甜菜不育系育性核基因差异对应的2DE蛋白图谱差异分析
2016-04-23贾敢敢张惠忠李晓东付增娟
贾敢敢,白 晨,张惠忠,李晓东,张 辉,付增娟,王 良
(内蒙古农牧业科学院,内蒙古呼和浩特 010031)
甜菜是中国北方的重要的糖料作物,随着杂种优势理论提出及其实践效应证明,杂交育种在各类农作物育种过程中已屡见不鲜。因甜菜雌雄同体、花器较小等一系列原因造成的杂交育种过程中的困难就需要不育系来解决,普遍应用甜菜CMS(Cytoplasmic Male Sterility)接受其对应的保持系花粉进行繁殖[1]。
在所发现的甜菜不育系类型中分为核不育、质不育、质核互作不育3种类型,其中质核互作造成的不育植株在甜菜杂交育种中广泛应用。近年来对甜菜质核互作不育植株(CMS)的研究成果也越来越多,结合前人对甜菜线粒体基因的特异序列研究及引物开发的成果,可以初步辨别不育系选育过程中出现的诸多类型。目前,对甜菜CMS植株的研究应用集中表现在核质基因特异序列的分子鉴定技术的广泛应用以及甜菜蛋白质组学的差异蛋白分析。2009年黑龙江大学曹洪祥等[2]对甜菜M14品系的花期花器蛋白的双向电泳差异分析检测到1 000+的蛋白质点,牛佳[3]等2013年也得到了高丰度的蛋白质点图谱,得到400+的蛋白质点并对甜菜保持系及对应不育系的差异蛋白进行了初步分析,提出甜菜育性相关基因的开始表达时期和花期同育性相关的几种蛋白。同时王有昭在2009年对国内的55个甜菜品种的胞质基因进行VNTR序列电泳检测并得到了准确的多类型不育系甜菜植株的检测结果及特异重复序列的测定[4]。且课题组在不久前对内蒙古农牧业科学院特色作物所的现有不育系种质资源进行了不育植株类型的检测[5]。
在很多物种中都已经发现了决定CMS的线粒体基因,例如,在日本野生萝卜中发现的小仓型(Ogura)细胞质雄性不育[6]是由雄性不育诱导基因(orf138)决定[7-9],且推测该基因是基因内重组的结果,并且这个位点的特殊结构决定了orf138基因的稳定性。同时绝大多数的核恢复基因会影响CMS基因,导致CMS相关的双顺反mRNAs突变,进而导致其伴侣基因表达的单顺反mRNAs的增加。鉴于甜菜育种的两系杂交特殊性在研究方面侧重于不育系和保持系的差异蛋白分离、鉴定及各差异蛋白的相互关系。
本试验将田间调查和分子标记辅助育种技术相结合,对选育过程中多种类型不育系进行双向蛋白电泳差异比对,以1945年美国科学家F.Owen报道提出的雄性不育机制猜想为理论基础,进一步探究其在分子标记技术上的差异是否存在对应的蛋白表达差异,并寻求育性相关核基因的表达规律。
1 材料和方法
1.1 材料
1.1.1 植物材料 以内蒙古农牧业科学院特色作物所拟选育的甜菜CMS植株为材料,共取91株。
1.1.2 引物 甜菜细胞质育性相关的小卫星分子标记引物TR1、TR3,引物序列由国家甜菜产业技术体系分子育种岗位提供,引物由南京金斯瑞生物公司完成。其引物序列见表1。
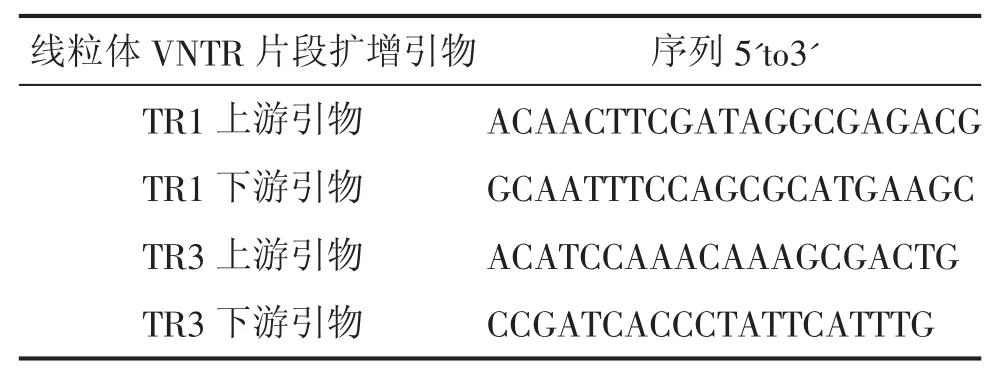
表1 对线粒体VNTR片段的扩增引物序列
1.1.3 试剂 植物DNA提取试剂盒、丙酮、TCA、DTT、碘乙酰胺、SDS、尿素、甘氨酸等。
1.2 方法
1.2.1 CMS型选育过程中的单株出粉调查 从甜菜CMS植株的发现,人们一直对甜菜质核互作的核基因调控情况存在疑惑。一种认为其为两对显性基因X、Z控制,另一种则认为其是由一组数量基因来控制的。本试验对选育过程中的P2代CMS系列进行田间花粉调查并记录。
1.2.2 PCR扩增 利用TR1、TR3引物,对所调查的CMS系列植株的全基因组进行扩增。25 μL体系含12.5 μL 2×Tap Master Mix,1 μL 上游引物,1 μL 下游引物,2 μL 基因组 DNA,8.5 μL ddH2O。PCR 程序为94℃预变性5 min;94℃变性30 s,55℃退火30 s,72℃延伸1 min,循环35;72℃延伸10 min,4℃保存。待PCR结束后将其扩增产物于1.2%的琼脂糖凝胶上电泳,凝胶成像仪上照相保存。
1.2.3 蛋白提取 采用丙酮(TCA)沉淀法分别对CMS系列植株进行苗期叶片、花期花序的蛋白提取。
公司立足广东农垦,面向“三农”,提供具有农业特色的小微金融服务,构建良好的农业产融生态,核心业务板块包括产业集团供应链金融业务、垦区金融业务、农批交易市场金融业务等。
等电聚焦:采用伯乐IEF等电聚焦仪进行第一项水化蛋白等电聚焦,聚焦方案参考蛋白双向电泳培训手册并进行优化,已得到高通量蛋白图谱。
SDS-PAGE凝胶电泳:将第一项等电聚焦完毕的IPG预制胶条转入垂直板12%聚丙烯酰胺凝胶中电泳。电泳完毕后采用经典考染法进行蛋白凝胶染色,于凝胶成像仪中照相保存。
2 结果与分析
2.1 对来自同一系列的P2 CMS单株进行花期花粉调查分类
受试植株于2015年4月1日播种窖藏母根,同年6月1日对抽薹开花植株进行花粉出粉情况调查,见表2。
对内蒙古农牧业科学院特色作物所选育中的不育系进行二年生阶段的田间植株性状调查,种植母根由上年同系单株对应编号分株收获、窖存,次年栽植田间。由表2可知,选育过程中的不育系系列植株在花粉出粉情况方面存在一定的多样性,能够从中筛选出供后续试验选用的目的材料。
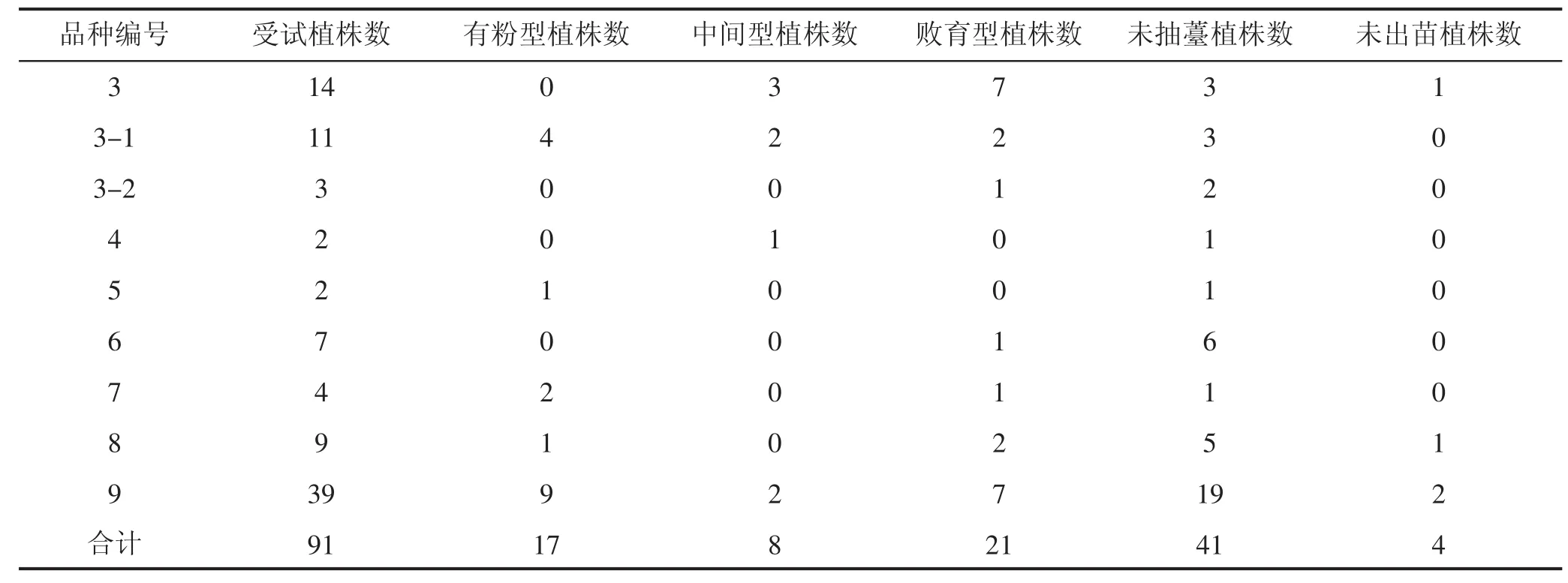
表2 甜菜不育系田间植株出苗、抽薹、花粉情况
2.2 对调查中不同类型的CMS单株进行基因组DNA提取并对其进行VNTR微卫星分子标记鉴定
从所选植株中选取具有代表性的植株进行DNA提取以备其VNTR微卫星分子标记鉴定,部分DNA提取情况见图1,其浓度符合鉴定所需的PCR模板浓度。
将提取的模板DNA按1.2.2中所列的PCR体系模式进行扩增,其电泳检测结果分别见图1-B,图1-C,图1-D。图1-B是在选育系列中趋于稳定并已进行扩繁的材料,其所用引物为TR1、TR3,标号1,2分别为保持系及对应不育系;图1-C是各类型受试植株TR1引物检测图;图1-D是各类型受试植株TR3引物检测图。选取能进行普遍鉴定甜菜Owen型胞质的两种引物TR1、TR3,并对已经趋于稳定符合育种标准的不育保持系进行模板对照检测。检测依据为对应引物下质基因特异性片段的重复数不同,其中Owen型不育系胞质的重复特异性片段重复数分别是TR1为4,TR3为2[6]。图1-C,1-D中标号1,2为稳定不育系及对应保持系,标号3~9则为调查结果中选取具代表性出粉情况的不育系植株。两种引物的检测结果有一定的统一性和差异性,表明了在甜菜田间育种过程中存在多种不同胞质类型的不育系体系,同时也显露出目前胞质鉴定引物的局限性。
表3为代表性不育系试验材料编号及对应VNTR鉴定结果。材料2为稳定保持系,鉴定结果为可育胞质,材料 4,5,6,7,9 则为 Owen 型不育胞质有粉植株,3为鉴定结果争议植株,8为Owen型不育胞质败育植株。

表3 代表性受试材料编号、出粉情况、胞质类型
2.3 对不同类型的CMS单株花序蛋白双向蛋白图谱比对分析
对受试植株中的参考组,提取其苗期叶片蛋白进行双向蛋白图谱Image Master 2D Platinum 5.0软件分析,并在参考组内不育系间及保持系间进行2DE蛋白图谱比对分析。结果发现,在选定的参考组内385个蛋白点图谱中发现差异点12个,在参考组内的保持系间及不育系间的多对比对发现蛋白差异点总和23个,并且包含前者发现的12个蛋白差异点。也就是说在苗期保持系与不育系的双向蛋白电泳图谱有较高的匹配率,并且此类差异能够在不育系间和保持系间重复显现。进一步得出无论甜菜的胞质基因还是核基因在育性相关上,在苗期的蛋白表达均不表现差异表达。之后又对编号3~9的7种类型差异表达,在甜菜植株雄蕊原基分化期的花蕾提取蛋白进行蛋白提取及双向蛋白电泳,各自的蛋白图谱同参考组中的不育图谱进行比对分析,再将其差异同参考组内的差异进行比对,其结果见图2。其中各不育系同参考不育系对比发现差异点数分别为 7,5,4,6,8,11,18 个;参考组间的差异蛋白点数为13个,在各个差异中所共存的差异蛋白点数2个。初步推测在雄蕊原基分化期甜菜育性基因已经参与调控表达,且质核基因导致的差异在表达调控上存在共同之处,但对最终表达的差异上又存在不同,导致了在性状上所表现的花药颜色、出粉程度上的不同。
3 讨论
整个试验首先在苗期对内蒙古农牧业科院特色作物所选育过程中的不育系及选育成型的不育保持系对进行VNTR分子标记检测;在受试材料花期中的雄蕊原基分化期花蕾进行取样预留;对花期花粉出粉情况及花药颜色进行田间调查统计;最后在田间花粉情况和分子标记检测的共同基础上将取样预留的花蕾分类进行2DE蛋白电泳,并将所得蛋白图谱通过Image Master 2D Platinum 5.0软件分析,得出甜菜质核基因在育性表达上的推测,为质核互作不育的机理探究指明方向。
3.1 甜菜不育系选育过程中的花粉情况分析
受试植株在花期除未出苗和未抽薹外共有38株在同一时期到达雄蕊原基分化期,其中分为:白败、黄败、不育I型白败、不育I型黄败、不育II型白败、不育II型黄败、不育黄色有粉7种类型,各自的植株数分别为 7,8,6,4,5,5,3 株。这就说明在内蒙古农牧业科学院特色作物所提供的选育中不育系核基因会分离出不同的类型,但对于前人提出甜菜质核互作双隐性核基因调控或是数量基因调控并不能得到推测认定。若想验证不育系核育性核基因类型还需要进一步的大量统计试验。
3.2 关于VNTR甜菜不育胞质特异序列的分类鉴定
对得到的38株重点植株进行已有引物TR1、TR3的PCR特异序列检测见图1-C,图1-D,其中对于前面提到的7种类型的检测结果显示前6种均属于Owen型不育胞质,对于不育有粉植株中存在Owen型保持系胞质见图1-D及表3,对比可以看出标号5同参考组中的保持系即标号2一致,属于育种过程中的混杂现象,编号9结合图1-D和表3发现其是拥有不育胞质的有粉植株。同时成功体现现有引物对Owen型甜菜不育系列鉴定上的参考价值,也为后期蛋白图谱比对过程中除去胞质基因调控引起差异的干扰。
3.3 7种类型间双向蛋白图谱比对分析讨论
对蛋白图谱存在的差异结果分析不难得出不育I,不育II在基因调控上同参考组不育系调控上存在一定的一致性,同时各自也有自身的特点,推测为核恢复基因在改变育性上的决定性作用,也可能是花粉颜色基因引起的差异。如要排除此类影响还需在对同一稳定系列甜菜品种的花药颜色进行参考对比。而对于不育有粉植株则均属于黄色花药,且不育有粉型存在的差异点不仅包含了对照组共有大多数差异,还出现了其他各类型均不存在的差异点5个,推测为全部核恢复基因共同调控的结果,并且是在质基因调控的基础上出现的核基因表达,而不是核恢复基因抑制质基因的表达,这同2004年在大豆中鉴定出的一些在乙烯合成,细胞程序化死亡,ATP合成,淀粉合成过程中起关键作用的蛋白在导致雄性不育的协作关系相似。
试验为甜菜不育蛋白质组学中差异蛋白质组学提出了研究方向和大胆推测,为后续对蛋白差异点的质谱分析、不育质核互作机理的完善做准备。
致谢:
感谢白晨老师对试验的指导意见和资金支持,感谢内蒙古农牧业科学院特色作物所提供的试验材料以及试验过程中的帮助,感谢李晓东、张辉等的试验指导。
参考文献:
[1]Schmutz J,Cannon S B,Schlueter,et al.Genome sequence of thepalaeopolyploidsoybean[J].Nature,2010,463:178-183.
[2]曹洪祥.甜菜M14品系花器官特异表达蛋白质的研究[D].大庆:黑龙江八一农垦大学,2009.
[3]牛 佳.甜菜细胞质雄性不育系及其保持系花期蛋白差异表达分析[D].哈尔滨:哈尔滨工业大学,2013.
[4]王有昭,程大有.中国甜菜主要品系细胞质育性相关片段的分子差异[D].哈尔滨:哈尔滨工业大学,2006.
[5]田自华,张子义,张剑锋,等.甜菜细胞质雄性不育系与其保持系线粒体DNA的RAPD分析[J].分子植物育种,2004(2):817-822.
[6]Ogura H.Studies on the new male-sterility in japanese radish with special reference to the utilisation of this sterility towars the practicalraising of hybrid seeds[J].Mem Fac Agric Kagoshima Univ,1968,6:39-78.
[7]Bonhomme S,Budar F,Ferault M,et al.A 2.5 Nco I fragment of Ogura radish mitochondrial DNA is correlated with cytoplasmic male sterility in Brassicacybrids[J].Curr Genet,1991,19:121-127.
[8]Bonhomme S,Budar F,Lancelin D,et al.Sequence and transcript analysis of the Nco2.5 Oguraspecific fragment correlated with cytoplasmic male sterility in Brassicacybri ds[J].Mol Gen Genet,1992,235:340-348.
[9]Grelon M,Budra F,Bonhomme S,etal.Ogura CMS associated orf138 istranslated into a mitochondrial membrane polypeptide in male-sterile Brassicacybrids[J].Mol Gen Genet,1994,243:540-547.
