自然降温对邓恩桉叶片MDA含量及SOD活性的影响
2016-04-13邓紫宇陈健波郭东强
邓紫宇,陈健波,郭东强
(1.广西壮族自治区林业科学研究院,广西南宁530002;2.国家林业局中南速生材繁育实验室,广西南宁530002;3.广西优良用材林资源培育重点实验室,广西南宁530002)
自然降温对邓恩桉叶片MDA含量及SOD活性的影响
邓紫宇1,2,3,陈健波1,2,3,郭东强1,2,3
(1.广西壮族自治区林业科学研究院,广西南宁530002;2.国家林业局中南速生材繁育实验室,广西南宁530002;3.广西优良用材林资源培育重点实验室,广西南宁530002)
为探明邓恩桉的耐寒机理,为其抗寒优良种源的选育与推广提供理论依据,以邓恩桉6个种源为试验材料,测定自然降温条件下邓恩桉叶片中丙二醛(MDA)含量及超氧化物岐化酶(SOD)活性。结果表明:在自然降温过程中,6个种源邓恩桉叶片中MDA含量随着温度的降低而升高,SOD活性随温度降低呈下降→上升→下降变化,但总体呈下降趋势,种源GED9509MDA含量的升高幅度与SOD活性的下降幅度均最小,在邓恩桉6个种源中抗寒性最强。
邓恩桉;丙二醛;超氧化物岐化酶;自然降温
桉树(Eucalyptus)是我国南方重要的速生丰产林树种,受寒冷气候条件的制约,在其分布北缘区发展缓慢。在我国广泛推广的桉树品种中,或因速生但不耐寒,或因耐寒但生长适应性差而限制桉树北移的速度[1]。邓恩桉(Eucalyptusdunnii)天然分布于澳大利亚新南威尔士州和昆士兰州,具有生长快、干形好等特点,在其自然分布区经历过-5.5℃的绝对低温,具有一定的抗寒性[2-3]。我国引种邓恩桉已有30多年的历史,在夏雨凉冷地区尤其中低海拔区域具有优良的适应性[4]。
形态指标、代谢指标、理化指标和生化指标是国内外评价林木抗寒性的主要指标,桉树抗寒性研究一直是当前的热点,主要集中在抗寒树种的选择、抗寒生长适应性调查及抗寒树种间抗寒能力的评价等方面[5-9],对已表现出一定抗寒适应性的树种种源/家系/个体间的抗寒机制与差异研究报道不多,人工低温胁迫下桉树抗寒生理生化指标变化研究较多[10-16],自然降温过程中桉树抗寒生理生化指标变化研究较少[17]。为此,笔者以间伐保留的邓恩桉6个种源为材料,对其部分生理生化指标进行测定,以期为抗寒优良种源的选育与推广提供理论依据与技术支撑。
1 材料与方法
1.1 试验材料与采样
以环江县华山林场雅龙分场2009年建立的邓恩桉种源试验林生长势相近的6个种源为试验材料(表1)。2014年10月至2015年3月每月初采取树冠中上部不同方向当年生侧枝的完整功能叶,每个种源采集15个单株,每株30枚,采后立即用锡纸包裹做好记录放入液氮中运回实验室。以2014年10月采样为对照。
1.2 指标测定
MDA含量的测定采用硫代巴比妥酸(TBA)比色法,SOD活性的测定采用NBT光照化学还原比色法[18-20]。
2 结果与分析
2.1 采样期间试验区的气候
2014年10月至2015年3月环江县华山林场雅龙分场邓恩桉种源试验林平均最高最低及平均温度如表2,全年最低温度出现在2015年1月,平均最低温3℃,极端低温-1℃。
2.2 自然降温对MDA含量的影响
MDA是生物膜膜脂过氧化最终产物,其含量是评价膜系统过氧化程度与受伤害程度的重要指标之一,二者呈正相关,MDA含量越高说明细胞质膜遭受破坏越严重,组织保护能力越弱。自然降温过程中,随着温度的不断下降,邓恩桉6个种源叶片中MDA含量不断升高,2015年1月达最大值。开春气温回升,MDA含量随着温度的升高而降低(表3)。邓恩桉6个种源MDA含量在自然降温过程中的变化趋势一致,但其增幅存在一定的差异。以2014年10月为对照,MDA含量达最大值时,种源GED9501、GED9502、GED9504、GED9506、GED9507和GED9509的增幅分别为64.01%、53.79%、55.91%、59.18%、58.77%和53.06%。MDA含量增幅与其抗寒力呈负相关,种源GED9509MDA含量增幅最小,抗寒力最强;种源GED9501MDA含量增幅最大,抗寒力最弱。以MDA含量为评价指标,邓恩桉6个种源的抗寒力依次为GED9509>GED9502>GED9504>GED9507>GED9506>GED9501。

表1 邓恩桉种源基本信息Table 1 Basic information ofE.dunniiprovenances
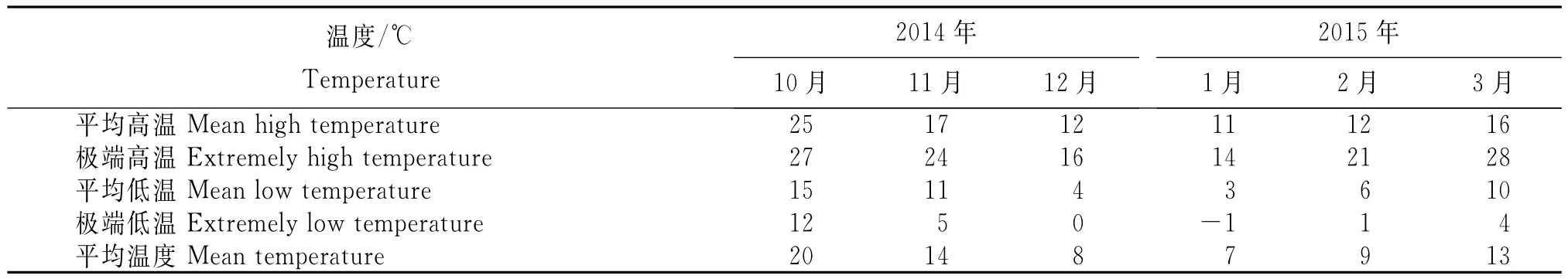
表2 试验地的温度条件Table 2 Temperature conditions in experiment place

表3 自然降温过程中邓恩桉6个种源MDA含量的变化Table 3 Change of MDA content of leaves in sixE.dunniiprovenances during the natural temperature decreasing process μmol/g
2.3 自然降温对SOD活性的影响
SOD是生物膜膜脂过氧化主要保护酶,其主要功能为催化活性氧歧化产物无毒氧和过氧化氢,从而达到保护植物遭受伤害。自然降温过程中,邓恩桉6个种源叶片中SOD活性呈下降→上升→下降的趋势,降温初期,SOD大体呈上升趋势以保护植株免受寒害,持续低温下,SOD活性明显下降,2015年1月达最小值。开春气温回升,植株恢复一定的抗寒能力,SOD活性逐渐上升(表4)。SOD活性是邓恩桉自身遗传因子决定的,SOD活性高低与其抗寒性强弱一致。在未遭受寒害时,种源GED9509SOD活性最大,说明其抗寒力最强,种源GED9501SOD活性最小,说明其抗寒力最弱。邓恩桉6个种源自然降温过程中SOD活性变化趋势一致,但其降幅存在一定的差异。以2014年10月为对照,SOD活性达最小值时,种源GED9501、GED9502、GED9504、GED9506、GED9507和GED9509的降幅分别为30.05%、19.66%、21.42%、21.76%、29.10%和19.41%。SOD活性下降率与植物品种本身抗寒能力强弱呈负相关,种源GED9509下降率最小,抗寒能力最强,种源GED9501SOD活性下降率最大,抗寒能力最弱。以SOD活性为评价指标,邓恩桉6个种源的抗寒力依次为GED9509>GED9502>GED9504>GED9506>GED9507>GED9501。
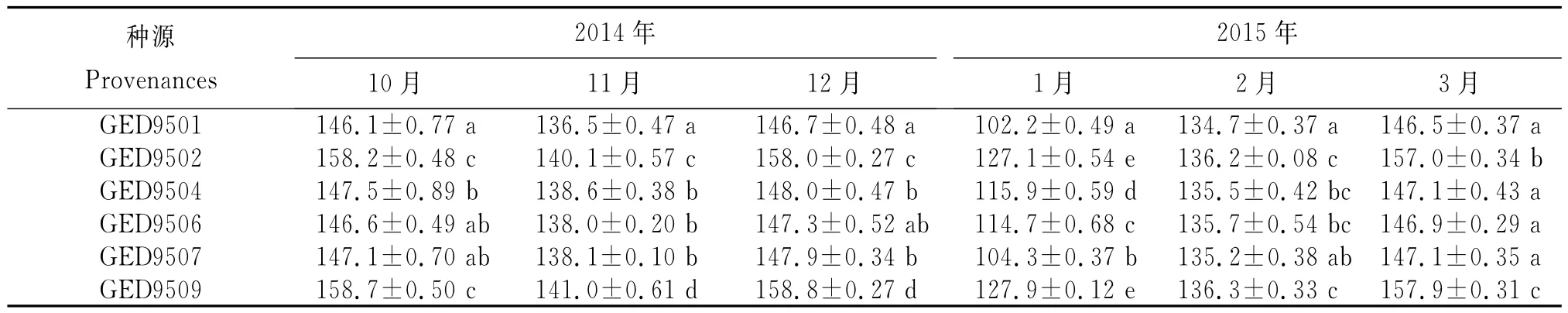
表4 自然降温过程中邓恩桉6个种源SOD活性的变化Table 4 Change of SOD activity of leaves in sixE.dunniiprovenances during the natural temperature decreasing process U/(min/g)
3 结论与讨论
3.1 样品的采集
样品的采集是试验成功与否的关键,由于试验地距离实验室路程较远,样品采集后不能及时处理,因此,正式采样前在实验室附近就地取样,对样品采取3种不同处理方式:不处理样品、湿布包裹样品和锡纸包裹样品置于液氮中。放置一段时间,测定MDA含量与SOD活性(以新采样品为对照),前2种处理方式MDA含量与SOD活性均出现较大波动,不适合长途样品采集处理,锡纸包裹样品置于液氮,样品MDA含量与SOD活性基本不变,适合长途样品采集的处理。如液氮不方便长途运输,也可采用样品采集后立即液氮处理,然后干冰运输。
3.2 MDA含量与邓恩桉抗寒性的相关性
MDA能与细胞内各种成分发生反应,导致生物膜的结构、生物功能分子、生理完整性的破坏。张学明[12]研究发现,不同桉树种类各季节及人工低温胁迫下其叶片MDA含量随着处理温度的降低而升高,在秋季转冬季期间,MDA含量也明显增加。黄月华等[13-14]研究表明,5种桉树苗期耐寒性随着温度的降低,MDA含量基本都高于对照,MDA含量高的品种抗寒性弱。刘建[15]研究发现,2种桉树对低温胁迫的响应机制与对照相比,低温胁迫促使其幼苗叶片MDA含量增加,随着时间的延长,MDA含量逐渐升高。靖长柏等[17]研究表明,自然低温下(日最低气温-3.6℃),3种桉树MDA含量相比对照明显增加。本研究中,邓恩桉6个种源在自然降温过程中叶片MDA含量随着温度下降而上升,随着温度回升而下降,与前人研究结论一致。MDA含量上升是因为邓恩桉受低温胁迫后,活性氧代谢平衡遭受破坏,抗氧化系统调节能力下降,活性氧过剩加剧膜脂过氧化作用,开春温度回升后,膜受损程度减弱,膜脂过氧化作用减少,MDA含量下降。
3.3 SOD活性与邓恩桉抗寒性的相关性
SOD酶作为植物体内有害物质的清除剂,可以清除过氧化物、过氧化氢和超氧自由基,减少羟基自由基。当植物受到逆境胁迫时,SOD酶可以起到保护植物细胞的作用[21]。黄月华等[13-14]研究表明,不同低温处理时5种桉树的SOD活性普遍低于对照,变化趋势基本一致,呈下降→上升→下降趋势。刘建[15]研究发现,连续3d夜间低温胁迫后,2种桉树幼苗叶片SOD活性均呈上升趋势,这是植株自身抵御寒害能力的体现,SOD活性水平的高低可以反映植株对某种逆境抵抗能力的大小。靖长柏等[17]研究表明,自然低温下,3种桉树SOD含量相比对照明显增加,低温胁迫期间,植株体内保护酶活性高,清除自由基能力强。郭祥泉[22]研究发现,不同低温处理,个体SOD活性在拐点前随着温度下降而上升,拐点后随着温度下降而急剧下降,SOD活性的拐点最低温度可确定为个体低温临界点。阚文靖等[16]研究表明,在25个桉树品种中抗寒能力强的桉树种类SOD活性高,抗寒能力弱的桉树种类SOD活性低。本研究中,邓恩桉6个种源在自然降温过程中叶片SOD活性随着温度的下降总体呈下降趋势,随着温度回升而升高,降温初期,SOD活性呈一定的上升趋势。说明,植株对低温具有一定的防御反应,以减缓伤害程度与速度,这是SOD活性在环境胁迫下保护功能的体现,持续低温下,膜脂过氧化程度加剧,植株体内酶促防御系统受到危害,活性氧未能及时清除而积累,SOD活性显著下降,开春温度回升后,膜脂过氧化程度减弱,植株体内酶促防御系统恢复,活性氧被及时清除,SOD活性上升,与前人研究结论相似。在此次自然降温过程中邓恩桉6个种源叶片SOD活性变化较平稳,-1℃的低温未对其造成严重伤害,与邓恩桉半致死温度在-8℃左右相符[11,15]。
3.4 邓恩桉抗寒性与地理分布的相关性
邓恩桉6个种源在自然降温过程中其抗寒性存在一定的差异,种源GED9509和GED9502抗寒力较强,2个种源引种地的纬度较高,在其自然分布区经受过相对较低温度的抗寒锻炼。海拔也是影响邓恩桉抗寒能力的因素之一,引种地海拔高的种源其抗寒力略强。因此,在今后引种选育速生抗寒桉树品种时,地理分布是必须考虑的因子。
3.5 提高植物抗寒性的措施
植物是一个开放的体系,低温能引起植物的生理障碍,使其受伤甚至死亡。植物抗寒力受其自身因素与外界环境的共同影响。自身因素由基因决定,可利用基因工程技术,将抗寒基因转入不抗寒或抗寒力弱的植株以提高其抗寒力。外界环境可一定程度上提高或降低植株的抗寒力,植物抗寒力的强弱与其生长速率呈负相关,植株生长速率快,其含水量高,组织疏松,呼吸旺盛,易受寒害;植株生长稳定,其含水量低,组织致密,呼吸速率适中,抗寒性较强。因此,低温来袭之前,合理施用P、K肥,少施N肥,不宜灌水,控制其生长,提高抗寒力。还可以喷施植物生长延缓剂或生长抑制剂等,延缓植株生长,提高其脱落酸含量,以提高其抗寒性能。此外,低温锻炼对提高植株的抗寒性具有一定的效果,可对种子或造林前的幼苗进行低温锻炼,以提高其抗寒能力[23]。
[1]罗建中.耐寒桉树良种——邓恩桉[J].桉树科技,2002(2):1-8.
[2]祁述雄.中国桉树[M].2版.北京:中国林业出版社,2002.
[3]TOM J,ROGER A,TREVOR B.邓恩桉在澳大利亚、中国和中南美洲的栽培候适宜性的确定[J].桉树科技,2001(1):55-60.
[4]郭东强,叶 露,周 维,等.2个种源邓恩桉木材纤维特性及变异[J].浙江农林大学学报,2014,31(4):502-507.
[5]刘友全,李志辉,马 英,等.速生耐寒邓恩桉在桉树分布北缘区的适应性[J].中南林业科技大学学报,2007,27(2):66-69.
[6]罗建中,曹加光,卢万鸿.邓恩桉等6种桉树的耐寒能力研究[J].桉树科技,2011,28(2):34-38.
[7]卢新瑛.2011年初春持续低温对邓恩桉生长的影响[J].亚热带农业研究,2011,7(2):100-104.
[8]郭祥泉.邓恩桉适应性栽植决策与抗寒-速生优株选择应用的研究[D].福州:福建农林大学,2009.
[9]庄国庆,胡天宇,郭洪英,等.不同种源地桉树抗寒性能的比较[J].桉树科技,2010,27(1):14-20.
[10]张 露,张俊红,温忠辉,等.引种桉树苗期的抗寒性分析[J].江西农业大学学报,2011,33(1):47-51.
[11]裘珍飞,曾炳山,刘 英,等.应用电导法评价5个桉树抗寒性[J].广东林业科技,2011,27(3):27-31.
[12]张学明.桉树在低温胁迫下的膜脂过氧化作用和膜伤害[J].经济林研究,1994(12):12-14.
[13]黄月华.五种桉树苗期耐寒性能的初步研究[D].海口:华南热带农业大学,2003.
[14]黄月华,徐建民,余雪标.低温胁迫对桉树代谢的影响[J].热带农业科学,2005,25(5):24-28.
[15]刘 建.两种桉树对低温胁迫的响应机制研究[D].南京:南京林业大学,2008.
[16]阚文靖,朱宁华,邱运亮.桉树对低温反应的生理生态学基础[J].经济林研究,1994(12):1-6.
[17]靖长柏,张利阳,童再康,等.自然低温胁迫对3种桉树生理生化特性的影响[J].防护林科技,2010(5):18-19,35.
[18]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[19]皱 琦.植物生理学实验指导[M].北京:中国农业出版社,2000.
[20]高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006.
[21]马旭俊,朱大海.植物超氧化物岐化酶研究进展[J].遗传,2003,25(2):225-231.
[22]郭祥泉.用膜透性、SOD酶活性和膜脂脂肪酸成分探讨林木抗寒性[J].西南林业大学学报,2012,32(4):6-11.
[23]潘瑞炽.植物生理学[M].5版.北京:高等教育出版社,2006.
(责任编辑:刘 海)
Influence of Natural Temperature Decrease on MDA and SOD Contents inEucalyptusdunniiLeaves
DENG Ziyu1,2,3,CHEN Jianbo1,2,3,GUO Dongqiang1,2,3
(1.GuangxiForestryScienceResearchInstitute,Nanning,Guangxi530002;2.KeyLaboratoryofCentralSouth Fast-growingTimberCultivationofStateForestryAdministrationofChina,Nanning,Guangxi530002;3.Guangxi KeyLaboratoryofSuperiorTimberTreesResourceCultivation,Nanning,Guangxi530002,China)
In order to studying on investigate the cold-resistant mechanism ofE.dunnii,to provide theoretical references for its excellent cold-resistant seedingsource breeding and spred the malondiadehyde(MDA)content and activity of superoxide dismutase(SOD)were investigated inE.dunniileaves from six provenances.Results:MDA content increased with temperature decreasing,SOD activity showed a change trend of decreasing at first then increasing and decreasing at last,while it decreased in general.The smallest amplitude of change is provenance GED9509during the natural temperature decrease,the cold resistance of provenance GED9509was stronger than other provenances.
Eucalyptusdunnii;MDA;SOD;natural temperature decrease
S792.39
A
2015-12-09;2016-07-31修回
广西林业科学院基本科研业务费项目“邓恩桉保护酶活性与抗寒性关系研究”(林科201408);广西优良用材林资源培育重点实验室自主项目“邓恩桉遗传多样性的AFLP分析”(13-A-01-02)
邓紫宇(1984-),男,工程师,硕士,从事林木遗传育种研究。E-mail:365204914@qq.com
1001-3601(2016)09-0375-0033-04
