基于短时程突触可塑性的随机Hodgkin-Huxely神经元网络
2016-09-29周家程丁少杰张雪娟
周家程, 丁少杰, 张雪娟
(浙江师范大学 数理与信息工程学院,浙江 金华 321004)
基于短时程突触可塑性的随机Hodgkin-Huxely神经元网络
周家程,丁少杰,张雪娟
(浙江师范大学 数理与信息工程学院,浙江 金华321004)
根据神经元囊泡释放与离子通道随机切换的生理学机制,给出一个基于短时程突触可塑性的随机HH神经元网络模型,并讨论了通道噪声与突触噪声对网络发放行为的效应,以及易化与抑制对突触效能的影响.
突触可塑性;突触易化;突触抑制;网络模型;HH神经元
0 引 言
神经元的突触可塑性(synapse plasticity)一般被认为是大脑学习与记忆的分子生物学机制,它是指突触传递效率增强或减弱的变化现象.若这种变化只持续数十毫秒到几分,便称之为短时程突触可塑性(short-term synaptic plasticity).其中:效率增强与减弱分别叫作短时程增强(short-term enhancement)与短时程抑制(short-term depression);而持续数百毫秒的增强又被称为短时程易化(short-term facilitation)[1].
易化与抑制是2种并存的机制,它们的共同作用对神经系统的信息传递起到至关重要的作用.因此,研究短时程突触可塑性具有重要意义.1998年,Tsodyks等[2]提出了包含易化和抑制2种机制的突触传递模型,构建简化的神经元网络,并进行动力学分析;2008年,Mongillo等[3]在上述模型的基础上构造了一个复杂的Integrate-and-fire神经元网络,提出了具有工作记忆性的突触理论;2015年,Romani等[4]又在此基础上提出了基于短时程突触可塑性的位置细胞(place cell)网络模型.
神经元在活动中总受到噪声的影响.噪声的来源广泛, 除了从外界环境信号输入噪声外,还包括神经元的通道噪声(channel noise)及突触噪声(synaptic noise)等系统内部存在的噪声.通道噪声作为一类重要的噪声,主要是由离子通道开关的随机性而产生的[5],常常被用来解释某些神经元的自发发放现象[6].这使得经典的确定性Hodgkin-Huxely神经元(简称HH神经元)模型[7]发展成为由随机混合系统(stochastic hybrid system)表示的随机HH神经元模型[8-10].然而,目前并没有较完善的数值模拟方法和定性分析理论来处理这个随机混合系统.因而,往往需要将其Langevin近似成一个Gauss噪声驱动的随机微分方程组[11-14]以减小问题的复杂性.神经元的自发发放与突触可塑性一样,均是生物体内的自组织现象.通过建立数学模型的方法来研究这些现象,可以更清楚地理解这些生物机制,不仅在生理学和医学领域中对阐明相关大脑疾病(如癫痫、老年痴呆症、药物成瘾等)的机理有重要意义,而且在人工智能领域中对利用这些自组织现象进行仿生研究提供理论指导.
事实上,文献[3]提出的神经元网络过于简化,未像文献[15]用α电导来刻画突触噪声,也未像文献[16-17]考虑通道噪声与突触噪声的共同作用,而文献[18]提出的短时程突触可塑性神经元网络中的α电导只包含抑制机制.因此,本文构建了一个通道噪声和突触噪声共同作用下的HH神经元网络,并且突触噪声基于α电导并同时具有易化和抑制2种短时程突触可塑性机制.
1 模 型
考虑由N个兴奋型HH神经元全连接组成的网络.根据Kirchhoff电流守恒律得到每个神经元的膜电位的演化方程为
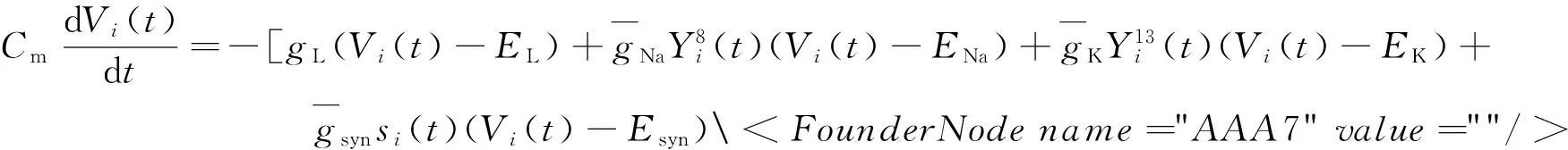
(1)

钠通道和钾通道均为电压门控通道.每个钠通道由3个相同的m门和1个h门组成,从而具有8种状态和20种反应,即
(2)
式(2)中,mphq表示只有p个m门和q个h门打开的状态(p=0,1,2,3,q=0,1).类似地,每个钾通道由4个n门组成,从而具有5种状态和8种反应,即
(3)
式(3)中,np表示有p个n门打开的状态(p=0,1,2,3,4).式(2)和式(3)中的各个状态和各个反应分别被标记为Sl和Rk(l=1,2,…,13,k=1,2,…,28).当一个离子通道的所有门都打开时,这个通道处于开放状态.这样,m3h1(S8)和n4(S13)分别对应钠通道和钾通道的开放状态.需要指出的是,式(2)和式(3)中的反应速率常数{αj,βj}(j=m,n,h)依赖于对应神经元的膜电位,其形式在文献[7]中给出.定义Yi(t)=(Y1i(t),Y2i(t),…,Y13i(t))T,其中Yli(t)为状态Sl的通道开放比例,其形式为
其中:Xli(t)为t时刻第i个神经元处于状态Sl的通道数; NNa=ρNa×S,NK=ρK×S分别是钠通道和钾通道的通道总数(ρNa,ρK为相应的通道密度,S为膜面积).易见Yi(t)∈SY,其中
由随机HH神经元的Langevin近似理论[11-14]可知,Yi(t)的演化可以由如下随机微分方程近似表示:
(4)
式(4)中,υk为反应Rk的状态改变向量,满足
ξki(t)(i=1,2,…,N,k=1,2,…,13)是一簇独立的标准Gauss白噪声.另外,函数bk是反应Rk的倾向函数(propensity function),被定义为反应Rk的反应物浓度与相应反应速率常数的乘积[19],如b1(Yi(t),Vi(t))=3αm(Vi(t))Y1i(t).函数σk满足
网络中的各神经元通过化学突触相互连接,其树突上接收递质的突触通道是配体门控的,可用α电导模型来刻画;释放递质的轴突末端具有短时程突触可塑性.因此,由α电导的微分方程[20]和短时程突触可塑性模型[3]得到如下突触动力学模型:
(5)
式(5)中:si(t),hi(t)和τs分别为α电导的突触通道开放比例,递质水平变化率和衰退时间常数;ri(t)和ui(t)分别为t时刻神经元轴突末端贮藏的囊泡比例和囊泡释放概率,满足0≤ri(t)≤1,u0≤ui(t)≤1,其中ui(t)由突触末端的残余钙离子水平所决定,残留钙离子水平越高,ui(t)就越大[3];τD,τF分别为ri(t),ui(t)的衰退时间常数,u0是ui(t)的基准值.另外,δ为Dirac-delta函数;τd为神经递质传输延迟,服从1~5 ms上的均匀分布[3];tki为第i个神经元第k次发放时刻.这样,ri(t)和ui(t)在tki时刻上的跳跃反映了该神经元递质的量子式释放和钙离子的流入;而hi(t)在tkj+τd时刻上的跳跃代表接收第j个神经元释放的递质.
联立式(1)、式(4)和式(5),构成一个耦合的跳扩散随机微分方程组,其初值条件如下:对i=1,2,…,N,Vi(0)为-80~-65 mV的均匀分布随机数;Yi(0)由
且Yi(0)∈SY确定;ri(0)=1;ui(0)=u0.本文采用步长为0.01 ms的前向Euler-Maruyama算法进行数值模拟,进而得到一个离散时刻在0~3 000 ms的近似样本序列.在模拟中,对Yi的模拟值未落在SY的情形采用文献[14]给出的方法进行截断处理,tki是通过设置10 mV的阈值和2 ms的绝对不应期来确定的一个离散时刻.数值模拟的其他参数见表1.

表1 随机HH神经元网络参数
2 结 果

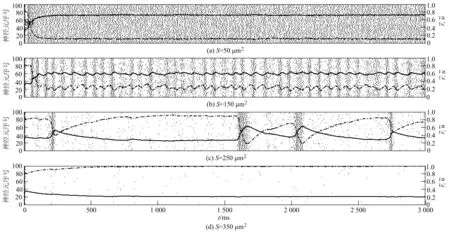
黑点表示神经元的一次发放,虚线和实线分别表示(t)和(t)随t的变化情况图1 当τD=200 ms,τF=1 500 ms时不同膜面积下神经网络的发放情况
黑点表示神经元的一次发放,虚线和实线分别表示(t)和(t)随t的变化情况图2 当τD=200 ms,τF=200 ms时不同膜面积下神经网络的发放情况
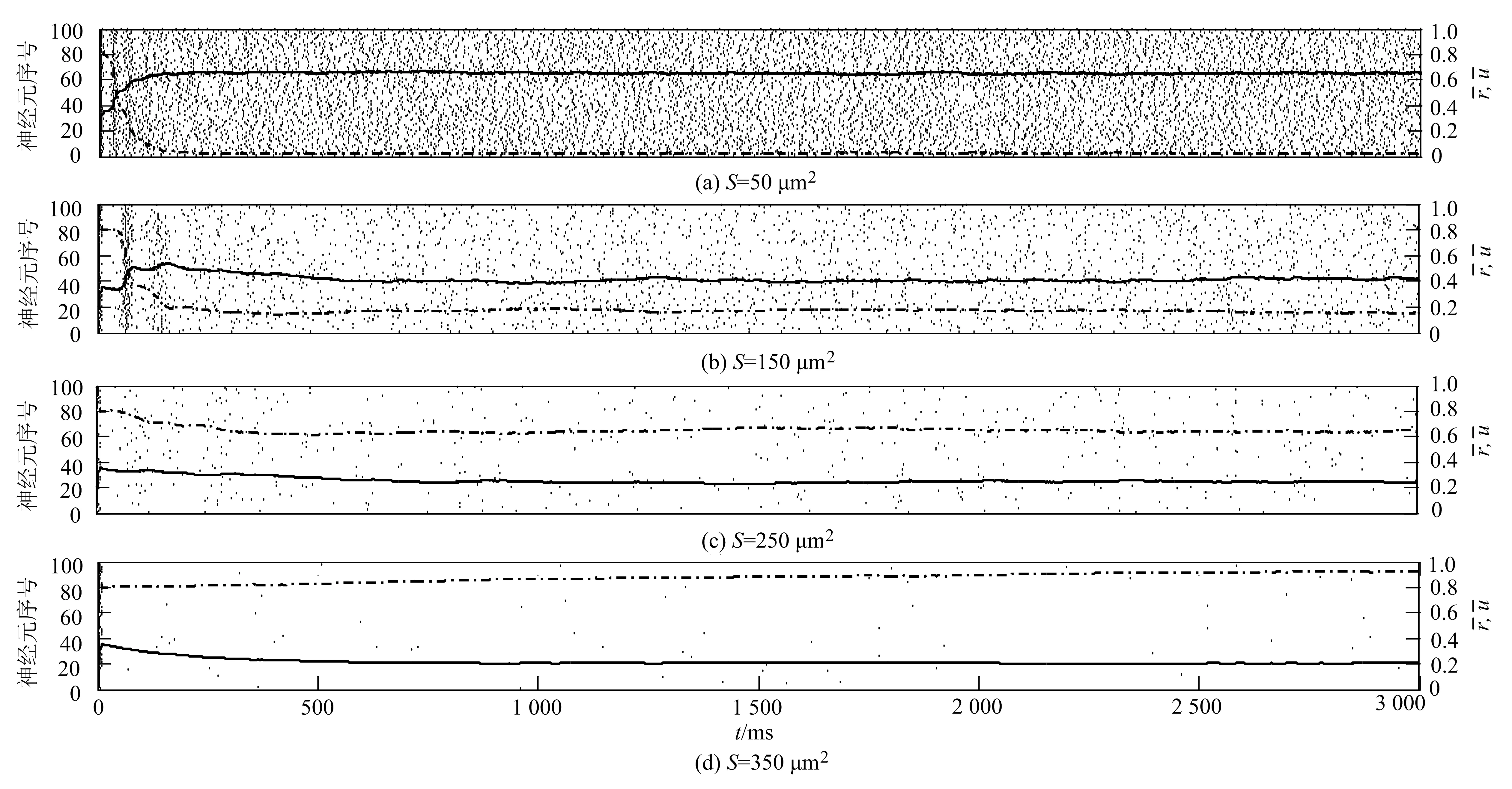
黑点表示神经元的一次发放,虚线和实线分别表示(t)和(t)随t的变化情况图3 当τD=1 500 ms,τF=200 ms时不同膜面积下神经网络的发放情况
为进一步系统地研究该网络的性质,需引入发放率R、变异系数CV、同步指数C及突触效能(synaptic efficacy)E,分别刻画该网络的活跃性、时间相干性、空间相干性及突触可塑性.R,CV和C的定义在文献[17]中给出,并且R和C越大分别代表网络的活跃性、空间相干性越好,而更小的CV对应更好的时间相干性.定义E为
其中:T为观察时间(取为3 000 ms);r(tki)u(tki)是第i个神经元在第k次发放释放的囊泡比例.易见,E越大,网络的突触传递效率越高.上述4个指标通过20次独立重复的数值模拟的平均结果来估计.下面分别取S值为50,150,250,350 μm2,研究不同的τD和τF值对R,CV,C和E的影响 (见图4).
首先比较图4(a).R在区域中的最大值随着S值的增大而减小.当S=50 μm2时,R随τF变化基本保持不变,但可以随着τD增大而增大,并趋于某个水平;当S=150,250 μm2时,τF越大,R越大,τD越大,R越小;当S=350 μm2时,R在大部分区域小于1 Hz,仅在τF≫τD的部分超过1 Hz.
接着比较图4(b).E在区域中的最大值在S值较大时处于较高水平.当S=50,150 μm2时,若τD小于某个水平,则τF的增大会引起E增大,τD的增大会导致E减小;当S=250,350 μm2时,这种现象几乎在整个区域都会出现.值得注意的是,当S=350 μm2时,E与τD,τF接近线性关系.
最后,分别比较图4(c)和图4(d).当S=50 μm2时,τF不会影响CV和C,但τD的增大却可以导致CV的增大和C的减小;当S=150,250 μm2时,需要在τD较小时,随着τF变化CV和C才会出现明显变化;当S=350 μm2时,CV和C在R较小的区域内数值较低,在R较大的区域内数值较高.

(a),(b),(c)和(d)4列分别对应网络的R,E,CV和C在不同S下关于τD,τF的变化;第1~4行分别表示面积S=50,150,250,350 μm2图4 不同S,τD,τF对网络性质的影响
3 讨 论
本文构造了一个由通道噪声和突触噪声驱动的全连接的HH兴奋型神经网络,并且在突触噪声中通过引入突触变量ri(t),ui(t)来反映短时程突触可塑性.进一步分析模拟结果并结合生物意义可知,τD和τF分别反映了突触可塑性的抑制和易化强度.在网络中,τD值越大,对神经元发放的抑制效果越强,而τF值增加可以促进神经元发放.具体见图1和图3.
上述结论也可从式(5)得出:若τD值越大,则ri(t)上升趋于1的速率越慢,其中ri(t)表示突触末端贮藏的囊泡比例.贮藏的囊泡个数始终保持较低的值,导致每次释放的囊泡数量较少,从而抑制神经元发放.类似地,若τF值越大,则ui(t)下降趋于u0的速率越慢,其中ui(t)表示突触末端囊泡释放的概率.囊泡始终保持较高的释放概率,致使突触后神经元接收的神经递质的量增加,从而促进神经元的发放.
图4的结果说明:当τD足够小且S值适中时,τD和τF的增加可以分别降低和增加网络的活跃性、时空相干性和突触传递效率.由式(4)可知,S值与通道噪声的强度成反比.在S=50 μm2的情形中,通道噪声过强,使得τF对4个指标的影响不明显,但τD仍能影响这4个指标;在S=350 μm2的情形中,通道噪声较弱,网络发放不频繁,使得τD和τF的变化均不能显著影响网络的活跃性和时空相干性,但是E仍可受到τD和τF影响,并且由于过少的发放使得ri(t)和ui(t)维持在一个平稳水平,从而使这种影响呈现一种接近线性的关系.同时发现,即使通道噪声减弱,短时程抑制的存在仍可以破坏网络的发放同步性.因此,突触可塑性使得突触噪声与通道噪声协同作用更为复杂多样.
[1]Zucker R S,Regehr W G.Short-term synaptic plasticity[J].Annu Rev Physiol,2002,64(1):355-405.
[2]Tsodyks M,Pawelzik K,Markram H.Neural networks with dynamic synapses[J].Neural Comput,1998,10(4):821-835.
[3]Mongillo G,Barak O,Tsodyks M.Synaptic theory of working memory[J].Science,2008,319(5869):1543-1546.
[4]Romani S,Tsodyks M.Short-term plasticity based network model of place cells dynamics[J].Hippocampus,2015,25(1):94-105.
[5]Hamill O P,Marty A,Neher E,et al.Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches[J].Pflügers Archiv,1981,391(2):85-100.
[6]Hille B.Ion channels of excitable membranes[M].Sunderland:Sinauer Associates,2001.
[7]Hodgkin A L,Huxley A F.A quantitative description of membrane current and its application to conduction and excitation in nerve[J].J Physiol,1952,117(4):500-544.
[8]Skaugen E,WallØe L.Firing behaviour in a stochastic nerve membrane model based upon the Hodgkin-Huxley equations[J].Acta Psychiatr Scand,1979,107(4):343-363.
[9]Clay J R,Defelice L J.Relationship between membrane excitability and single channel open-close kinetics[J].Biophys J,1983,42(2):151-157.
[10]Chow C C,White J A.Spontaneous action potentials due to channel fluctuations[J].Biophys J,1996,71(6):3013-3033.
[11]Fox R F,Lu Y.Emergent collective behavior in large numbers of globally coupled independently stochastic ion channels[J].Bakhtiniana Revista De Estudos Do Discurso,2012,7(49):3421-3431.
[12]Pakdamank K,Thieullen M,Wainrib G,et al.Fluid limit theorems for stochastic hybrid systems with application to neuron models[J].Adv App Probab,2010,42(3):761-794.
[13]Goldwyn J H,Imennov N S,Famulare M,et al.Stochastic differential equation models for ion channel noise in Hodgkin-Huxley neurons[J].Phys Rev E,2011,83(4):041908.
[14]Huang Yandong,Rüdiger S,Shuai Jianwei.Langevin approach for stochastic Hodgkin-Huxley dynamics with discretization of channel open fraction[J].Phys Lett A,2013,377(44):3223-3227.
[15]Yu Y,Liu F,Wang W.Synchronized rhythmic oscillation in a noisy neural network[J].J Phys Soc Japan,2003,72(12):3291-3296.
[16]Chen J,Ding S,Li H,et al.Synchronization and array-enhanced resonances in delayed coupled neuronal network with channel noise[J].Chaos:An Interdisciplinary Journal of Nonlinear Science,2014,24(3):033131.
[17]Zhang X,Li H,Chen J,et al.The impact of channel and external synaptic noises on spatial and temporal coherence in neuronal networks[J].Neurocomputing,2015,164:230-239.
[18]Sakumura Y,Ishii S.Stochastic resonance with differential code in feedforward network with intra-layer random connections[J].Neural Networks,2006,19(4):469-476.
[19]Gillespied D T.A rigorous derivation of the chemical master equation[J].Physica A:Statistical Mechanics and its Applications,1992,188(1):404-425.
[20]Destexhe A,Mainen Z F,Sejnowski T J.Synthesis of models for excitable membranes, synaptic transmission and neuromodulation using a common kinetic formalism[J].J Comput Neurosci,1994,1(3):195-230.
(责任编辑陶立方)
Short-term synaptic plasticity based network of stochastic Hodgkin-Huxely neurons
ZHOU Jiacheng,DING Shaojie,ZHANG Xuejuan
(CollegeofMathematics,PhysicsandInformationEngineering,ZhejiangNormalUniversity,Jinhua321004,China)
According to the physiological mechanism of the vesicle release and random switching of ion channels, a short-term synaptic plasticity based network model of stochastic Hodgkin-Huxely neurons was proposed. The effect of the channel noise and the synaptic noise on the firing behavior of the network and the influence of facilitation and depression to synaptic efficacy were discussed.
short-term synaptic plasticity; facilitation; depression; network model; Hodgkin-Huxley neuron
10.16218/j.issn.1001-5051.2016.03.006
收文日期:2015-09-06;2015-11-26
周家程(1992-),男,浙江台州人,硕士研究生.研究方向:计算神经科学.
张雪娟.E-mail: xuejuanzhang@gmail.com
O211.63;Q332
A
1001-5051(2016)03-0268-08
