乌兰布和沙漠霸王与白刺秋季光合日变化特征比较
2016-09-26黄雅茹刘芳马迎宾葛根巴图余新春吕永军董礼隆
黄雅茹,刘芳,马迎宾,葛根巴图,余新春,吕永军,董礼隆
(中国林业科学研究院沙漠林业实验中心,内蒙古 磴口 015200)
乌兰布和沙漠霸王与白刺秋季光合日变化特征比较
黄雅茹,刘芳,马迎宾,葛根巴图,余新春,吕永军,董礼隆
(中国林业科学研究院沙漠林业实验中心,内蒙古 磴口015200)
【目的】 在自然状态下研究乌兰布和沙漠东北部的霸王与白刺的光合日变化特征.【方法】 采用Li-6400xt便携式光合测定系统分析仪测定光合日动态.【结果】 霸王与白刺的净光合速率日变化呈双峰型,峰值均出现在10∶00与16∶00,有明显的光合“午休”现象,且主要是由气孔因素引起的.白刺净光合速率日均值(4.91±0.74)μmol/(m2·s)大于霸王(3.24±0.76)μmol/(m2·s);2种植物蒸腾速率日变化呈双峰型,峰值出现在10∶00与16∶00,白刺蒸腾速率日均值(4.15±0.31)mmol/(m2·s)大于霸王(2.44±0.33)mmol/(m2·s);霸王与白刺水分利用效率日变化曲线呈双峰型.【结论】 相同环境条件下霸王的水分利用效率高于白刺.
光合特性;日变化;霸王;白刺
白刺(NitrariatangutorumBobr)是蒺藜科白刺属植物,匍匐性小灌木,俗称地枣、地椹子、沙樱桃等。白刺分布于陕西、内蒙古、宁夏、甘肃等地。白刺为旱生型阳性植物,耐盐性能极强,多生长在干燥、多风、盐碱重、土壤贫瘠、植物稀疏的严酷环境中,往往自成群落,伴生植物较少.霸王(SarcozygiumxanthoxylonBunge)是蒺藜科霸王属的荒漠植物,分布于内蒙古西部,适应性强,对气候土壤要求不高,耐干旱、耐盐碱,在沙漠地区根系发达,萌生力强,耐风蚀、耐贫瘠.白刺与霸王是我国西北部干旱荒漠区主要的优势种和建群种之一,对于防风固沙有重要作用.
在干旱半干旱区,由于降水稀少,蒸发强烈,沙区植被常常会面临不同程度的土壤干旱环境.水分是影响植物生长和水分利用效率的关键因子[3],并与温度、光照、CO2浓度等环境因子共同决定了植物对环境的响应与适应[4].不同植被适应干旱的方式不同,目前大多学者通过测定植物的光合速率,蒸腾速率等相关生理指标,不同程度评估植物的抗旱性,进一步评价其适应性.吉小敏[5]对不同水分条件下大田栽植的梭梭和多花柽柳苗期的净光合速率、蒸腾速率、水分利用效率、气孔导度和胞间CO2浓度等光合生理指标进行了测定,表明梭梭的抗旱性强于多花柽柳.何炎红[6]对沙冬青等3种沙生植物的光合生理进行了研究,表明较低的水分利用效率是沙生植物长期适应干旱高温气候环境的结果.张利刚[7]对绿洲-荒漠过渡带多枝柽柳、梭梭、头状沙拐枣和沙枣的光合特性、水势以及生理指标进行了研究,运用平均隶属函数法对这4种植物进行抗旱性排序依次是梭梭>多枝柽柳>沙枣>头状沙拐枣.尤扬[8]以多年生大叶黄杨为研究对象,采用Li-6400便携式光合测定仪测定其光合生理指标的日变化和光合指标响应,结果表明,净光合速率呈双峰型,存在光合“午休”现象.李清河[9-10]对乌兰布和沙漠东北部的甘蒙柽柳、沙木蓼、霸王、沙冬青、花棒、白刺、梭梭和柠条8种沙生灌木生长季末期的光合生理特性进行了研究,还对不同种源白刺幼苗生理生长对水分的响应进行了研究.由于白刺与霸王是乌兰布和沙漠东北部主要的建群种和优势种之一,且在乌兰布和沙漠分布广泛,尤其在乌兰布和沙漠东北部分布较集中,为主要的固沙灌木植被群落,而目前学者们对乌兰布和沙漠主要灌木的光合日动态特性研究较少,因此,本试验主要针对内蒙古乌兰布和沙漠2种优势植物白刺与霸王光合日变化特征进行研究,为进一步评价其抗旱性、适应性提供基础,为乌兰布和沙漠植物筛选提供参考.
1 材料与方法
1.1试验地概况
研究区位于内蒙古自治区磴口县中国林业科学研究院沙漠林业实验中心第三实验场附近.该区是河套平原与乌兰布和沙漠的结合部,属于温带大陆性干旱气候,年平均气温7.8 ℃,多年平均降水量约145 mm,降雨集中在6~9月份,年蒸发量约2 327 mm,年日照时数约3 210 h,占全年可照时数的72%,无霜期136 d,土壤类型为风沙土,主要植被为沙地灌木植被.
1.2试验方法
以优势植物霸王与白刺为研究对象,选择全天晴朗无云的天气,选择完整、生长健康的植株,采用Li-6400xt便携式光合测定系统分析仪(USA,LICOR)对植被进行叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)等的测定,同时采用美国生产的HOBO小型自动气象站对地面2m以内主要气象因子光合有效辐射(PAR)、气温(Ta)、相对湿度(RH)、大气CO2浓度(Ca)进行测定,测定时间与光合测定时间同步进行,仪器布设在样地内,重复三次.查阅相关文献,植物叶片水分利用效率(WUE)=Pn/Tr[11],气孔限制值的计算参照Berry等的方法,Ls=1-Ci/Ca[12].各材料随机选择6片叶片进行测定,测定时间为2013年8月23日到25日,连续测定3 d,数据重复次数为18次,每天测定时间为8∶00~20∶00,每2 h观测1次,并取其平均值为该植物的日均值.测定完成后,将叶片做好标记,装入自封袋,带回实验室内,用扫描仪扫描后存入电脑,用Ipp 6.0图像分析软件计算叶片面积.
数据处理使用Excel 2003,误差值采用函数STDEVP计算,气象因子误差值的计算是采用3次数据的重复,叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)等的计算以及误差值计算是采用18次数据的重复.
2 结果与分析
2.1主要气象因子日变化特征
由图1可知,光合有效辐射(PAR)在14∶00达到最大值,为796.36 μmol/m2,之后逐渐下降,日变化呈典型的钟罩型.在强烈的光照下,空气温度也随之变化,最高气温出现在14∶00,为34.09 ℃.在光照与温度的变化下,相对湿度(RH)和大气CO2浓度(Ca)呈现出相反的日变化特征,早晨和傍晚较高,有利于光合作用,而中午较低,不利于光合作用.RH最低值出现在16∶00,为16.36%,Ca最低值出现在14∶00,为380.59 μmol/mol.

图1 主要气象因子日变化特征Fig.1 Diurnal variations of main meteorological factors
2.2净光合速率(Pn)日变化
霸王与白刺净光合速率(Pn)日变化过程均表现为双峰型(图2),且最大值均出现在上午10∶00,分别为(7.14±0.85)μmol/(m2·s)和(11.10±0.65)μmol/(m2·s),白刺大于霸王;第2峰值出现在午后16∶00,峰值相对较小,分别为(4.97±0.95)μmol/(m2·s)和(7.94±0.56)μmol/(m2·s),白刺的净光合速率也大于霸王;全天日平均净光合速率为白刺大于霸王,其值分别为(4.91±0.74)μmol/(m2·s)和(3.24±0.76)μmol/(m2·s).相同环境条件下白刺与霸王植物叶片光合日变化曲线表现出一定的相似性.

图2 净光合速率日动态Fig.2 Diurnal variations of net photosynthesis rate
2.3蒸腾速率(Tr)日动态
霸王与白刺蒸腾速率日变化特征与净光合速率变化特征相似(图3).2种植物蒸腾速率峰值出现在10∶00与16∶00,霸王与白刺的最大蒸腾速率分别为(3.50±0.36)mmol/(m2·s)与(6.33±0.15)mmol/(m2·s),日平均蒸腾速率分别为(2.44±0.33)mmol/(m2·s)与(4.15±0.31)mmol/(m2·s),蒸腾速率无论是最大值还是全天日平均值白刺均大于霸王.

图3 蒸腾速率(Tr)日动态Fig.3 Diurnal variations of transpiration rate
2.4水分利用效率(WUE)日动态
霸王与白刺水分利用效率日变化曲线呈双峰型(图4).相同环境条件下霸王的水分利用效率高于白刺,日平均水分利用效率是白刺的1.18倍;2种植物的水分利用效率最大值出现在上午,这与一般植物的特征相同,且霸王的最大水分利用效率大于白刺.
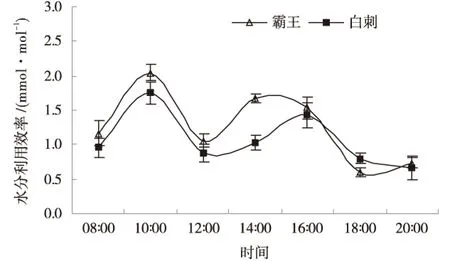
图4 水分利用效率日动态Fig.4 Diurnal variations of water use efficiency
2.5气孔导度(Gs)、气孔限制值(Ls)及胞间CO2浓度(Ci)日动态
霸王与白刺气孔导度的日变化曲线如图5所示,2种植物气孔导度日变化均是早晨较高,之后随着温度升高和空气相对湿度降低,气孔逐渐关闭,到中午达到最低.2种植物无论是最低值还是日均值都是白刺高于霸王,其中白刺日均值为(0.12±0.01)mol/(m2·s),霸王日均值为(0.10±0.02)mol/(m2·s).霸王与白刺的气孔限制值日变化趋势与气孔导度的日变化相反(图6),早晨最低,随着温度升高与空气相对湿度降低,气孔限制值增大.霸王的气孔限制值日均值大于白刺.霸王与白刺的胞间CO2浓度日变化曲线是单峰型(图7),早晨与晚上高,中间最低.胞间CO2浓度日均值分别为(282.77±15.88)μmol/mol与(319.04±15.67)μmol/mol,白刺大于霸王.

图5 气孔导度日动态Fig.5 Diurnal variations of stomatal conductance

图6 气孔限制值日动态Fig.6 Diurnal variations of stamatal limitation

图7 胞间CO2浓度日动态Fig.7 Diurnal variations of intercellular CO2 concentration
3 讨论
光合作用是植物生长与发育的基础,同时也是植物生产力构成的最主要影响因素.气孔是植物与外界环境进行气体和水分交换的门户和通道,对水分变化十分敏感,几乎所有的中生和旱生植物都可以通过关闭气孔来适应无间叶片过度蒸腾或低水势的土壤环境[13].从植物本身来说,光合作用的限制因素大致可分为气孔因素和非气孔因素.Farquhar等的研究表明,当净光合速率和胞间CO2浓度的变化方向相同(同时减小)且气孔限制值增大时,净光合速率减少的主要原因是由气孔因素引起的;否则净光合速率的下降的原因是由于非气孔因素引起[14-15].本研究中,霸王与白刺净光合速率日变化曲线均存在不同程度的光合“午休”现象.霸王与白刺在10∶00~12∶00时间段内净光合速率下降,胞间CO2浓度下降,且气孔限制值增大,可以推断光合“午休”现象是由气孔因素引起的.
植物蒸腾是植物生理特性之一,植物蒸腾速率(Tr)不仅受气候和土壤水分的影响,而且还受植物种类等多种因素的制约,因此导致了不同的植物表现出不同的蒸腾速率(Tr)变化.植物的气孔多分布于叶片表面,既能保证植物与外界环境进行气体交换,又能适当抑制蒸腾作用[16].本研究中霸王与白刺在8∶00~10∶00间,蒸腾速率呈上升趋势,到10∶00蒸腾速率达到一天中的峰值,此后随温度的升高而下降,到16∶00达到第二个峰值,但明显低于第1个峰值,蒸腾速率呈双峰型.由于中午光照强度最大,气温最高,蒸腾速率逐渐增大,然而水分供应不足,为了保护叶片不被灼伤,减少水分的损耗,气孔会自动调节,开度明显减小或部分关闭一段时间,Tr大幅降低.当光强和气温在午后逐渐减弱时,蒸腾作用又逐渐增强,Tr值有所回升,最后随着PAR,Ta等气象因素的下降以及气孔的逐渐关闭,Tr又趋于减弱[17].蒸腾作用午间降低是中生和旱生植物通过关闭气孔来适应午间高温或防止叶片过度蒸腾失水的一种生态适应对策[18].
水分利用效率(WUE)是指植物消耗单位重量的水所固定二氧化碳的量,是评价植物耗水量多少或适应干旱胁迫能力的一个重要的生理生态指标,其值越大,说明植物对水分的利用效率越高[19],植物对环境的适应性就可能发生改变[20].植物水分利用效率受净光合速率与蒸腾速率的共同影响.本试验中,霸王与白刺水分利用效率均呈双峰型,在上午时段的水分利用效率高于下午,这与丁晓纲[21]的研究结果一致,最高值出现在10∶00左右,此时光合有效辐射较强,气孔开度较大,空气相对湿度较大,净光合速率增加较快,而蒸腾速率相对较低,水分利用效率较高,10∶00以后由于光照增强、气温上升、空气相对湿度明显下降,部分气孔关闭,光合速率降低的幅度小,蒸腾速率降低的幅度大,水分利用效率减小.Ci/Ca(胞间CO2浓度/大气CO2浓度)是一个抗旱性指标,表征植物对水分变化的响应,Ci/Ca越小,抗旱性越强[22],Ls=1-Ci/Ca,即气孔限制值越大,一定程度上说明抗旱性越强,在相同的环境条件下,霸王的Ls最大,水分利用效率日均值最高,有研究表明,水分利用效率在一定程度上可以作为评价植物抗旱性的强弱指标之一,本文仅研究了霸王与白刺的秋季光合日变化特征,而对于其它时间段的光合特征以及抗旱性、适应性如何,还需要进一步观测与研究.
4 结论
1)霸王与白刺的净光合速率日变化呈双峰型,峰值均出现在10∶00与16∶00,有明显的光合“午休”现象,且主要是由气孔因素引起的.
2)白刺净光合速率日均值为大于霸王,其值分别为(4.91±0.74)μmol/(m2·s)和(3.24±0.76)μmol/(m2·s);两种植物蒸腾速率日变化呈双峰型,峰值出现在10∶00与16∶00,白刺蒸腾速率日均值(4.15±0.31)mmol/(m2·s)大于霸王(2.44±0.33)mmol/(m2·s).
3)霸王与白刺水分利用效率日变化曲线呈双峰型,相同环境条件下霸王的水分利用效率高于白刺.
[1]吴鸿宾.内蒙古主要气象灾害分析[M].北京:气象出版社,1990
[2]陈炳浩,郝玉光.乌兰布和沙区区域性防护林体系气候生态效益评价的研究[J].林业科学研究,2003,16(1):63-68
[3]O'Connor T G,Haines L M,Snyman H A.Influence of precipitation and species composition on phytomass of a semi-arid African grassland[J].Journal of Ecology,2001,89:850-860
[4]Villalobos A E,Pel ez D V.Influence of temperature and water stress on germination and establishment ofProsopiscaldeniaDurk[J].Journal of Arid Environments,2001,49(2):321-328
[5]吉小敏,宁虎森,梁继业.不同水分条件下梭梭和多花柽柳苗期光合特性及抗旱性比较[J].中国沙漠,2012,32(2):399-406
[6]何炎红,田有亮,李建,等.沙冬青等3种沙生植物气体交换特征[J].干旱区资源与环境,2014,26(7):144-149
[7]张利刚,曾凡江,刘波.绿洲-荒漠过渡带四种植物光合及生理特征的研究[J].草业学报,2012,21(1):103-111
[8]尤扬,周秀梅,李保印,等.大叶黄杨秋季光合特性研究[J].甘肃农业大学学报,2014,49(2):120-124
[9]李清河,刘建锋,张景波,等.乌兰布和沙漠东北部8种沙生灌木生长季末期的光合生理特性[J].西北植物学报,2006,26(11):2318-2323
[10]李清河,张景波,李慧卿,等.不同种源白刺幼苗生理生长对水分梯度的响应差异[J].林业科学,2008,4(1):52-56
[11]Fischer R A,Turner N C.Plant productivity in the arid and semiarid zones[J].Annual Review of Plant Physiology,1978,29:227-317
[12]Berry J A,Downton W J S.Environmental regulation of photosynthesis[M]∥Photosynthesis:Development,carbon metabolism and plant productivity,Vol.Ⅱ.New York:Academic Press,1982:263-343
[13]温达志,周国逸,张德强,等.四种禾本科牧草植物蒸腾速率与水分利用效率的比较[J].热带亚热带植物学报,2000(增刊Ⅲ):67-76
[14]Sharkey T D.Estimation the rate of photorespiration in leaves[J].Physiol Plant,1988,73:147 -152
[15]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Rev Plant Physiol,1982,33:31-34
[16]Sam O,Jerdz E,Dell-Amico J,et al.Water stress induced changes in anatomy of tomato leaf epidermis[J].Biologia Plantarum,2000,43:275-277
[17]张淑勇,周泽福,张光灿,等.半干旱黄土丘陵区天然次生灌木山桃与山杏叶片气体交换参数日动态差异[J].生态学报,2009,29(1):499-507
[18]廖行,王百田,武晶.不同水分条件下核桃蒸腾速率与光合速率的研究[J].水土保持研究,2007,14(4):30-34
[19]樊卫国,李迎春.梨属4个重要种的光合特性及水分利用效率[J].西南农业学报,2006,19(6):1144-1146
[20]孙惠玲,马剑英,王绍明,等.准格尔盆地荒漠植物碳同位素组成研究[J].中国沙漠,2007,27(6):972-976
[21]丁晓纲,何茜,李吉跃,等.毛乌素沙地樟子松和油松人工林光合生理特性[J].水土保持研究,2011,18(1):215-219
[22]Terashima I,Hikosaka K.Comparative ecophysiology of leaf and canopy photosynthesis[J].Plant,Cell & Environment,1995,18(10):1111-1128
(责任编辑李辛)
A comparison of diurnal variations of photosynthetic charac-teristics ofNitrariatangutorumandSarcozygiumxanthoxylonin Autumn in Ulanbuh Desert
HUANG Ya-ru,LIU Fang,MA Ying-bin,GE GEN Ba-tu,YU Xin-chun,LYU Yong-jun,DONG Li-long
(Experimental Center of Desert Forestry,Chinese Academy of Forestry,Dengkou 015200,China)
【Objective】 The diurnal variations of photosynthetic characteristics ofS.xanthoxylonandN.tangutorumwere studied under natural environment.【Method】 Determination of Li-6400 xt portable photosynthesis system analyzer was adopted to measure photosynthesis dynamic.【Result】 Diurnal variations of net photosynthesis rate ofS.xanthoxylonandN.tangutorumwere double-peak curves,the peaks were 10∶00 and 16∶00,had obvious siesta phenomena of photosynthesis,and the affecting factors were mainly stomata ones.Average net photosynthesis rate ofN.tangutorumwas greater thanS.xanthoxylon,the value respectively was (4.91±0.74)μmol/(m2·s) and (3.24±0.76)μmol/(m2·s).Diurnal variations of transpiration rate ofS.xanthoxylonandN.tangutorumwere double-peak curves,the peaks were 10∶00 and 16∶00,average transpiration rate ofN.tangutorumwas greater thanS.xanthoxylon,the value respectively was (4.91±0.74)μmol/(m2·s) and (3.24±0.76)μmol/(m2·s).Diurnal variations of water use efficiency ofS.xanthoxylonandN.tangutorumwere double-peak curves.【Conclusion】 Water use efficiency ofS.xanthoxylonwas greater thanN.tangutorumunder the same conditions.
photosynthetic characteristics;diurnal variations;SarcozygiumxanthoxylonBunge;NitrariatangutorumBobr
黄雅茹(1987-),女,硕士,工程师,主要从事荒漠化防治研究.E-mail:hu_angyaru@126.com
刘芳,女,硕士,高级工程师,主要从事荒漠化防治技术与干旱区抗逆植物资源筛选研究.E-mail:liuf98@126.com
中央级院所基本科研业务专项(CAFYBB2012003).
2015-01-15;
2016-04-26
Q 945.79
A
1003-4315(2016)04-0078-06
