Developing high quality chemical probes targeting ubiquitin-specific proteases
2016-02-16YufengTONG
Yu-feng TONG
(1.Structural Genomics Consortium,Toronto,Ontario,Canada;2.Department of Pharmacology and Toxicology,University of Toronto,Toronto,Ontario,Canada)
·COMMENTARY·
Developing high quality chemical probes targeting ubiquitin-specific proteases
Yu-feng TONG1,2
(1.Structural Genomics Consortium,Toronto,Ontario,Canada;2.Department of Pharmacology and Toxicology,University of Toronto,Toronto,Ontario,Canada)
Ubiquitin-specific proteases(USPs)regulate the deconjugation of ubiquitin tag from substrate proteins.Dysregulation of USPs has been linked to many diseases.Despite extensive efforts by academia and industry,high quality chemical molecules targeting the USPs family of ubiquitin enzymes are still scarce.In this commentary,I propose the criteria of a high quality chemical probe and the strategies to target USPs.
chemical probes;drug discovery;small molecule inhibitors;ubiquitin-specific proteases
1 UBIQUITINSIGNALINGPATHWAY AND DRUG DISCOVERY
Ubiquitination is a highly dynamic and reversible post-translationalmodification that conjugates a well-conserved small regulatory protein,ubiquitin,to the lysine side chain of a substrate protein through an isopeptide.It is catalyzed by a multi-enzyme cascade comprising ubiquitin-activating enzymes(E1), ubiquitinconjugating enzymes(E2),and ubiquitin ligases (E3).The ε-amino group of the side chain of sev⁃en lysines(K6,K11,K27,K29,K33,K48,and K63),and the α-amino group of the N-terminal methionine(M1)in the ubiquitin,can also serve as sites for the covalent attachment of another ubiquitin molecule,thus forming polyubiquitin chain of different linkages[1-2].A single or a chain of multiple ubiquitins can be deposited on a target protein.The size and the nature of the linkage of the polyubiquitin chain have consequences in determining the fate of the tagged proteins. The classical K48-linked polyubiquitination was discovered in proteasome-mediated protein degradation[3]and was most well-studied.In recent years,mono-ubiquitination and atypical polyubiq⁃uitin modification were shown to play a plethora of important cellular functional roles involving DNA damage repair,immune response,gene transcription,receptor endocytosis and trafficking, and cell signaling[4-5]and are gaining increasing interest.
The conjugated ubiquitin can be removed by a family of specialized enzymes known as deubiquitinases(DUBs).The human genome encodes about 100 DUBs[6-7]that can be divided into five subfamilies based on sequence homology, which consist of four cysteine proteases:ubiquitinspecific proteases(USPs),ubiquitin carboxyterminal hydrolases(UCHs),ovarian-tumor proteases (OTUs),Machado-Joseph disease protein domain proteases(MJDs),and one JAMM/MPN domainassociated metallopeptidases(JAMMs). With about 57 members,the USPs comprise the largest and most diverse subfamily of DUBs.Most USPs are multiple domain proteins containing a core catalytic domain of about 350 a.a.,which is often bifurcated by one or two insertions of 40-600 a.a. size.The intricate architecture of USPs indicates the function of this family of proteases is delicatelyregulated.
Since ubiquitination is involved in almost every biological process in the cell,dysregulation of ubiquitination has been linked to many diseases. Over the last decade,mutation,aberrant expression, or mislocalization of increasing number of USPs have been linked to cancers,neurological disorders, infertility,viral infection,auto-immune diseases(Tab.1).Latestexamples include the whole exome sequencing studies[8-11]of Cushing′s disease, which discovered thatsomatic mutations of USP8 lead to a hyper-activated USP8 proteolytic fragment.Hyper-activation of USP8 upregulates epidermal growth factor receptors(EGFRs)level and finally leads to the overproduction of adreno⁃corticotropic hormone.In another study,mutation of USP53 was found to cause progressive hearing loss[12].

Tab.1 Disease association of ubiquitin-specific proteases (USP)
Since the FDA approval of two proteasome inhibitors,bortezomib[13]and carfilzomib[14]in anticancer therapy,the ubiquitin-proteasome pathway has gain tremendous interest from both academia and industry to search for inhibitors that can selectively modulate the activity of the ubiquitination regulators.Both bortezomib and carfilzomib inhibit the chymotrypsin-like activity of the 20S proteasome and have relatively narrow therapeutic index.Inhibitors of ubiquitin regulatory enzymes (E1s,E2s,E3s or DUBs)hold the promise of better effectiveness and fewer non-specific side effects compared with proteasome inhibitors.It was also proposed thatthe ubiquitinationdeubiquitination process is analogous to the phosphorylation-dephosphorylation process and could furnish as many drug targets as protein kinases[15].The DUBs are believed to be more druggable compared to E3 ubiquitin ligases as the latterlacks defined catalytic residues[16]. Several recent reviews have summarized published DUB inhibitors[16-17]in the public domain and in patents[18-20].
Despite the progress,only one dual-specificity inhibitor targeting both UCHL5 and USP14 of the human USPs family has advanced to clinical trial to our knowledge[21].Based on our experience in the validation of literature reported USP inhibi⁃tors and an on-going compound screening campaign targeting USP proteins,we feel high quality molecular probes are needed to explore the functional roles of USPsin vivoand to vali⁃date whether USPs are tractable therapeutic tar⁃gets for disease treatment.
2 CHEMICAL PROBES CRITERIA AND REALITY IN UBIQUITIN REGULATOR INHIBITOR
The dramatic stories of GlaxoSmithKline′s failed$720M investment on resveratrol,and the downfall of iniparib as a poly(ADP-ribose)poly⁃merase(PARP)inhibitor in phaseⅢ clinical trial[22]remind us of the importance of data reproducibili⁃ty[23-24]and the need of well-characterized,high quality chemical compounds in preclinical drug discovery to understand the biological function of target proteins.First established in 2009 by the Structural Genomics Consortium(SGC),the con⁃cept of a″chemical probe″has now gained wide acceptance in the chemical biology research community as a standard for a high-quality small molecule compound for target validation and for biological function studies[25].Chemical probes are defined as drug-like small molecules that meet the following four criteria:(a)potency:potent inhibition of the target activity with an IC50< 100 nmol·L-1in anin vitrobiochemical assay; (b)selectivity:selective(>30 fold)inhibition of the target protein(or small group of proteins) relative to other members of the same protein family(for example the deubiquitinase family); (c)cellular activity:on-target activity demonstrated in cells at<1 μmol·L-1;(d)toxicity:low or no off-target cellular toxicity.A three-dimensional high-resolution complex structure,mostly deter⁃mined using crystallography,of the target protein bound with the compound,is the holy grail of a chemical probe,which gives an intuitive view of target engagement.Chemical probes are intended for use in cellular studies to link pharmacological inhibition of a target with a phenotypic change, such as modulation of a disease-related biology. They are complementary to genetic methods, such as miRNA,siRNA,or CRISPR/CRISPR associated protein 9,for target validation.
Chemical probes have a significant impact on basic and translational sciences.For basic science,chemical probes are complementary to genetic methods of target validation.They are mostly reversible,inexpensive to purchase and relatively easy to use.For translational science, chemical probes demonstrate that the target is″druggable″and show that pharmacological inhibition of the target,instead of genetic ablation of the entire protein,leads to the modification of diseaserelated pathway at a cellular level.Furthermore, majority,if not all,of the proteins are engaged in a dynamic protein-protein interaction network in the celland often play multiple functionalroles besides their enzymatic activities.Genetic ablation of the entire protein,or even a domain,may introduce unwanted effects and may not reveal the genuine link between phenotype and genotype. Among all the target validation approaches available today,pharmacological perturbation using small molecule compounds remains the most relevant to industry.
The field of ubiquitin enzyme inhibitors are plagued with low quality chemical compounds that do not survive scrutiny,yet such studies were often cited and the compounds were used in follow-up studies,which will eventually lead to irreproducibility of the results or a complete reversal of previous conclusion.In one case of E3 ubiquitin ligases,menadione,commonly known as vitamin K3,was claimed to be a specific inhibitor for Sevenin absentiaHomolog 2(SIAH2)E3 ubiquitin ligase at an IC50of 20 μmol·L-1based on an electrochemiluminescent ubiquitination assay[26].Following this work,menadione was used in several high profile studies[27-31]to investigate the function of SIAH2in vivo.Actually,menadione is a cysteine alkylator[32]that covalently modifies free cysteine of the proteins and could thus destabilize substrate proteins nonspecifically(Zhang,et al,manuscript submitted).It belongs to the so-called pan-assay interference compounds(PAINS)[33]that should be avoided in preclinical drug discovery at all cost.
In another case of USPs,spautin-1 was found to be a potent inhibitor for USP10 and USP13 at an IC50of 0.6-0.7 μmol·L-1in a high profile publication[34].However,we were unable to reproduce the inhibitory activity of spautin-1 against USP10 using ubiquitin-AMC assay(Tong,et al,unpublished result).From a structural biology point of view,the USP13 has an insertion of about 180 a.a.within the catalytic domain while the USP10 has none,and the two USPs have very low sequence identity and different residue compositions around the catalytic sites.It is highly unlikely that spautin-1 would inhibit these two distant USPs at a similar IC50.While spautin-1 and its derivatives[34]may still have the potency to inhibit autophagy at a cellular level,our obser⁃vation calls for a more careful and detailed char⁃acterization of the compounds using orthogonal validation methods before they were reported.
3 DRUGGABILITY OF THE USP FAMILY
The USP family of DUBs are different from other enzymes,such as methyltransferases and protein kinases,that take metabolites or peptides as substrates.USPs recognize the whole ubiquitin protein as a substrate and the substrate binding pocket is relatively flat and shallow(Fig.1).To date,no crystal structure of a human USP bound with a chemical compound has been released in the public domain,although a co-crystal structure of USP7 with a potent inhibitor has been reported in a conference[35].The field is in dire need of more well-validated and proven inhibitors to demonstrate that the USP family is druggable[16].
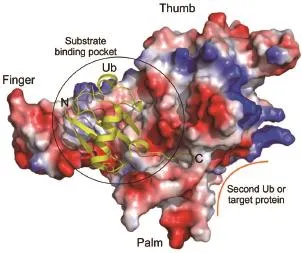
Fig.1 Typicalsubstrate binding mode ofUSP catalytic domain,exemplified by USP7/Ub complex (PDB:1NBF).USP7 is shown in surface charge representa⁃tion,where blue represents positive charges and red for nega⁃tive charges.The catalytic domain can be divided into finger, thumb,and palm regions.USPs have a large and shallow sub⁃strate binding pocket.
While most of the current compound screening campaigns target the catalytic domains of USPs, the USPs often require binding partners or auxiliary domains for its full function,which provides an opportunity for developing allosteric regulators[36]. For example,the full length USP7 is much more active than the catalytic domain,yet guanosine monophosphate synthetase(GMPS)binding to the ubiquitin-like domains of USP7 further hyperactivates its DUB activity[37].An active USP22 requiresthe formation ofa mandatory fourcomponent complex[38-39].USP1,USP12,and USP46 require WD40-repeat(WDR)proteins for their full activities[40-42].Indeed,the current best-in-class USP inhibitor,ML-323,allosterically inhibits the activity of the USP1/WDR48 complex[43]at an IC50of around 70 nmol·L-1.The WDR proteins have a donut-shaped structuralfold and well-defined substrate binding pockets.A chemical probe has been developed recently targeting the substrate binding pocket of WDR5[44]and shown activity towards the WDR5-MLL complex[45].As many USP proteins forms complex with WDR proteins, these complexes present potentially more tractable targets for chemical probe development[46].
It is noteworthy that we and the Sidhu lab at the University of Toronto have shown a phage display-based technology can be utilized to develop ubiquitin-based biological probes to target ubiquitin enzymes[47-48].These ubiquitin variants(UbVs) are highly potent and selective,and can be expressed in the cell to modulate the function of the cognate ubiquitin enzymes.While introduction of UbVs into the cell as a therapeutic reagent is still very challenging,these biological probes can be easily used for target validation and to explore the new biology of ubiquitin enzymes[48].
4 CONCLUSIONS AND PROSPECTIVE
Almost four decades passed since the first description of ubiquitination,fascinating new biology of ubiquitin are still being discovered occasionally. The latest big story is the phosphorylation of ubiquitin and its involvementin Parkinsondisease[49-51].Phosphorylation of ubiquitin adds another layer of regulation onto the already sophisticated network of ubiquitination,suggesting new challenges and opportunities to characterize the function and therapeutic potential of targeting ubiquitin regulatory enzymes.Interestingly,10 out of 12 tested USPs have impaired activity towards Ser65-phosphorylated polyubiquitin chain substrates[49],suggesting ubiquitin phosphorylation has repercussions for ubiquitination and deubiq⁃uitination cascades beyond Parkin E3 ligase activation.Other less well-studied USPs may play important roles in regulating the function of phosphorylated ubiquitin.Currently,only a handful of crystal structures of the human USP catalytic domains have been solved and no USP-compound complex structure has been released in the public domain.The USPs provide both opportunities and challenges to develop rigorously characterized, well-defined,and highly quality chemical probes to explore to the therapeutic potentials of these enzymes in diseases.
ACKNOWLEDGMENTS:The StructuralGenomics Consortium is a registered charity(number 1097737)that receives funds from AbbVie,Bayer Pharma AG,Boeh⁃ringer Ingelheim,Canada Foundation for Innovation, Eshelman Institute forInnovation,Genome Canada through Ontario Genomics Institute,Innovative Medicines Initiative(EU/EFPIA)〔ULTRA-DD grantno.115766〕Janssen,Merck&Co.,Novartis Pharma AG,Ontario Ministry of Economic Development and Innovation,Pfizer, São Paulo Research Foundation-FAPESP,Takeda,and the Wellcome Trust.Y.T.is also jointly supported by Janssen and the Centre for Collaborative Drug Research through the Neuroscience Catalyst program.
REFERENCES:
[1] Kirisako T, Kamei K, MurataS,KatoM,Fukumoto H,Kanie M,et al.A ubiquitin ligase complex assembles linear polyubiquitin chains[J].EMBO J,2006,25(20):4877-4887.
[2] Komander D,Rape M.The ubiquitin code[J]. Annu Rev Biochem,2012,81:203-229.
[3] Varshavsky A.The early history of the ubiquitin field[J].Protein Sci,2006,15(3):647-654.
[4] Ikeda F,Dikic I.Atypical ubiquitin chains:new molecular signals.′Protein Modifications:Beyond the Usual Suspects′review series[J].EMBO Rep,2008,9(6):536-542.
[5] Kulathu Y,Komander D.Atypical ubiquitylationthe unexplored world of polyubiquitin beyond Lys48 and Lys63 linkages[J].Nat Rev Mol Cell Biol,2012,13(8):508-523.
[6] Komander D.The emerging complexity of protein ubiquitination[J].Biochem Soc Trans,2009,37(Pt 5):937-953.
[7] Komander D.Mechanism,specificity and struc⁃ture of the deubiquitinases[M]//Groettrup M,ed. Conjugation and Deconjugation of Ubiquitin Family Modifiers.Springer,New York,2010:69-87.
[8] Ma ZY,Song ZJ,Chen JH,Wang YF,Li SQ,Zhou LF,et al.Recurrent gain-of-function USP8 mutations in Cushing′s disease[J].Cell Res,2015,25(3):306-317.
[9] Reincke M,Sbiera S,Hayakawa A,Theodoro⁃poulou M,Osswald A,Beuschlein F,et al.Muta⁃tions in the deubiquitinase gene USP8 cause Cushing′s disease[J].Nat Genet,2015,47(1):31-38.
[10] Theodoropoulou M,Reincke M,Fassnacht M,Komada M.Decoding the genetic basis of Cushing′s disease:USP8 in the spotlight[J].Eur J Endocrinol,2015,173(4):M73-M83.
[11] Perez-Rivas LG,Theodoropoulou M,Ferraù F,Nusser C,Kawaguchi K,Stratakis CA,et al. The gene of the ubiquitin-specific protease 8 is frequently mutated in adenomas causing Cushing′s disease[J].J Clin Endocrinol Metab,2015,100(7):E997-E1004.
[12] Kazmierczak M, Harris SL,Kazmierczak P,Shah P,Starovoytov V,Ohlemiller KK,et al.Pro⁃gressive hearing loss in mice carrying a mutation in Usp53[J].J Neurosci,2015,35(47):15582-15598.
[13] Bignell GR,Warren W,Seal S,Takahashi M,Rapley E,Barfoot R,et al.Identification of the familial cylindromatosis tumour-suppressor gene[J].Nat Genet,2000,25(2):160-165.
[14] Nijman SM,Huang TT,Dirac AM,Brummelkamp TR,Kerkhoven RM,D′Andrea AD,et al.The deubiquitinating enzyme USP1 regulates the Fanconi anemia pathway[J].Mol Cell,2005,17(3):331-339.
[15] Kim J,Kim WJ,Liu Z,Loda M,Freeman MR. The ubiquitin-specific protease USP2a enhancestumor progression by targeting cyclin A1 in bladder cancer[J].Cell Cycle,2012,11(6):1123-1130.
[16] Cleynen I,Vazeille E,Artieda M,Verspaget HW,Szczypiorska M,Bringer MA,et al.Genetic and microbial factors modulating the ubiquitin protea⁃some system in inflammatory bowel disease[J]. Gut,2014,63(8):1265-1274.
[17] Hou X,Wang L,Zhang L,Pan X,Zhao W.Ubiq⁃uitin-specific protease 4 promotes TNF-α-induced apoptosis by deubiquitination of RIP1 in head and neck squamous cell carcinoma[J].FEBS Lett,2013,587(4):311-316.
[18] García-Caballero A,Gadotti VM,Stemkowski P,Weiss N,Souza IA,Hodgkinson V,et al.The deubiquitinating enzyme USP5 modulates neuro⁃pathic and inflammatory pain by enhancing Cav3.2 channel activity[J].Neuron,2014,83(5):1144-1158.
[19] Oliveira AM,Chou MM.The TRE17/USP6 onco⁃gene:a riddle wrapped in a mystery inside an enigma[J].Front Biosci(Schol Ed),2012,4(4):321-334.
[20] Bhattacharya S,Ghosh MK.HAUSP,anovel deubiquitinase for Rb-MDM2 the critical regulator[J].FEBS J,2014,281(13):3061-3078.
[21] Byun S,Lee SY,Lee J,Jeong CH,Farrand L,Lim S,et al.USP8 is a novel target for overcoming gefitinib resistance in lung cancer[J].Clin Cancer Res,2013,19(14):3894-3904.
[22] Homan CC,Kumar R,Nguyen LS,Haan E,Raymond FL,Abidi F,et al.Mutations in USP9X are associated with X-linked intellectual disability and disrupt neuronal cell migration and growth[J].Am J Hum Genet,2014,94(3):470-478.
[23] Yang Q,Ou C,Liu M,Xiao W,Wen C,Sun F. NRAGE promotes cell proliferation by stabilizing PCNA in a ubiquitin-proteasome pathway in esophageal carcinomas[J].Carcinogenesis,2014,35(7):1643-1651.
[24] Wu HC,Lin YC,Liu CH,Chung HC,Wang YT,Lin YW,et al.USP11 regulates PML stability to control Notch-induced malignancy in brain tumours[J].Nat Commun,2014,5:3214.
[25] Burkhart RA,Peng Y,Norris ZA,Tholey RM,Talbott VA,Liang Q,et al.Mitoxantrone targets human ubiquitin-specific peptidase 11(USP11)and is a potent inhibitor of pancreatic cancer cell survival[J].Mol Cancer Res,2013,11(8):901-911.
[26] Mcclurg UL,Harle VJ,Nabbi A,Batalha-Pereira A, Walker S,Coffey K,et al.Ubiquitin-specific protease 12 interacting partners Uaf-1 and WDR20 are potential therapeutic targets in prostate cancer[J]. Oncotarget,2015,6(35):37724-37736.
[27] Geng J,Huang X,Li Y,Xu X,Li S,Jiang D,et al. Down-regulation of USP13 mediates phenotype transformation of fibroblasts in idiopathic pulmo⁃nary fibrosis[J].Respir Res,2015,16:124.
[28] Ortuno D,Carlisle HJ,Miller S.Does inactivation of USP14 enhance degradation of proteasomal substrates that are associated with neurodegener⁃ative diseases?[J].F1000Res,2016,5:137.
[29] WilsonSM,vBhattacharyyaB,Rachel RA,Coppola V,Tessarollo L,Householder DB,et al. Synaptic defects in ataxia mice result from a mutation in Usp14,encoding a ubiquitin-specific protease[J].Nat Genet,2002,32(3):420-425.
[30] Vos RM,Altreuter J,White EA,Howley PM.The ubiquitin-specific peptidase USP15 regulates human papillomavirus type 16 E6 protein stability[J].J Virol,2009,83(17):8885-8892.
[31] Adorno M,Sikandar S,Mitra SS,Kuo A,Nicolis Di Robilant B,Haro-Acosta V, etal.Usp16 contributes to somatic stem-cell defects in Down′s syndrome[J].Nature,2013,501(7467):380-384.
[32] Mcfarlane C,Mcfarlane S,Paul I,Arthur K,Scheaff M,Kerr K,et al.The deubiquitinating enzyme USP17 is associated with non-small cell lung cancer(NSCLC)recurrence and metastasis[J].Oncotarget,2013,4(10):1836-1843.
[33] Pereg Y,Liu BY,O'rourke KM,Sagolla M,Dey A,Komuves L,et al.Ubiquitin hydrolase Dub3 promotes oncogenic transformation by stabilizing Cdc25A[J].Nat Cell Biol,2010,12(4):400-406.
[34] LiuLQ,IlariaR,KingsleyPD,Iwama A,Van Etten RA,Palis J,et al.A novel ubiquitinspecific protease,UBP43,cloned from leukemia fusion protein AML1-ETO-expressing mice,func⁃tions in hematopoietic cell differentiation[J].Mol Cell Biol,1999,19(4):3029-3038.
[35] Altun M,Zhao B,Velasco K,Liu H,Hassink G,Paschke J,et al.Ubiquitin-specific protease 19(USP19)regulates hypoxia-inducible factor 1α(HIF-1α)during hypoxia[J].J Biol Chem,2012,287(3):1962-1969.
[36] Li Z,Wang D,Na X,Schoen SR,Messing EM,Wu G.Identification of a deubiquitinating enzyme subfamily as substrates of the von Hippel-Lindau tumor suppressor[J].Biochem Biophys ResCommun,2002,294(3):700-709.
[37] Fan Y,Mao R,Yu Y,Liu S,Shi Z,Cheng J,et al. USP21 negatively regulates antiviral response by acting as a RIG-Ⅰdeubiquitinase[J].J Exp Med,2014,211(2):313-328.
[38] Hu J,Yang D,Zhang H,Liu W,Zhao Y,Lu H,et al.USP22 promotes tumor progression and induces epithelial-mesenchymal transition in lung adenocarcinoma[J].Lung Cancer,2015,88(3):239-245.
[39] Oliveira SA,Li YJ,Noureddine MA,Zuchner S,Qin X,Pericak-Vance MA,et al.Identification of risk and age-at-onset genes on chromosome 1p in Parkinson disease[J].Am J Hum Genet,2005,77(2):252-264.
[40] Li J,Tan Q,Yan M,Liu L,Lin H,Zhao F,et al. miRNA-200c inhibits invasion and metastasis of human non-small cell lung cancer by directly targeting ubiquitin specific peptidase 25[J].Mol Cancer,2014,13:166.
[41] Stouffs K,Lissens W,Tournaye H,Van Steirteghem A,Liebaers I.Possible role of USP26 in patients with severely impaired spermatogenesis[J].Eur J Hum Genet,2005,13(3):336-340.
[42] Popov N,Wanzel M,Madiredjo M,Zhang D,Beijersbergen R, Bernards R, etal.The ubiquitin-specific protease USP28 is required for MYC stability[J].Nat Cell Biol,2007,9(7):765-774.
[43] Bingol B,Tea JS,Phu L,Reichelt M,Bakalarski CE,Song Q,et al.The mitochondrial deubiquitinase USP30 opposes parkin-mediated mitophagy[J]. Nature,2014,510(755):370-375.
[44] Akhavantabasi S,Akman HB,Sapmaz A,Keller J,Petty EM,Erson AE.USP32 is an active,membranebound ubiquitin protease overexpressed in breast cancers[J].Mamm Genome,2010,21(7-8):388-397.
[45] Yuasa-Kawada J,Kinoshita-Kawada M,Rao Y,Wu JY.Deubiquitinating enzyme USP33/VDU1 is required for Slit signaling in inhibiting breast cancer cell migration[J].Proc Natl Acad Sci USA,2009,106(34):14530-14535.
[46] Pan J,Deng Q,Jiang C,Wang X,Niu T,Li H,et al.USP37 directly deubiquitinates and stabilizes c-Myc in lung cancer[J].Oncogene,2015,34(30):3957-3967.
[47] Wang H,Ji X,Liu X,Yao R,Chi J,Liu S,et al. Lentivirus-mediated inhibition of USP39 suppresses the growth of breast cancer cells in vitro[J].Oncol Rep,2013,30(6):2871-2877.
[48] Li Y,Schrodi S,Rowland C,Tacey K,Catanese J,Grupe A.Genetic evidence for ubiquitin-specific proteasesUSP24 and USP40 ascandidate genes for late-onset Parkinson disease[J].Hum Mutat,2006,27(10):1017-1023.
[49] Hou K,Zhu Z,Wang Y,Zhang C,Yu S,Zhu Q,et al.Overexpression and biological function of ubiquitin-specific protease 42 in gastric cancer[J].PLoS One,2016,11(3):e0152997.
[50] Zhang Y,Foreman O,Wigle DA,Kosari F,Vasmatzis G,Salisbury JL,et al.USP44 regulates centrosome positioning to prevent aneuploidy and suppress tumorigenesis[J].J Clin Invest,2012,122(12):4362-4374.
[51] Zhang Y,Van Deursen J,Galardy PJ.Overex⁃pression of ubiquitin specific protease 44(USP44)induces chromosomal instability and is frequently observed in human T-cell leukemia[J]. PLoS One,2011,6(8):e23389.
[52] Tomida S,Mamiya T,Sakamaki H,Miura M,Aosaki T,Masuda M,et al.Usp46 is a quantitative trait gene regulating mouse immobile behavior in the tail suspension and forced swimming tests[J]. Nat Genet,2009,41(6):688-695.
[53] Zhang B,Yin Y,Hu Y,Zhang J,Bian Z,Song M,et al.MicroRNA-204-5p inhibits gastric cancer cell proliferation by downregulating USP47 and RAB22A[J].Med Oncol,2015,32(1):331.
[54] Figueroa ME, Lugthart S, LiY, Erpelinck-Verschueren C,Deng X,Christos PJ,et al.DNA methylation signatures identify biologically distinct subtypes in acute myeloid leukemia[J].Cancer Cell,2010,17(1):13-27.
[55] Atanassov BS,Mohan RD,Lan X,Kuang X,Lu Y,Lin K,et al.ATXN7L3 and ENY2 coordinate activity of multiple H2B deubiquitinases important for cellular proliferation and tumor growth[J].Mol Cell,2016,62(4):558-571.
[56] Bross PF,Kane R,Farrell AT,Abraham S,Benson K,Brower ME,et al.Approval summary for bortezomib for injection in the treatment of multiple myeloma[J].Clin Cancer Res,2004,10(12 Pt 1):3954-3964.
[57] Thompson JL.Carfilzomib:a second-generation proteasome inhibitor for the treatment of relapsed and refractory multiple myeloma[J].Ann Pharmacother,2013,47(1):56-62.
[58] Cohen P,Tcherpakov M.Will the ubiquitin system furnish as many drug targets as protein kinases?[J].Cell,2010,143(5):686-693.
[59] D′Arcy P,Wang X,Linder S.Deubiquitinase inhibition as a cancer therapeutic strategy[J]. Pharmacol Ther,2015,147:32-54.
[60] Ndubaku C,Tsui V.Inhibiting the deubiquitinating enzymes(DUBs)[J].J Med Chem,2015,58(4):1581-1595.
[61] FarshiP, DeshmukhRR,NwankwoJO,Arkwright RT,Cvek B,Liu J,et al.Deubiquitinases(DUBs)and DUB inhibitors:a patent review[J]. Expert Opin Ther Pat,2015,25(10):1191-1208.
[62] Kessler BM.Selective and reversible inhibitors of ubiquitin-specific protease 7:a patent evaluation(WO2013030218)[J].Expert Opin Ther Pat,2014,24(5):597-602.
[63] Guédat P,Colland F.Patented small molecule inhibitors in the ubiquitin proteasome system[J]. BMC Biochem,2007,8(Suppl 1):S14.
[64] Sheridan C.Drug makers target ubiquitin protea⁃some pathway anew[J].Nat Biotechnol,2015,33(11):1115-1117.
[65] Sinha G.Downfall of iniparib:a PARP inhibitor that doesn′t inhibit PARP after all[J].J Natl Cancer Inst,2014,106(1):djt447.
[66] Frye SV,Arkin MR,Arrowsmith CH,Conn PJ,Glicksman MA,Hull-Ryde EA,et al.Tackling reproducibility in academic preclinical drug discovery[J].Nat Rev Drug Discov,2015,14(11):733-734.
[67] Dolgin E.Drug discoverers chart path to tackling data irreproducibility[J].Nat Rev Drug Discov,2014,13(12):875-876.
[68] Arrowsmith CH,Audia JE,Austin C,Baell J,Bennett J,Blagg J,et al.The promise and peril of chemical probes[J].Nat Chem Biol,2015,11(8):536-541.
[69] ShahM,StebbinsJL,DewingA,QiJ,Pellecchia M,Ronai ZA.Inhibition of Siah2 ubiquitin ligase by vitamin K3(menadione)attenuates hypoxia and MAPK signaling and blocks melanoma tumorigenesis[J].Pigment Cell Melanoma Res,2009,22(6):799-808.
[70] Ma B,Chen Y,Chen L,Cheng H,Mu C,Li J,et al.Hypoxia regulates Hippo signalling through the SIAH2 ubiquitin E3 ligase[J].Nat Cell Biol,2015,17(1):95-103.
[71] Baba K,Morimoto H,Imaoka S.Seven in absentia homolog 2(Siah2)protein is a regulator of NFE2-related factor 2(Nrf2)[J].J Biol Chem,2013,288(25):18393-18405.
[72] Sun RC,Denko NC. Hypoxicregulation of glutamine metabolism through HIF1 and SIAH2 supports lipid synthesis that is necessary for tumor growth[J].Cell Metab,2014,19(2):285-292.
[73] Chan P,Möller A,Liu MC,Sceneay JE,Wong CS,Waddell N,et al.The expression of the ubiquitin ligase SIAH2(seven in absentia homolog 2)is mediated through gene copy number in breast cancer and is associated with a basal-like pheno⁃type and p53 expression[J].Breast Cancer Res,2011,13(1):R19.
[74] Wong CS,Möller A.Siah:a promising anticancer target[J].Cancer Res,2013,73(8):2400-2406.
[75] Bolton JL,Trush MA,Penning TM,Dryhurst G,Monks TJ.Role of quinones in toxicology[J]. Chem Res Toxicol,2000,13(3):135-160.
[76] Whitty A.GrowingPAINS inacademic drug discovery[J].Future Med Chem,2011,3(7):797-801.
[77] Liu J,Xia H,Kim M,Xu L,Li Y,Zhang L,et al. Beclin1 controls the levels of p53 by regulating the deubiquitination activity of USP10 and USP13[J].Cell,2011,147(1):223-234.
[78] Gavory G,O′Dowd C,Mcclelland K,Odrzywol E,Brown A, Burton S,et al.AbstractLB-257:discovery and characterization of novel,highly potent and selective USP7 inhibitors[J].Cancer Res,2015,75(15 Suppl):LB-257
[79] Sowa ME,Bennett EJ,Gygi SP,Harper JW. Defining the human deubiquitinating enzyme interaction landscape[J].Cell,2009,138(2):389-403.
[80] Faesen AC,Dirac AM,Shanmugham A,Ovaa H,Perrakis A,Sixma TK.Mechanism of USP7/ HAUSP activation by its C-terminal ubiquitin-like domain and allosteric regulation by GMP-synthe⁃tase[J].Mol Cell,2011,44(1):147-159.
[81] Morgan MT,Haj-Yahya M,Ringel AE,Bandi P,Brik A,Wolberger C.Structural basis for histone H2B deubiquitination by the SAGA DUB module[J].Science,2016,351(6274):725-728.
[82] Samara NL,Wolberger C.A new chapter in the transcription SAGA[J].Curr Opin Struct Biol,2011,21(6):767-774.
[83] Kee Y,Yang K,Cohn MA,Haas W,Gygi SP,D′andrea AD.WDR20 regulates activity of the USP12 x UAF1 deubiquitinating enzyme complex[J].J Biol Chem,2010,285(15):11252-11257.
[84] Dahlberg CL,Juo P.TheWD40-repeat proteins WDR-20 and WDR-48 bind and activate thedeubiquitinating enzyme USP-46 to promote the abundance of the glutamate receptor GLR-1 in the ventral nerve cord of Caenorhabditis elegans[J].J Biol Chem,2014,289(6):3444-3456.
[85] Chen J,Dexheimer TS,Ai Y,Liang Q,Villamil MA,Inglese J,et al.Selective and cell-active inhibitors ofthe USP1/UAF1 deubiquitinase complex reverse cisplatin resistance in non-small cell lung cancer cells[J].Chem Biol,2011,18(11):1390-1400.
[86] Liang Q,Dexheimer TS,Zhang P,Rosenthal AS,Villamil MA,You C,et al.A selective USP1-UAF1 inhibitorlinks deubiquitination to DNA damage responses[J].Nat Chem Biol,2014,10(4):298-304.
[87] Bolshan Y,Getlik M,Kuznetsova E,Wasney GA,Hajian T,Poda G,et al.Synthesis,optimization,and evaluation of novel small molecules as antag⁃onists of WDR5-MLL interaction[J].ACS Med Chem Lett,2013,4(3):353-357.
[88] Senisterra G,Wu H,Allali-Hassani A,Wasney GA,Barsyte-Lovejoy D,Dombrovski L,et al.Smallmolecule inhibition of MLL activity by disruption of its interaction with WDR5[J].Biochem J,2013,449(1):151-159.
[89] Villamil MA,Liang Q,Zhuang Z.The WD40-repeat protein-containing deubiquitinase complex:catalysis,regulation,and potential for therapeutic intervention[J].Cell Biochem Biophys,2013,67(1):111-126.
[90] Ernst A,Avvakumov G,Tong J,Fan Y,Zhao Y,Alberts P,et al.A strategy for modulation of enzymes in the ubiquitin system[J].Science,2013,339(6119):590-595.
[91] Zhang W,Wu KP,Sartori MA,Kamadurai HB,Ordureau A,Jiang C,et al.System-wide modulation ofHECT E3 ligases with selective ubiquitin variant probes[J].Mol Cell,2016,62(1):121-136.
[92] Wauer T,Swatek KN,Wagstaff JL,Gladkova C,Pruneda JN,Michel MA,et al.Ubiquitin Ser65 phosphorylation affects ubiquitin structure,chain assembly and hydrolysis[J].EMBO J,2015,34(3):307-325.
[93] Chin LS,Li L.Ubiquitin phosphorylation in Parkinson′s disease:Implications for pathogenesis and treatment[J].Transl Neurodegener,2016,5:1.
[94] Matsuda N.Phospho-ubiquitin: upending the PINK-Parkin-ubiquitin cascade[J].J Biochem,2016,159(4):379-385.
开发针对泛素特异性蛋白酶的高质量化学探针
童宇峰1,2
(1.Structural Genomics Consortium,Toronto,Ontario,Canada;2.Department of Pharmaoclogy and Toxicology,University of Toronto,Toronto,Ontario,Canada)
泛素特异性蛋白酶能去除靶蛋白上的泛素标签。泛素特异性蛋白酶失调可以导致多种疾病。虽然工业界和学术界对泛素特异性蛋白酶投入了大量研究,目前针对泛素特异性蛋白酶的高质量化学抑制剂仍然很少。本文提出了泛素特异性蛋白酶化学探针开发需要达到的标准以及策略。
化学探针;药物开发;小分子抑制剂;泛素特异性蛋白酶
童宇峰,Tel:+1(416)946-3876,E-mail:yufeng.tong@utoronto.ca
2016-05-30 接受日期:2016-06-14)
R914
:A
:1000-3002-(2016)06-0611-09
10.3867/j.issn.1000-3002.2016.06.001
Yu-feng TONG,Tel:+1(416) 946-3876,E-mail:yufeng.tong@utoronto.ca
(本文编辑:乔 虹)
猜你喜欢
杂志排行

中国药理学与毒理学杂志的其它文章
- Brief Introduction of the Executive Committee of the Chinese Pharmacological Society North America Chapter(CPSNAC) and the Specialists in This Issue
- Role of calcium-activated potassium channels in neuronal pacemaker activity
- Reversal effects of desipramine on resistance of U251/TR cells to temozolomide
- Effects of calcium channel blockers on growth cone and their clinical implications
- Pathological role of transient receptor potential melastatin member 2 channel in neurodegenerative diseases and Alzheimer disease
- Drug reward memory:implication from drug-induced conditioned place preference model