肝癌细胞系Hep3B的泛素化蛋白质组学
2016-11-14齐英姿邓晨苏纳张令强徐平
齐英姿,邓晨,苏纳,张令强,徐平
肝癌细胞系Hep3B的泛素化蛋白质组学
齐英姿,邓晨,苏纳,张令强,徐平
军事医学科学院放射与辐射医学研究所蛋白质组学国家重点实验室国家蛋白质科学中心 (北京) 北京蛋白质组研究中心蛋白质药物国家工程研究中心,北京 102206
泛素化修饰是细胞内最重要的翻译后修饰形式之一,对细胞内蛋白质的稳定、降解、定位以及生物活性的调节起到重要作用。但因其在细胞内丰度低、降解周期短等特点而很难被检测。本研究中,制备的泛素结合结构域蛋白 (Ubiquitin-binding domains,UBDs) 用于富集肝癌细胞系Hep3B中的泛素化蛋白,并通过液相色谱-串联质谱联用的方法对富集的泛素化蛋白进行鉴定。实验共鉴定到1 900个潜在的泛素化蛋白和158个泛素化位点,这些被鉴定到的泛素化位点分属于102个蛋白。生物信息学分析发现泛素化蛋白显著富集的相关通路与肿瘤的发生发展密切相关,此结果暗示肿瘤细胞内泛素化-蛋白酶体的失调与肿瘤细胞的信号传导及细胞外基质的变化等具有较高的关联性。
Hep3B,蛋白质组学,泛素化,ThUBDs
蛋白质是组成细胞的基石,是细胞生命活动的最终执行者。机体的正常功能的行使依赖于细胞内众多蛋白质有条不紊地发挥着多种多样的功能。真核细胞内普遍存在着复杂的蛋白质质量控制系统,而正是这种质量控制系统的存在,细胞内的蛋白质才能够在复杂的体内环境中生存,并正确地行使其功能。而在细胞内这些不同层次的蛋白质质量控制体系中,蛋白质翻译后修饰被认为是最为快速、高效、经济的调控方式,可迅速地调控细胞进程[1-4]。蛋白质泛素化修饰是调控蛋白质质量的最重要的翻译后修饰形式。细胞内的泛素化修饰主要参与泛素-蛋白酶体途径,执行对细胞内蛋白有序降解的功能,以此来保证翻译过程中及翻译后的折叠、装配出现错误的或有害的蛋白质被降解,避免对细胞产生毒害作用。泛素-蛋白酶体途径是对蛋白质具有高度选择特异性的降解过程,而蛋白质的降解又可调控其他细胞事件的进行,从而对一系列的细胞行为产生影响。
以泛素-蛋白酶体系统为代表的蛋白质质量控制系统失调,会导致一系列疾病,如阿尔茨海默症等神经退行性疾病以及肿瘤的发生等[5]。在肿瘤发生过程中,这种调节机制的丧失会导致如抑癌蛋白的过量降解或癌蛋白降解途径失效等,都会导致癌症的发生。泛素蛋白酶体系统对p53、p27、分子伴侣、26S蛋白酶体等的调控均可影响肿瘤的发展进程[6-9]。肝细胞癌是严重威胁我国国计民生的重大疾病。世界范围内,肝细胞癌居于男性患者癌症发病率第5位,死亡率第2位[10]。其显著的诱因包括乙型肝炎病毒 (HBV) 的感染而引发的慢性肝炎。我国是HBV的高发区,因此HBV感染引发的细胞内泛素蛋白酶体的变化及其肝细胞癌病机制也是基础医学研究应当关注的重点。已有研究表明,HBV病毒中的一个独特开放读码框所编码的HBx蛋白可与蛋白酶体的亚基相互作用,作为转录辅助激活因子发挥其功能[11]。此功能可能是HBV作用于受体细胞的分子基础之一。还有研究显示HBx蛋白中有一段保守的肽链可下调p97表达量而降低蛋白酶体的活性,引起HepG2细胞的增殖减缓和凋亡[12],这也为针对HBV引起的肝细胞癌的蛋白酶体抑制剂药物的研制带来希望。
与细胞内大部分翻译后修饰一样,泛素化修饰也同样具有丰度低、动态范围大、降解快速及结构较为复杂等问题,使得对其大规模鉴定和研究成为挑战。传统的泛素化富集方法主要通过在泛素链的末端引入亲和标签的技术来实现的。如Gygi实验室[13]用His标签对泛素进行标记,首次实现了质谱技术对泛素化蛋白及位点的鉴定。随后又发展了HB标签、Flag标签标记的方法等对细胞系、果蝇等样品中泛素化蛋白及位点的鉴定[14-16]。但是,该方法应用的样本有限,尤其是不能应用于组织样本或病理标本,使得其在医学研究范畴的应用受到限制。此外,该方法还存在对泛素化位点的富集具有一定的选择性,可能干扰细胞的正常功能,无法表征细胞内真正的泛素化指标等缺点。而后,高亲和抗体的方法也成功应用于泛素化位点的富集[17-18];该方法是基于泛素化蛋白经胰酶酶解后会产生两个甘氨酸残基残留的特点进行富集,优点是富集的泛素化位点数及蛋白数较多,缺点是抗体高背景所带来的非特异的污染蛋白多。而泛素结合结构域(Ubiqutin-binding domains,UBDs) 也是可以识别和结合泛素化修饰的小蛋白,是进行泛素化富集的有效工具之一。本实验室自主开发了串联杂合UBD (Tandem hybrid UBD,ThUBDs) 的方法对生物样本中的泛素化蛋白进行无偏向性的富集,该方法应用范围广,可应用于各种细胞系、动物组织及病理样本的泛素化富集研究[19]。
本实验利用ThUBDs富集泛素化蛋白与质谱鉴定蛋白翻译后修饰相结合的方法,对肝癌细胞系Hep3B中的泛素化蛋白进行富集和LC-MS/MS鉴定,我们共鉴定到1 900个潜在的泛素化蛋白和158个分属于102个蛋白质的泛素化位点。对1 900个潜在的泛素化蛋白进行生物信息学分析发现,这些蛋白主要聚类于细胞内或细胞间信号转导通路、转移侵袭相关的胞间连接以及蛋白泛素化信号通路等,提示肿瘤细胞内的泛素化-蛋白酶体的失调可能与细胞的癌症特征的出现密切相关。
1 材料与方法
1.1 材料
1.1.1 细胞系
肝癌细胞系Hep3B购于中国医学科学院基础医学研究所北京协和细胞资源中心。细胞保存于含有体积比10%二甲基亚砜 (Dimethyl sulfoxide,DMSO) 以及50%胎牛血清的DMEM培养基中,置于液氮中保存。
1.1.2 小鼠品系
Balb/c小鼠,雌性,SPF级,购于北京维通利华实验动物技术有限公司。按照实验动物的3R原则给予人道关怀。
1.2 方法
1.2.1 细胞复苏、培养和传代
将细胞冻存管从液氮中取出,迅速置于37 ℃水浴锅中。待冻存管中的液体融化后,液体转移到15 mL离心管中,加入4 mL培养基,常温离心 (1 000 r/min,5 min)。弃上清,保留沉淀4 mL培养基将细胞沉淀重悬,置于37 ℃、5% CO2细胞恒温培养箱中培养。
待贴壁生长的细胞达到80%−90%汇合度时,加入0.25%胰蛋白酶消化,待细胞形态开始收缩变形时,弃掉胰蛋白酶液,加入培养基重悬细胞,按1∶2比例传代接种于培养瓶中,置于饱和湿度37 ℃、5% CO2细胞恒温培养箱中培养。
1.2.2 细胞超声破碎提取蛋白
收集的细胞沉淀加入3 mL裂解液 (PBS,10%甘油、1 mmol/L EDTA、0.5% 乙基苯基聚乙二醇 (Nonidet P 40,NP-40)、1×N-乙基马来酰亚胺 (N-ethylmaleimide,NEM)、5 mmol/L IAA、1×蛋白酶体抑制剂cocktail,1×苯甲基磺酰氟 (Phenylmethanesulfonyl fluoride (PMSF),pH 7.4),吹打重悬后,置于超声破碎仪中破碎 (功率30%,超声2 s,间歇4 s,超声10 min)。超声后蛋白溶液于4 ℃、13 300 r/min离心 10 min。收集上清用0.45 μm的滤器过滤,置于冰上备用。
1.2.3 融合蛋白GST-UBD的纯化及固定化耦联
表达GST-UBD融合蛋白的BL21 (DE3) 工程菌株由本实验室构建和保存,在此基础上进行融合蛋白GST-UBD的诱导、纯化及固定化耦联。
1.2.4 融合蛋白GST-UBD的诱导
将冻存于–80 ℃的菌种划线接种于氨苄青霉素 (100 μg/mL) 抗性的选择平板上,37 ℃条件下培养12 h获得单菌落。挑取单菌落接种于 5 mL液体LB培养基 (100 μg/mL) 的氨苄青霉素),37 ℃振荡培养12 h。取培养菌液1%接种到3 800 mL液体摇瓶中,37 ℃、220 r/min振荡培养3 h (此时菌体600约为 0.6)。添加终浓度为0.2 mmol/L的IPTG,37 ℃、220 r/min继续振荡培养5 h。4 500×离心10 min收集菌体,收集的菌体用PBS重悬洗涤1次,6 000×离心10 min收集菌体。用适量PBS重悬菌体 (至菌体600约为40 mL),低温超声 (超声工作2 s,间歇4 s,180 W,超声20 min) 破碎细胞。超声完成后,4 ℃、8 000×离心20 min,收集上清,用于目的蛋白的纯化。
1.2.5 融合蛋白GST-UBD的纯化
使用AKTA Purifier 100进行蛋白的纯化,纯化介质为5 mL预装GST-resin柱。将GST-resin柱装到合适的柱位,10个柱体积的20%乙醇清洗系统后再以20个柱体积的ddH2O清洗系统;用PBS对系统进行平衡,至UV值不变,调零。2 mL/min上样,至上样完全。上样结束后,用PBS进行洗涤,至UV值不变。用含有50 mmol/L还原型谷胱甘肽的PBS进行目的蛋白的洗脱,并收集有吸光值处的流分,即为所纯化蛋白质溶液。
1.2.6 融合蛋白GST-UBD的固定化耦联
使用超滤管 (分子截留量为30 kDa) 对纯化所得的蛋白溶液进行耦联缓冲液(0.2 mol/L NaHCO3,0.5 mol/L NaCl,pH 8.3) 的置换和蛋白浓缩,每次10 000×、3 min,共5次。15个柱体积的4 ℃预冷的1 mmol/L HCl洗涤平衡琼脂糖珠。将浓缩后的融合蛋白与平衡好的琼脂糖珠进行混合,于4 ℃翻转孵育24 h。耦联结束后,将混合物于4 ℃、200×离心3 min,弃上清液,加入2倍柱体积0.1 mol/L Tris-HCl (pH 8.5),于4 ℃翻转孵育4 h。用2倍柱体积30%甘油的PBS洗涤,4 ℃、200×离心3 min,弃上清,共3次。洗涤完毕冻存于–20 ℃备用。
1.2.7 蛋白质组样本制备
样本制备参考Zhai等[25]的方法进行。取 10 μL蛋白溶液,加入等体积2×SDS上样缓冲液,均匀混合后,使用10%的SDS-PAGE胶进行电泳。待溴酚蓝指示剂距离分离胶与浓缩胶交界面约1 cm处时停止电泳。凝胶经考马斯亮蓝G250染色、脱色后,将胶条均分成1 mm3的胶粒,然后进行脱色、干燥步骤。所得胶粒使用Trypsin进行酶解16 h后,抽提肽段,蒸干后于–80 ℃保存备用。
1.2.8 LC-MS/MS分析
使用LTQ QrbitrapVelos质谱仪 (Thermo Fisher Scientific,Waltham,MA,USA) 与超高压液相色谱系统(Waters Corporation,Milford,MA,USA) 联用对肽段样品进行检测。使用自制的反相色谱柱 (75 μm I.D.×15 cm,3 μm,200A) 进行梯度洗脱。样品经自动进样器加载于色谱柱上,通过100 min的液相梯度进行分离,其中流动相A为0.1%甲酸 (Formic acid,FA),2%乙腈 (Acetonitrile,AcN),98% H2O,流动相B为0.1% FA,90% AcN,10% H2O;流速为 300 nL/min。洗脱后的肽段经高精度的LTQ-OribitrapVelos质谱仪进行分析,喷雾电压2 kV,扫描范围为300–1 600/;自动增益1×106,最大离子注入时间为150 ms,一级谱精度30 000。质谱采用一级谱数据依赖的二级谱扫描模式 (Data Dependent MS/MS Scan) 碰撞诱导裂解 (CID) 模式碎裂一级离子。选择20个信号最强的母离子肽段进行碎裂,35%归一化碰撞能量,动态排除设置为30 s。
1.2.9 质谱数据搜库鉴定
质谱产生的谱图使用MaxQuant (version 1.5.3.30) 软件进行数据搜索,人蛋白质序列库下载自UniProt (http://www.uniprot.org/,release2015.11)。使用Target-Decoy策略进行搜库,控制肽段和蛋白水平FDR<1%。搜库参数设置为:允许最大2个漏切位点,允许半酶切,每个肽段允许长度为6−20个氨基酸;母离子质量误差 20 ppm,子离子质量误差0.1 Da;固定修饰为半胱氨酸烷基化修饰 (C+57.021 5 Da),可变修饰为甲硫氨酸氧化修饰 (M+15.994 9 Da) 和GlyGly (K+114.042 9 Da)。
1.2.10 生物信息学分析
使用信号网络分析软件 (Ingenuity pathway analysis,IPA) 进行疾病与功能的生物信息学分析。
2 结果与分析
2.1 融合蛋白GST-UBD纯化结果
采用亲和层析和凝胶过滤层析相结合的两步纯化方法对目的蛋白进行纯化。图1A为亲和层析时280 nm处吸光值的检测曲线,可以看出有明显的洗脱峰出现。对过程中的各阶段留样进行的SDS-PAGE检测,结果如图1B所示,可见50−60 kDa分子量之间有明显的蛋白条带,与理论分子量大小 (54.2 kDa) 一致,判断为目的蛋白。为了除去目的条带之外的杂带,进行凝胶过滤层析实验,检测曲线如图1C所示,可以看出有不同的洗脱峰。对不同的组分进行SDS-PAGE检测,结果如图1D所示,与凝胶过滤前的洗脱组分相比,第5、6、7洗脱组分的杂带有所减少,而之后组分中的杂蛋白又开始增加,因此取5、6、7洗脱组分进行浓缩、置换和固定化耦联。
2.2 融合蛋白GST-UBD固定化耦联beads的承载能力检测结果
首先利用正常小鼠Balb/c品系的肝脏组织,进行融合蛋白GST-UBD固定化耦联beads的承载能力的测试实验。提取出的全蛋白溶液通过灰度值方法进行定量[21],结果显示,鼠肝全蛋白溶液浓度为4.5 μg/μL (图2A)。将全蛋白裂解溶液 (Total cell lysate,TCL) 设置为6个梯度,分别是20、50、100、200、800、1 500 μL,分别与20 μL GST-UBD固定化耦联beads在4 ℃条件下进行旋转孵育2 h,纯化的UBC蛋白通过银染方法进行检测。结果显示,TCL梯度 200 μL前富集到的UBC蛋白量逐渐增加,而梯度200 μL后的TCL中富集到的UBC蛋白量基本持平 (图2B)。利用灰度值定量的方法对银染结果进行定量,后对20 μL GST-UBD固定化耦联beads可结合的UBC蛋白的得量作图 (图2C),发现在200 μL的TCL附近出现拐点。计算可得纯化并偶联所得的UBDs-beads,每20 μL最大可与900 μg全蛋白进行反应,结果表明UBDs-beads的UBC结合能力强。以该beads与TCL结合比例进行后续的肝癌细胞系Hep3B中的泛素化蛋白富集实验。
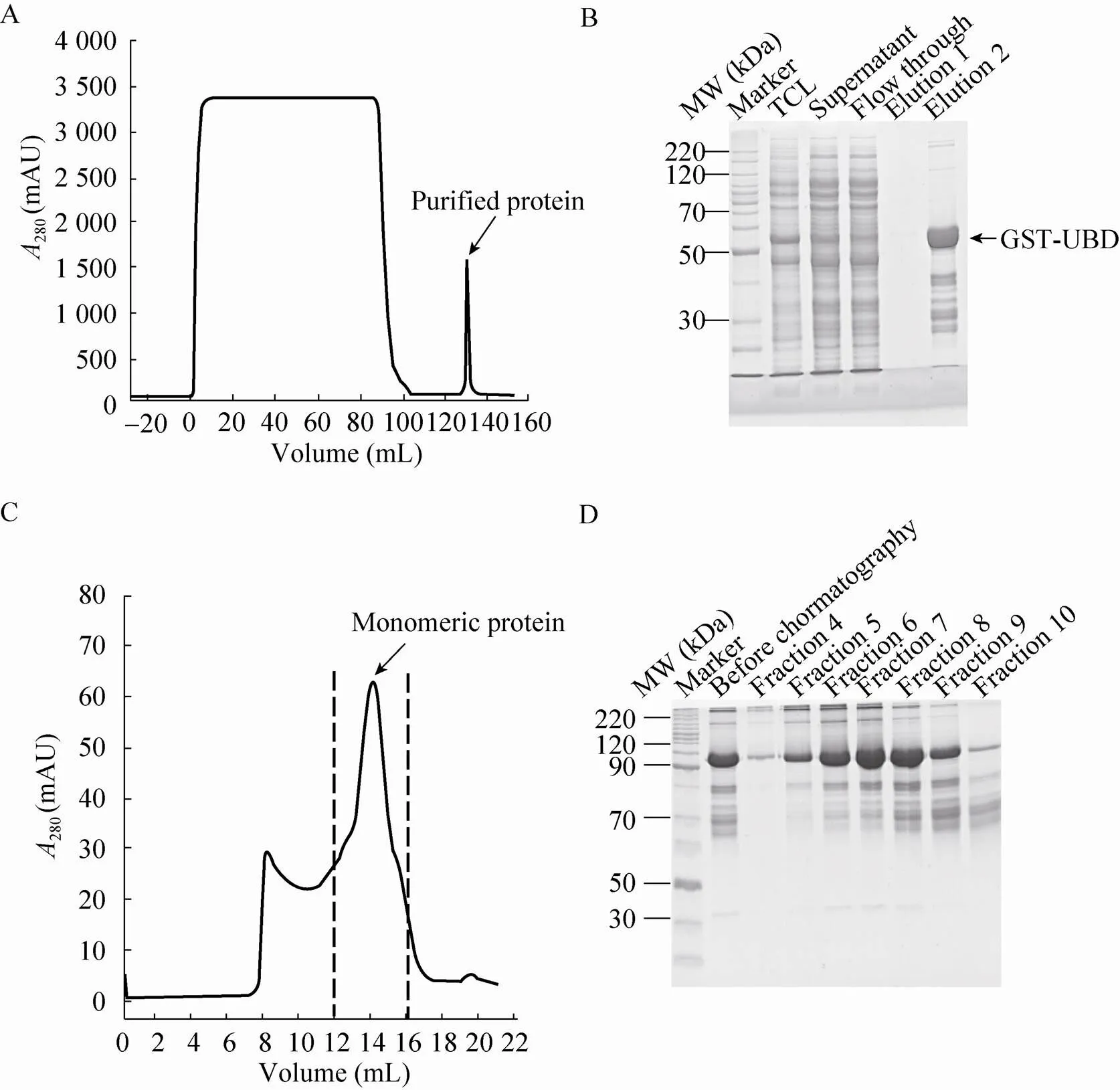
图1 融合蛋白GST-UBD纯化结果
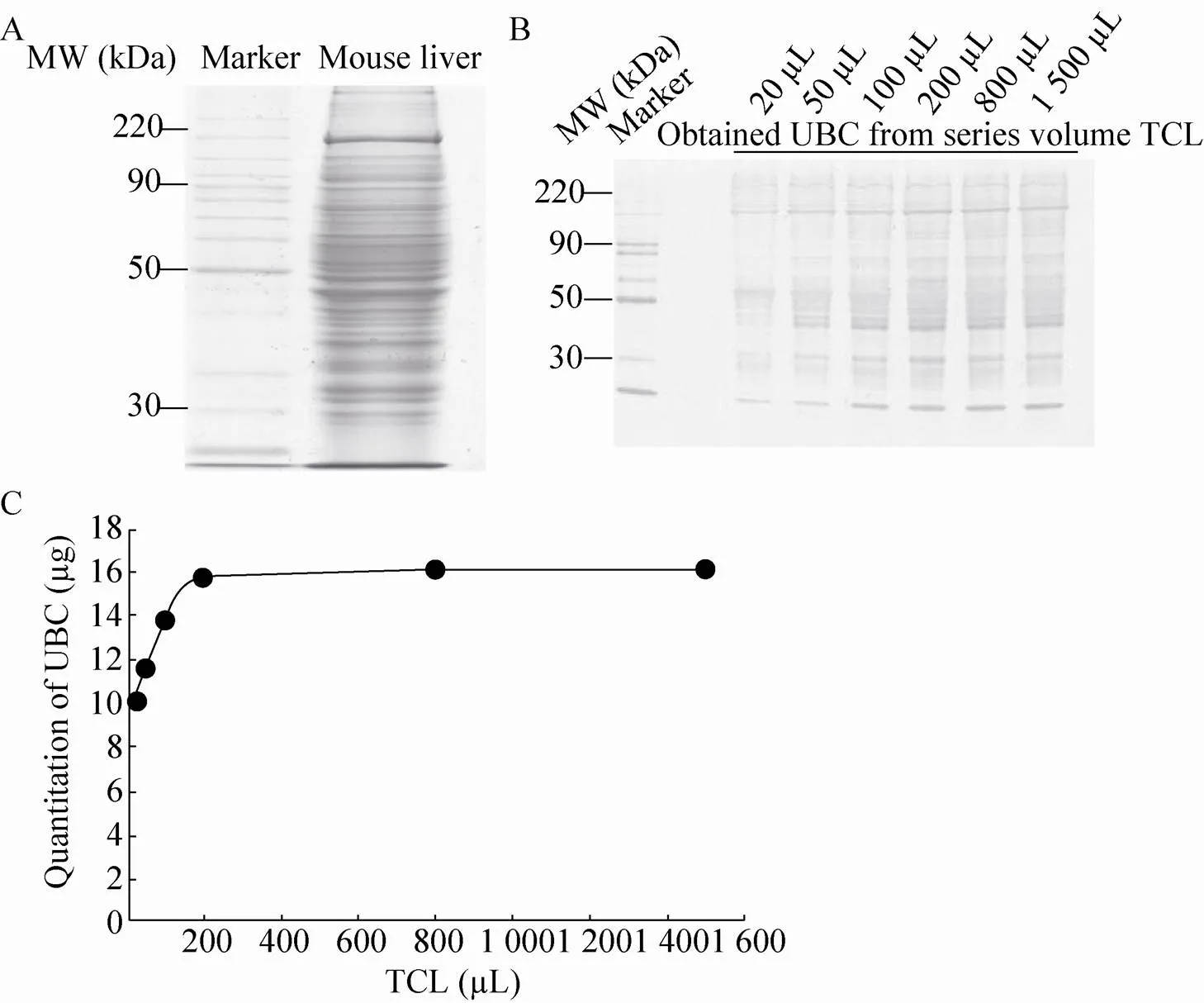
图2 融合蛋白GST-UBD固定化耦联beads的承载能力检测结果
2.3 肝癌Hep3B细胞系泛素化富集实验结果
Hep3B细胞系是Aden等从一名8岁的美国男性黑人肝癌患者的活体标本分离出来的肝癌细胞系,具有合成多种人血浆蛋白的能力,是研究原发性肝癌的重要的细胞系模型。故我们选取Hep3B细胞系作为实验材料,研究细胞内泛素化-蛋白酶体系统的失调情况,探索肿瘤细胞内蛋白质质量控制系统的调控机制。
泛素化富集实验流程如图3A所示。我们收集了4×107的Hep3B细胞进行泛素化富集实验 (图3B)。提取细胞总蛋白并进行SDS-PAGE分离,结果如图3C所示,细胞破碎裂解充分,无降解等情况发生。同样根据灰度值定量算法,Hep3B细胞系的全蛋白裂解液浓度为1.2 μg/μL (图3C),根据计算,我们共投入80 μL的UBDs-beads以保证泛素化蛋白的富集效果。

图3 肝癌Hep3B细胞系泛素化富集实验结果
2.4 Hep3B细胞系UBC蛋白质谱检测结果
所得全蛋白溶液在4 ℃条件下与UBD-Besds进行结合,富集细胞内的UBC蛋白。为检测富集效果,富集得到的UBC蛋白进行SDS-PAGE胶分离后,使用银染的方法进行检测,结果如
图4A所示。所得UBC蛋白条带与富集前全细胞裂解液 (图3C) 相比,条带带型发生了明显的变化,大分子量蛋白增多,富集效果显著。为进一步确证UBC蛋白的富集效果并检测富集的UBC蛋白的组成情况,我们使用质谱技术和非标记定量的方法对Hep3B细胞系中富集得到的蛋白进行检测 (SDS-PAGE上样胶图4B)。如图4C所示,在保留时间25 min左右时,S57肽段集中出峰,且具有较高的峰容量,说明UBC实验富集效果显著。

图4 Hep3B细胞系UBC蛋白质谱检测结果
2.5 质谱搜库数据及生物信息学分析结果
通过MaxQuant软件进行搜库,总鉴定谱图数110 196,总鉴定肽段数26 579,MS2鉴定率24.12%,鉴定到潜在的泛素化蛋白1 900个,鉴定到的泛素化位点158个,分属于102个蛋白。
我们对1 900个潜在的泛素化蛋白通过IPA软件进行生物信息学分析。经典通路分析结果显示,聚类排名前列的通路分别为EIF2信号通路、EIF4和p70S6K信号调节通路、蛋白泛素化信号通路、mTOR信号通路、氨酰tRNA合成途径、上皮细胞紧密连接重塑途径、生精细胞-支持细胞连接通路和线粒体失调通路等 (图5)。这些通路均与肿瘤的发生发展密切相关,提示肿瘤细胞内的泛素化-蛋白酶体的失调与肿瘤细胞的信号传导及细胞外基质的胞间连接的失调等具有很高的关联性,从而导致了如细胞凋亡受阻或细胞永生化现象的出现以及肿瘤形态或癌症的出现及异常。
3 讨论
富集技术的发展是翻译后修饰组学发展的限速步骤,翻译后修饰由于存在丰度低、变化快速等问题,从而制约着对其的鉴定和研究。泛素化修饰同样面临着这些问题,它在细胞内丰度极低且可被26S蛋白酶体降解使其难以被质谱鉴定到。本研究中,利用串联UBDs的方法对细胞内的泛素化蛋白进行无偏向性的富集。我们以肝癌细胞系Hep3B作为实验材料,对细胞内的泛素化蛋白进行富集,并通过LC-MS/MS进行鉴定。我们共鉴定到1 900个潜在的泛素化蛋白和158个泛素化位点,这些被鉴定到的泛素化位点分属于102个蛋白。对 1 900个潜在的泛素化蛋白进行生物信息学分析发现,这些蛋白主要聚类于细胞内或细胞间信号转导通路、转移侵袭相关的胞间连接以及蛋白泛素化信号通路等,提示肿瘤细胞内的泛素化-蛋白酶体的失调可能与细胞的癌症特征的出现密切相关。

图5 IPA经典通路分析结果
与其他的泛素化富集的方法相比,串联UBDs的方法不需要泛素的工程化过表达,使其更适合于哺乳动物细胞或临床病理组织等样本的泛素化富集实验。在本实验室前期的工作中,我们利用串联UBDs的方法,尝试富集了肝癌细胞系MHCC-97H细胞中的泛素化蛋白[24]。MHCC-97H肝癌细胞系是由中国复旦大学肝癌研究所的研究团队所建立的具有自发性转移潜能的高转移性单克隆细胞株人肝癌细胞系[27-28]。比较Hep3B和MHCC-97H的泛素化蛋白,我们发现MHCC-97H细胞所聚类的信号通路更多的富集于网格蛋白所介导的内吞作用的信号通路、RhoGDI通路、Rho家族GTP酶信号通路、14-3-3蛋白介导的通路和肌动蛋白细胞骨架信号通路等。这些通路基本都与细胞骨架的变化或细胞的侵袭与转移功能密切相关(图6),表明肿瘤细胞内泛素化蛋白酶体的失调与肿瘤本身所表现出来的癌症特性关系密切。我们对细胞内泛素化蛋白进行高效富集和LC-MS/MS分析,可以准确地表征细胞内泛素化蛋白酶体系统失调的情况,可为肿瘤特征表型蛋白质泛素化信号通路研究提供技术支持[24]。
肝细胞癌是威胁我国国计民生发展的重大疾病之一。当前,肝癌的临床诊疗方法所面临的主要的困难是肝癌的易转移、易复发的难题。作为高异质性的癌症,肝细胞癌本身复杂性极高,其本身的复杂性和长病程等特点给我们的研究工作带来诸多困难。如前文所述,肝癌的癌细胞特征如抗凋亡、特别是侵袭与转移等特征可能与细胞内泛素化蛋白酶体系统的失调有紧密的联系,而本文也提供了一种可以应用于哺乳动物细胞,特别是肿瘤细胞的泛素化富集的策略,为研究肝细胞癌在发生发展的病程变化过程中的细胞内泛素化蛋白酶体的连续变化过程提供了技术支撑。接下来对于肝细胞癌在发生发展病程中泛素化蛋白酶体失调规律的深入探讨,是揭示肝癌发生发展过程分子机制的热点方向。

图6 MHCC-97H细胞系UBC蛋白IPA经典通路分析结果[24]
REFERENCES:
[1] Kerscher O, Felberbaum R, Hochstrasser M. Modification of proteins by ubiquitin and ubiquitin-like proteins. AnnuRevCell DevBiol, 2006, 22(1): 159–180.
[2] Schwartz AL, CiechanoverA. The ubiquitin- proteasome pathway and pathogenesis of human diseases. Annu Rev Med, 1999, 50(1): 57–74.
[3] Mukhopadhyay D, Riezman H. Proteasome- independent functions of ubiquitin in endocytosis and signaling. Science, 2007, 315(5809): 201–205.
[4] Haglund K, Dikic I. Ubiquitylation and cell signaling. EMBO J, 2005, 24(19): 3353–3359.
[5] Ciechanover A. The ubiquitin-proteolytic system: from bench to bedside. Growth Horm IGF Res, 2008, 18(S1): S10.
[6] Chen F, ChangD, GohM, et al. Role of p53 in cell cycle regulation and apoptosis following exposure to proteasome inhibitors. Cell Growth Differ, 2000, 11(5): 239–246.
[7] Ljungman M. Dial 9-1-1 for p53: mechanisms of p53 activation by cellular stress. Neoplasia, 2000, 2(3): 208–225.
[8] Baldassarre G, Belletti B, Bruni P, et al. Overexpressed cyclin D3 contributes to retaining the growth inhibitor p27 in the cytoplasm of thyroid tumor cells. J Clin Invest, 1999, 104(7): 865–874.
[9] Petroski MD. The ubiquitin system, disease, and drug discovery. BMC Biochem, 2008, 9(S1): S7.
[10] Jemal A, BrayF, CenterMM, et al. Global cancer statistics. CA Cancer J Clin, 2011, 61(2): 69–90.
[11] Zhang ZS, ToriiN, FurusakaA, et al. Structural and functional characterization of interaction between hepatitis B virus X protein and the proteasome complex. J BiolChem, 2000, 275(20): 15157–15165.
[12] Liu H, Ye L, Wang QW, et al. Effect of a conserved peptide derived from Kunitz domain of hepatitis B virus x protein on the cell cycle and apoptosis of HepG2 cellsthe proteasome pathway. Chin Med J, 2009, 122(4): 460–465.
[13] Peng JM, Schwartz D, Elias JE, et al. A proteomics approach to understanding protein ubiquitination. Nat Biotechnol, 2003, 21(8): 921–926.
[14] Meierhofer D, Wang XR, Huang L, et al. Quantitative analysis of global ubiquitination in HeLa cells by mass spectrometry. J Proteome Res, 2008, 7(10): 4566–4576.
[15] Franco M, Seyfried NT, Brand AH, et al. A novel strategy to isolate ubiquitin conjugates reveals wide role for ubiquitination during neural development. Mol Cell Proteomics, 2011, 10(5): M110.002188.
[16] Tagwerker C, Flick K, Cui M, et al. A tandem affinity tag for two-step purification under fully denaturing conditions: application in ubiquitin profiling and protein complex identification combined withcross-linking. Mol Cell Proteomics, 2006, 5(4): 737–748.
[17] Vasilescu J, Smith JC, Ethier M, et al. Proteomic analysis of ubiquitinated proteins from human MCF-7 breast cancer cells by immunoaffinity purification and mass spectrometry. J Proteome Res, 2005, 4(6): 2192–2200.
[18] Kim W, Bennett EJ, Huttlin EL, et al. Systematic and quantitative assessment of the ubiquitin- modified proteome. Mol Cell, 2011, 44(2): 325–340.
[19] Gao Y, Li YC, Zhang CP, et al. Enhanced purification of ubiquitinated proteins by engineered tandem hybrid ubiquitin-binding domains (ThUBDs). Mol Cell Proteomics, 2016: mcp.O115.051839.
[20] Zhai LH, Chang C, Li N, et al. Systematic research on the pretreatment of peptides for quantitative proteomics using a C18microcolumn. Proteomics, 2013, 13(15): 2229–2237.
[21] Ma SZ, Zhang T, Zhai LH, et al. Short gel method for pretreatment of protein samples with high concentration of detergent. Chin J Biotech, 2014, 30(9): 1446–1453 (in Chinese). 马首智, 张涛, 翟琳辉, 等. 短胶法预处理含高浓度去垢剂的蛋白样品. 生物工程学报, 2014, 30(9): 1446–1453.
[22] Tian J, TangZY, Ye SL, et al. New human hepatocellular carcinoma (HCC) cell line with highly metastatic potential (MHCC97) and its expressions of the factors associated with metastasis. Br J Cancer, 1999, 81(5): 814–821.
[23] Li Y. Optimization of ametastaticcell model of human hepatocellular carcinoma and study on the mechanismsof metastasis [D]. Shanghai: Fudan University, 2002(in Chinese). 李雁. 转移性人肝癌细胞模型的优化及转移机理探讨[D]. 上海: 复旦大学, 2002.
(本文责编 陈宏宇)
Ubiquitinated proteomics research of Hep3B
Yingzi Qi, Chen Deng, Na Su, Lingqiang Zhang, and Ping Xu
Beijing Institute of Radiation Medicine, State Key Laboratory of Proteomics, National Center for Protein Sciences Beijing, Beijing Proteome Research Center, National Engineering Research Center for Protein Drugs, Beijing 102206, China
Ubiquitination is one of the most major post-translational modifications playing important role in regulation of intra-cellular proteins’ stability, degradation, localization and biological activity. However, these proteins are difficult to be detected due to their low abundance, short half-life. In this study, ubiquitin-binding domains (UBDs) were constructed to purify the ubiquitinated proteins from Hep3B cells. Ubiquitinated proteins and sites were detected by LC-MS/MS. A total of 1 900 potential ubiquitinated proteins were identified. Among them, 158 ubiquitinated sites were identified, belonging to 102 proteins. Bioinformatics analysis revealed that the enriched pathways of ubiquitinated proteins were closely related to tumor occurrence and development. The dysfunction of ubiquitin-proteasome has a high correlation with cell signaling and extracellular matrix changing in tumor cells.
Hep3B, proteomics, ubiquitination, ThUBDs
April 8, 2016; Accepted: May 26, 2016
s: Lingqiang Zhang. Tel: +86-10-68177417; Fax: +86-10-68177417; E-mail: zhanglq@nic.bmi.ac.cn
Ping Xu. Tel: +86-10-61777113; Fax: +86-10-61777050; E-mail: xuping@mail.ncpsb.org
时间:2016-08-05
http://www.cnki.net/kcms/detail/11.1998.Q.20160805.1009.001.html
Supported by:National Basic Research Program of China (973 Program) (Nos. 2011CB910600, 2013CB911200), National High Technology Research and Development Program of China (863 Program) (Nos. SS2012AA020502, 2011AA02A114), National Natural Science Foundation of China (Nos. 31070673, 31170780), Key Projects in the National Science & Technology Pillar Program (No. 2012BAF14B00).
国家重点基础研究发展计划(973计划) (Nos. 2011CB910600,2013CB911200),国家高技术研究发展计划(863计划) (Nos. SS2012AA020502,2011AA02A114),国家自然科学基金 (Nos. 31070673,31170780),国家科技支撑计划 (No. 2012BAF14B00) 资助。
