小尾寒羊TFB2M基因遗传变异分析
2016-12-12陈晓勇敦伟涛赵兴波
陈晓勇,王 薇,敦伟涛,赵兴波⋆
(1.中国农业大学动物科技学院,北京 100193;2.河北省畜牧兽医研究所,河北 保定 071000)
小尾寒羊TFB2M基因遗传变异分析
陈晓勇1,2,王薇1,敦伟涛2⋆,赵兴波1⋆
(1.中国农业大学动物科技学院,北京100193;2.河北省畜牧兽医研究所,河北保定071000)
试验旨在分析小尾寒羊线粒体基因转录因子B2(M itochondrial Transcription Factors B2,TFB2M)基因遗传变异,采集小尾寒羊血液,采用直接测序方法,对该基因8个外显子、5’-UTR和3’-UTR多态位点进行分析。结果发现,编码区错义突变1个(G226A,GCC→ACC)导致76位苏氨酸变为丙氨酸,5’-UTR发现一处颠换(C58G);3’-UTR有三个突变位点(C98G、A206G和A429T)。生物学软件对启动子和5’-UTR转录因子预测结果显示共有54个转录因子结合位点,5’-UTR 58bp处没有结合位点,因此,该突变并没有改变结合位点。本研究为绵羊TFB2M基因变异及其功能研究奠定了分子生物学基础。
线粒体转录因子B2;小尾寒羊;变异;转录因子
线粒体是细胞中具有独特结构和功能的细胞器,其功能的精细调节有赖于细胞核基因组与线粒体基因组之间的相互调控。线粒体基因组的复制、转录和翻译需要各种不同的核基因产物,而一些核基因的功能与线粒体基因(Mitochondrial DNA,mtDNA)转移、线粒体蛋白质的生物合成等有关,两者不但在生物发生、生物合成方面需要相互协调,更重要的是通过相互协调作用来调节呼吸链亚基的表达,从而维持线粒体呼吸链功能。在细胞内,线粒体遗传系统与细胞核遗传系统相互协作进行生物合成。
线粒体转录因子B2(Mitochondrial Transcrip-tion Factors B2,缩写为TFB2M或者mtTFB2)是调控哺乳动物线粒体基因(mtDNA)表达调控的重要因子[1],由于与酒精酵母转录因子mtTFB序列相似而被发现[2],二者都是核基因编码并转运到线粒体的蛋白质,其表达受核转录因子,如过氧化物体增殖活化受体γ辅活化因子1(Peroxisome prol i feratoractivated receptor(PPAR)-γcoactivatorα,PGC-1α)、核呼吸因子1和2(nuclear respiratory factorsNRF-1,NRF-2)等严格调控[3-5]。核转录因子的发现使细胞核调控线粒体呼吸功能途径的研究有了突破性进展。TFB2M蛋白质作为转录元件在转录起始位置与D-loop结合,组成转录起始装置启动mtDNA转录[1,6]。有研究报道发现牛T F A M 基因编码区无多态性位点,但启动子区域存在2个相距9 bp紧密连锁的A/C(TFAM Hae III)和C/T(TFAM Mbo I)SNP位点。通过对和牛×利木赞群体进行基因分型,证实2个SNPs位点都显著影响大理石纹和皮下脂肪沉积[7-8]。因此,分析该基因遗传变异将有利于揭示线粒体基因功能及其遗传效应。
目前,还未见绵羊TFB2M基因变异研究报道,本试验以小尾寒羊为研究对象分析TFB2M基因变异,将为该基因功能及其与表型性状的关系研究提供参考。
1 材料与方法
1.1试验动物和D D N N A A提取选择成年小尾寒羊10只个体,采用颈静脉方法用5 mL注射器采集全血(每只羊一个注射器),每个样品采集血液1.5 mL左右装入备好含有抗凝剂的2 mL EP管中,待采血结束后带回实验室。采用酚氯仿方法提取总DNA,具体如下:①向1.5 mL离心管(EP管)中加入200μL血样。加入500μL SNET(1%SDS,400 mM NaCl,5 mM EDTA,20 mM Tris-cl pH 8.0),混匀。②向以上溶液中加入蛋白酶K至终浓度100 ng/mL,混匀。③55℃孵育4 h。④加入等体积的Tris-饱和酚,缓慢颠倒离心管数次,12 000 r/min,离心10 min。⑤取水相于新的EP管中,加入等体积氯仿/异戊醇(24:1),震荡摇匀10 min,12 000 r/min,离心10 min。⑥取水相于新的EP管中,加入1/10体积的3 M NaAc及2.5倍体积的无水乙醇,摇匀,-20℃静置30 min,12 000 r/min,离心10 min。⑦去水相,干燥后用适量的超纯水溶解。
1.2外显子引物设计从NCBI中查找绵羊TFB2M基因序列(GenBank:NC_019469)作为参考序列,针对所有外显子设计引物,引物信息如表1。TFB2M基因含有8个外显子,每个外显子设计一对引物,其中第3和第4外显子紧邻,因此设计一对引物。
1.3P P C C R R扩增测序PCR反应体系(25μL):ddH2O 17.85μL,10×PCR buf fer 2.5μL,dNTP(10 mM)0.5μL,正向引物(10 mM)1.0μL,反向引物(10 mM)1.0μL,Taq DNA Polymerase(5 U/μL) 0.15μL,DNA 1μL。反应条件:95℃预变性5 min;循环条件95℃变性30 s,退火30~60 s(退火温度见表1),72℃延伸30 s,35个循环,72℃延伸7 min;4℃保存。将PCR产物纯化后直接测序,获得DNA序列信息。
1.4遗传变异分析测序图谱用Chromas 2.22软件进行碱基核查,以胶图文件为准,使用DNAStar软件包中SeqMan软件(DNASTAR Inc,2007)校对原始序列文件中明显的错误位点,根据测序峰图和序列比对获得遗传变异信息。使用DNAMAN 7.0对所有位点进行翻译,分析突变类型。根据同源性以已经解析蛋白结构(PDB ID:4GC5)为模板在SWISS-Model进行了TFB2M的结构建模,然后用Pymol软件进行处理,以Car toon图的形式展示蛋白三维结构,其中的突变位点以Stick形式表示。(Swissmodel:http://swissmodel.expasy.org/interactive;Pymol:ht tp://www.pymol.org/)
5’-UTR转录因子预测网址:ht tp://www. cbrc.jp/research/db/TFSEARCH.html
蛋白质二级结构分析:SOPMA(ht tp://npsapbi.ibcp.f r/)
2 结果与分析
2.1TFB2M多态分析绵羊TFB2M基因由8个外显子组成,其中5’-UTR为158 bp,3’-UTR为550 bp,编码区(CDS)1 185 bp,编码394个氨基酸(图1)。小尾寒羊mtDNA发现编码区错义突变1个(G226A,GCC→ACC)导致76位苏氨酸变为丙氨酸,5’-UTR发现一处颠换(C58G);3’-UTR有3个突变(C98G、A206G和A429T)。
2.2蛋白质二级结构预测分析利用swiss-model在线软件预测比对TFB2M野生型和突变型蛋白质二级结构,共394个氨基酸,野生型和突变型蛋白质二级机构在α-螺旋和无规则卷曲方面存在差异,Thr76Ala导致TFB2M野生型蛋白质α-螺旋比例由36.55%减少为36.04%,无规则卷曲由35.53%增加到36.04%,其他参数没有变化(图2)。

表1 TFB2M外显子扩增引物Table1 Primers for am p lifying exons of TFB2M gene
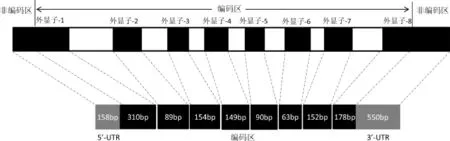
图1 TFB2M基因结构示意图Fig.1 Schem atic structure of TFB2M gene
2.3蛋白质三级结构预测分析错义突变导致晶体结构发生改变,突变位点氨基酸左上方的环和多肽链的羧基端发生改变,可能是由于氨基酸之间的作用发生改变,进而影响构象(图3)。
2.4启动子和55’--U U T T R R转录因子结合位点预测分析采用TFSEARCH生物学软件对小尾寒羊TFB2M基因启动子和5’-UTR转录因子预测,结果显示共有54个转录因子结合位点,5’-UTR 58 bp处没有结合位点,因此该突变并没有改变结合位点(图4)。
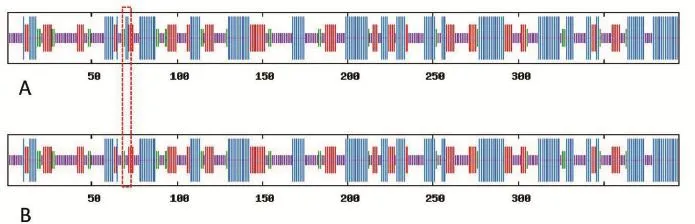
图2 TFB2M编码蛋白二级结构Fig.2 Secondary construction o f TFB2M coding protein
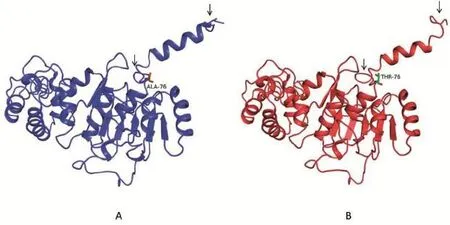
图3 TFB2M基因编码蛋白质晶体结构Fig.3 TFB2M coding protein crysta l structure
3 讨论
线粒体DNA生物合成和维持主要依赖于1 500个核内编码的基因调控,并且这些基因编码的蛋白质在细胞质合成,之后进入线粒体发挥调控作用,并整合与生产、繁殖、代谢和抗病性相关的信号通路。线粒体转录因子主要分为三大类:第一类是直接与mtDNA结合的因子如TFAM、TFB2M、甲状腺激素受体(T3)、RNA聚合酶等;第二类是调控第一类因子的蛋白质,如雌激素受体(ER)、核呼吸因子(NRF-1)等;第三类因子是调控第二类因子的蛋白质,如甲状腺素受体T3。

图4 TFB2M部分启动子和5’-UTR转录因子结合位点预测Fig.4 Binding sites prediction of transcrip tion factor in partia l promotor and 5’-UTR o f TFB2M
TFB2M是线粒体基因表达调控的关键因子,是核基因编码的蛋白质,与TFMA、线粒体RNA聚合酶、TFB1M等组成基本转录装置负责mtDNA转录。本研究的TFB2M基因错义突变导致蛋白质二级结构和三级结构发生改变,进而影响表型性状。基因表达是基因与多种转录因子相互作用的结果,参与基因表达调控的序列往往位于基因的非翻译区(UTR)。其中5’-UTR决定转录水平和方式,5’-UTR的启动子和转录因子结合位点是影响真核生物基因表达的重要调控元件。本研究小尾寒羊TFB2M基因部分启动子和5’-UTR转录因子预测,结果显示共有54个转录因子结合位点,5’-UTR 58 bp处没有结合位点,因此该突变并没有改变结合位点。转录后调控主要包括转录本在细胞内的定位、稳定性和翻译效率,而相关调控元件主要多位于非翻译区,其中以3’-UTR更为重要,3’-UTR包含调控基因转录的终止子、顺式作用元件、多聚腺苷酸化信号等多种重要的功能元件,调节mRNA的稳定性、亚细胞定位和翻译水平,决定某一特定转录本的命运,对基因表达起重要调控作用。本研究的ERα基因3’-UTR野生型和突变型分析比对,结果发现了一些UTR作用元件结合序列变化,但与表型性状无直接关联。
[1]Falkenberg M,M Gaspari,A Rantanen,et al. Mitochondrialt ranscriptionfactorsB1and B2activatet ranscriptionofhumanmtDNA. Nat.Genet,2002,(31):289-294.
[2]McCul loch V,B L Seidel-Rogol,G S Shadel.A humanmitochondrialtranscriptionfactoris relatedtoRNAadeninemethyl t ransferases and binds S-adenosylmethionine.Mol Cel l Biol,2002,22:1116-1125.
[3]Gleyzer N,K Vercauteren,R C Scarpul la.Cont rol of mitochondrial t ranscription speci f icity factors(TFB1M and TFB2M)by nuclear respiratory factors(NRF-1 and NRF-2)and PGC-1 fami ly coactivators.Mol Cel l Biol,2005,25:1354-1366.
[4]Spiegelman B M.Transcriptional cont rol of mitochondrialenergymetabolismthroughthe PGC1coactivators.Novar tisFound.Symp., 2007,287:60-63.
[5]Scarpul la R C.Transcriptional paradigms in mammal ian mitochondrial biogenesis and function.Physiol.Rev,2008:611-638.
[6]Litonin D M,Y Sologub,M Shi,et al.Human mitochondrialt ranscriptionrevisited:only TFAM and TFB2Mare required for t ranscription of the mitochondrial genes in vit ro.J.Biol. Chem,2010,(285):18129-18133.
[7]毕晓丹,储明星,金海国,等.小尾寒羊高繁殖力候选基因ESR的研究[J].遗传学报,2005,32(10):1060-1065.
[8]Eichner L J,V Giguère.Est rogen related receptors(ERRs):A new dawn in t ranscriptional cont rol of mitochondrial gene networks[J].Mitochondrion,2011,11:544-552.
[9]Chen J Q,Delannoy M,Cooke C,et al.Mitochondrial local ization of ERαand ERβin human MCF7 cel ls[J].Am J Physiol Endocrinol Metabol ism,2004,286:E1011-E1022.
Analysis ofgenetic variation of TFB2M for Small-tailed Han sheep
Chen Xiaoyong1,2,Wang Wei1,Dun Weitao2*,Zhao Xingbo1*
(1.Col lege Animal Science& Technology,China Agricul tural University;Bei j ing100193; 2.Institute of Animal Science and Veterinary of Hebei Province,HebeiBaoding071000)
s:In order to discusse genetic variation of TFB2M,the polymorphism in 8 exons,5’-UTR and 3’-UTR were analyzed through gene sequencing of Smal l-tai led Han sheep.The resul ts demonst rated that there were one missense mutations(G226A,GCC→ACC),which lead to the threonine change to alanine,one t ransversion C58G in 5’-UTR,and three substitutions C98G,A206G,A429T in 3’-UTR.Bioinformatics toos was used to predict the promoter and 5’-UTR t ranscription factor,it indicated that 54 transcription factor binding sites were found,however,the substitution(C58G) did not change the binding site.This work provided the molecular biological foundation for the variation and function of TFB2M gene in sheep.
TFB2M;Smal l-tai led Han sheep;Variation;Transcription factor
S826.2
B
1672-9692(2016)02-0015-06
2016-01-04
陈晓勇(1980-),男,河北省滦县人,高级畜牧师,博士,主要研究方向:动物遗传繁育及其生物技术。
敦伟涛(1963-),男,本学,研究员,从事羊遗传繁育与生物技术研究。赵兴波(1969-),男,博士,教授,从事动物分子遗传学与分子育种研究。
转基因生物新品种培育重大专项(2014ZX08009-002),河北省科技支撑计划项目(No.14226315D)和(No.15226308D)。
