118.73 U/mL),compared with the previous optimization(61.01 U/mL),the enzyme activity increased by 48.61%.
Aspergillus niger CⅠCC 2475;β-glucosidase;neural network
TQ925
A
1673-9159(2017)01-0087-07
10.3969/j.issn.1673-9159.2016.01.014
2016-07-11
广东省教育厅创新强校工程项目(2013050214)
高倩(1990—),女,硕士研究生,研究方向农产品加工与贮藏。
秦小明,博士,教授,研究方向亚热带食品新资源开发与利用。E-mail:xiaoming0502@21cn.com
