ω-3鱼油脂肪乳对肝细胞氧化损伤的修复作用及其机制
2016-02-07李智强杨大刚王惠群
李智强,杨大刚,王惠群
(贵州医科大学,贵阳550004)
ω-3鱼油脂肪乳对肝细胞氧化损伤的修复作用及其机制
李智强,杨大刚,王惠群
(贵州医科大学,贵阳550004)
目的 观察ω-3鱼油脂肪乳对肝细胞氧化损伤的修复作用,并探讨其机制。方法 体外培养人肝细胞系HL7702细胞。取对数生长期的HL7702细胞分成对照组、模型组和观察组,每组9孔。对照组仅加培养基常规培养;模型组、观察组在培养基中加入500 μmol /L过氧化氢(H2O2),培养1 h后观察组加入0.5% ω-3鱼油脂肪乳;另设空白对照组,只加培养基,无细胞。各组继续培养4 h后收集细胞,油红O染色观察各组细胞内脂滴变化,CCK-8法测算各组细胞存活率,DCFH-DA荧光探针法检测各组细胞内活性氧簇(ROS),比色法检测各组细胞培养上清液丙氨酸转氨酶(ALT) 、天冬氨酸转氨酶(AST),COD-PAP单试剂比色法检测各组细胞内甘油三酯(TG)、总胆固醇(TC),硫代巴比妥酸法检测各组细胞内丙二醛(MDA) ,黄嘌呤氧化酶法检测各组细胞内超氧化物歧化酶(SOD),实时荧光定量PCR法检测各组细胞中过氧化物酶体增生物激活受体(PPAR)α mRNA,Western blotting法检测各组细胞中肝型脂肪酸结合蛋白(L-FABP)相对表达量。结果 光镜下可见脂滴染为橘红色。对照组组HL7702肝细胞排列紧密, 仅见少许橘红色脂滴。模型组肝细胞可见大量橘红色脂滴并伴有脂滴融合现象。观察组肝细胞可见较多橘红色脂滴, 但与模型组比较显著减少。对照组、模型组、观察组细胞存活率分别为67.25%±4.68%、71.72%±2.47%、100%。观察组细胞存活率高于模型组(P<0.05),但低于对照组(P<0.05)。与对照组比较,模型组细胞内ROS增高(P<0.05),细胞培养上清液ALT、AST增高(P均<0.05),细胞内TG 、TC 、MDA增高(P均<0.05),细胞内SOD降低(P<0.05),PPARα mRNA降低(P<0.05),L-FABP相对表达量降低(P<0.05)。与模型组比较,观察组细胞内ROS降低(P<0.05),细胞培养上清液ALT、AST均降低(P均<0.05),细胞内TG 、TC 、MDA均降低(P均<0.05),细胞内SOD活性增高(P<0.05),PPARα mRNA增高(P<0.05), L-FABP相对表达量增高(P<0.05)。结论 ω-3鱼油脂肪乳可修复人肝HL7702细胞氧化损伤,其机制可能与其上调L-FABP / PPARα信号通路相关因子的表达、维持肝细胞脂质稳态有关。
ω-3鱼油脂肪乳;肝细胞;氧化损伤;细胞存活;过氧化物酶体增生物激活受体α;肝型脂肪酸结合蛋白
人机体内正常情况下活性氧的产生和清除处于动态平衡。当内源性或外源性的刺激破坏平衡后可致活性氧大量生成。当活性氧超过机体抗氧化系统清除能力时,会引起细胞DNA氧化损伤及细胞内蛋白质表达异常,并产生细胞毒效应,最终对机体造成不可逆的损伤[1,2]。肝脏细胞的氧化损伤在很多肝脏疾病发生发展过程中普遍存在,如肝脏的缺血-再灌注损伤、梗阻性黄疸并发肝损伤、脂肪性肝病、药物性肝病、病毒性肝炎、肝纤维化等[3,4]。ω-3鱼油脂肪乳含有二十碳五烯酸(EPA) 和二十二碳六烯酸(DHA)。研究表明EPA和DHA在抗炎、抗血栓、降低血脂、抗过敏等方面具有重要作用[5,6],在人体中也具有抗氧化的功能[7]。但ω-3鱼油脂肪乳的抗氧作用具体机制尚未完全明确。本研究观察ω-3鱼油脂肪乳对人肝HL7702细胞氧化损伤的修复作用,并探讨其可能机制。现报告如下。
1 材料与方法
1.1 细胞及主要试剂 人肝细胞系HL7702细胞购自中国科学院上海细胞生物研究所。RPMI1640培养基购自美国HyClone公司,胎牛血清购自杭州四季青公司。H2O2溶液购自国药公司。ω-3鱼油脂肪乳注射液购自奥地利费森尤斯卡比公司。CCK-8试剂盒购自日本同仁研究所,ROS、TG、TC、MDA、SOD、ALT、AST检则试剂盒均购自南京建成生物技术公司。油红O染液购自Solarbio公司。RIPA裂解液、SDS-PAGE凝胶试剂盒、SDS-PAGE 蛋白上样缓冲液和BCA蛋白浓度测定试剂盒均购自上海碧云天生物技术研究所。PCR引物由上海生工生物公司合成,总RNA提取纯化试剂盒及实时荧光定量PCR试剂盒购自日本TaKaRa公司。兔抗人L-FABP单克隆抗体购自Abcam公司,辣根过氧化物酶标记的羊抗兔二抗和兔抗人β-actin多克隆抗体购自博士德公司。
1.2 HL7702细胞分组、氧化损伤细胞模型建立及ω-3鱼油脂肪乳应用 人肝细胞系HL7702细胞用含10%FBS的RPMI1640培养基在37 ℃、5%CO2的培养箱内培养,取对数生长期的细胞,用0.25%胰蛋白酶消化将细胞浓度调整为5 × 104/mL,接种于6孔板或96孔板,细胞过夜贴壁后换液,分成对照组、模型组和观察组,每组9孔。对照组仅加培养基常规培养;模型组、观察组在培养基中加入500 μmol /L过氧化氢(H2O2),培养1 h后观察组加入0.5% ω-3鱼油脂肪乳。各组细胞在37 ℃、5%CO2的培养箱内继续培养4 h。另设空白对照组,只加培养基,无细胞。
1.3 模型、观察组组细胞存活率测算 采用CCK-8 法。各组细胞继续培养4 h后,每孔中加入10 μL的CCK-8溶液,孵育4 h,用酶标仪在450 nm 波长测定各孔的吸光度(OD值),计算细胞存活率。模型组、观察组细胞存活率(%)=(模型、观察组OD值-空白对照组OD值) /(对照组OD 值-空白对照组OD值)×100%。
1.4 各组细胞内脂滴观察 各组细胞继续培养4 h后弃培养液,PBS清洗2次,4%多聚甲醛固定30 min,加入油红O染色1 h,异丙醇脱色30 s,苏木素复染10 s,显微镜下(400×)观察细胞内的脂滴变化。
1.5 各组细胞内活性氧簇(ROS)检测 各组细胞继续培养4 h后,按照ROS试剂盒说明书用DCFH-DA荧光探针法检测ROS,荧光酶标仪测定荧光强度,激发波长(500±15)nm,发射波长(530±20)nm。结果用荧光强度表示。
1.6 各组细胞培养上清液ALT、AST检测 各组细胞继续培养4 h后,收集培养液1 000 r/min 离心5 min,取上清,按照ALT、AST检测试剂盒说明书检测ALT、AST。
1.7 各组细胞内TG、TC、MDA 、SOD检测 各组细胞继续培养4 h后,弃培养液,PBS清洗2次,用0.25%胰蛋白酶消化后加入完全培养基,1 000 r/min离心5 min后弃上清液,用RIPA裂解液裂解细胞,4 ℃下12 000 r/min离心10 min,取上清,BCA蛋白浓度试剂盒检测蛋白含量,按照TG 、TC、MDA、SOD检测试剂盒说明书上的方法计算1 mg蛋白质所对应的TG 、TC、MDA及SOD。
1.8 各组细胞过氧化物酶体增生物激活受体(PPAR)α mRNA检测 采用实时荧光定量PCR法。各组细胞继续培养4 h后收集细胞,按RNA提取试剂盒说明书方法提取各组细胞总RNA,按照实时荧光定量PCR试剂盒说明书扩增PPARα基因, PCR反应条件为95 ℃预变性3 min,95 ℃10 s,60 ℃ 30 s,40个循环。PPARα引物正向:5′-CAGGCTTCGCAAACTTGGAC-3′,反向:5′-GCTACCAGCATCCCGTCTTT-3′,扩增产物长度为119 bp。以β-actin为内参照,β-actin引物正向:5′-AGTTGCGTTACACCCTTTCTTG-3′,反向:5′-TCACCTTCACCGTTCCAGTTT-3′,扩增产物长度为185 bp。以2-ΔΔCt表示PPARα mRNA相对表达量。
1.9 各组细胞肝型脂肪酸结合蛋白(L-FABP)蛋白检测 采用Western blotting法。各组细胞继续培养4 h后收集细胞提取总蛋白,BCA 法测定蛋白浓度, 蛋白质进行变性处理后按各组细胞的蛋白浓度加样,进行SDS-PAGE电泳,电泳结束后将蛋白转移至PVDF膜,用5%脱脂奶粉室温封闭2 h, 兔抗L-FABP单克隆抗体(1∶500 稀释)4 ℃孵育过夜, TBST洗涤3次,二抗(1∶5 000稀释)室温孵育1 h, TBST洗涤3次,ECL化学发光法曝光显影,以β-actin作为内参照,Quantity One软件分析结果。以目的蛋白与内参照灰度值的比值来表示目的蛋白的相对表达量。

2 结果
2.1 各组细胞内脂滴形态比较 光镜下可见脂滴染为橘红色。对照组HL7702肝细胞排列紧密, 仅见少许橘红色脂滴。模型组肝细胞可见大量橘红色脂滴并伴有脂滴融合现象。观察组肝细胞可见较多橘红色脂滴, 但与模型组比较显著减少。
2.2 各组细胞存活率比较 对照组、模型组、观察组细胞存活率分别为100%、67.25%±4.68%、71.72±2.47%。观察组细胞存活率高于模型组(P<0.05),但低于对照组(P<0.05)。
2.3 各组细胞内ROS及细胞培养上清液ALT、AST比较 各组细胞内ROS及细胞培养上清液ALT、AST见表1。

表1 各组细胞内ROS、细胞上清液ALT和AST比较
注:与对照组比较,*P<0.05;与模型组比较,#P<0.05。
2.4 各组细胞TG、TC、MDA及SOD比较 各组细胞TG、TC、MDA及SOD见表2。
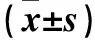
表2 各组细胞TG、TC、MDA及SOD比较
注:与对照组比较,*P< 0.05;与模型组比较,#P< 0.05。
2.5 各组细胞PPARα mRNA相对表达量比较 对照组、模型组、观察组细胞PPARα mRNA相对表达量分别为1、0.63±0.03、0.71±0.05。与对照组比较,模型组细胞PPARα mRNA相对表达量降低(P<0.05);观察组细胞PPARα mRNA相对表达量高于模型组(P<0.05),但低于对照组(P<0.05)。
2.6 各组细胞L-FABP蛋白相对表达量比较 对照组、模型组、观察组组细胞L-FABP蛋白相对表达量分别为0.87±0.03、0.65±0.03、0.76±0.02。与对照组组比较,模型组细胞L-FABP蛋白相对表达量降低(P<0.05);观察组细胞L-FABP蛋白相对表达量高于模型组(P<0.05),但低于对照组(P均<0.05)。
3 讨论
细胞氧化损伤由高浓度氧分子或氧的化学衍生物引起。目前常用的体外细胞氧化损伤模型是用H2O2诱导氧化损伤。H2O2是重要的活性氧,极易透过细胞膜与细胞内铁离子反应形成高活性自由基。本研究通过在培养基中加入500 μmol /L H2O2成功构建肝细胞氧化损伤模型。
ROS是反映细胞氧化损伤程度的直接指标,MDA作为脂质过氧化产物反映了细胞受氧自由基攻击的损伤程度,SOD 是体内最重要的抗氧化酶,SOD水平反映了细胞清除氧自由基的能力。ω-3鱼油脂肪乳在免疫营养治疗中具有维护机体和组织生理功能的作用。Heller等[8]研究发现ω-3鱼油脂肪乳能减少患者住院天数及抗生素的使用量,提高临床疗效和患者的生存率。本研究结果显示,与对照组相比,模型组肝细胞存活率显著降低,细胞内脂滴增多,ROS和MDA水平升高,细胞培养上清液ALT、AST升高,细胞内SOD水平降低。表明模型组肝细胞出现了明显的氧化损伤。在氧化损伤肝细胞培养基中加入ω-3鱼油脂肪乳的观察组,与未加ω-3鱼油脂肪乳的模型组相比,细胞存活率显著增高,细胞内脂滴减少,细胞内ROS及MDA水平降低,细胞培养上清液ALT、AST下降,细胞内SOD升高。表明ω-3鱼油脂肪乳能降低氧化损伤肝细胞内ROS,调节细胞脂质代谢,增加细胞SOD的合成,对肝细胞氧化损伤起到修复作用。其原因是ω-3鱼油脂肪乳的有效成分是ω-3多不饱和脂肪酸和DHA。ω-3多不饱和脂肪酸可调节脂类介质合成和细胞因子释放,从而发挥抗炎和免疫调节的作用。DHA具有很强的清除·OH和O2·的能力,可提高还原型辅酶的活力,抑制磷酸酶的活性,降低细胞内ROS,促进细胞增殖[9]。
PPAR是可被过氧化物酶体增殖物激活的转录因子,包括PPARα、PPARβ 及PPARγ 3种亚型。PPARα主要表达在肝细胞,具有调节脂肪代谢、炎性反应、免疫功能及细胞分化等作用[10]。肝脏内PPARα表达下降可引起细胞能量代谢紊乱、微血管周围炎症、脂肪沉积,引起脂蛋白合成代谢障碍,脂肪酸在肝脏氧化减少,肝脏脂质沉积[11]。李金莲等[12]的研究表明PPARα在H2O2对肝细胞氧化损伤起保护作用。L-FABP是脂肪酸结合蛋白家族重要成员,在肝脏、小肠、肾等组织中均有表达。L-FABP具有调节不饱和脂肪酸、饱和脂肪酸、胆固醇、胆汁酸等转运的功能[13]。L-FABP直接调节肝脏脂肪酸代谢,间接调节脂肪酸在肝细胞内的转运与吸收,维持肝脏脂质稳态[11]。L-FABP 作为内源性抗氧化剂在肝细胞氧化损伤中发挥了重要作用。L-FABP 对多不饱和脂肪酸和脂质过氧化产物有很高的亲和力和结合能力,L-FABP通过结合多不饱和脂肪酸来调节脂肪酸在细胞内氧化途径的利用度,从而控制ROS 的释放量[14]。PPARα可直接与L-FABP发生核信号传导构成L-FABP / PPARα信号通路,形成了正反馈调节[11]。本研究结果显示,模型组细胞PPARα mRNA及L-FABP蛋白相对表达量低于对照组,说明氧化损伤会导致肝细胞内的PPARα mRNA及L-FABP蛋白表达降低。观察组细胞PPARα mRNA及L-FABP蛋白相对表达量高于模型组,说明ω-3鱼油脂肪乳能够上调肝细胞内PPARα mRNA及L-FABP蛋白的表达水平,通过正反馈调节L-FABP / PPARα信号通路改善肝细胞氧化损伤程度。
综上所述,ω-3鱼油脂肪乳可修复人肝细胞系HL7702细胞氧化损伤,机制可能与其上调L-FABP / PPARα信号通路的表达,维持肝细胞脂质稳态有关。
[1] Miyamoto H, Doita M, Nishida K, et al. Effects of cyclic mechanical stress on the production of inflammatory agents by nucleus pulposus and anulus fibrosus derived cells in vitro[J]. Spine, 2006, 31(1):4-9.
[2] 张喜平,黄鑫梅,沈延飞.梗阻性黄疸并发肝损伤机制研究概况[J].医学研究杂志,2008,37(11):8-12.
[3] 吴娜,蔡光明,何群.氧化应激与肝脏损伤[J].世界华人消化杂志,2008,16(29):3310-3315.
[4] Trachootham D, Lu W, Ogasawara MA, et al. Redox regulation of cell survival[J]. Anti Redox Signal, 2008,10(8):1343-1374.
[5] Weintraub HS. Overview of prescription omega-3 fatty acid products for hypertriglyceridemia[J]. Postgrad Med, 2014,126(7):7-18.
[6] Miyata J, Arita M. Role of omega-3 fatty acids and their metabolites in asthma and allergic diseases[J]. Allergol Int, 2015,64(1):27-34.
[7] 来伟旗,张岭,刘臻,等.多不饱和脂肪酸抗氧化功能的人体试验[J]. 职业与健康, 2011, 27(23):2707-2708.
[8] Heller AR, Rossler S, Litz RJ, et al. Omega-3 fatty acids improve the diagnosis-related clinical outcome[J]. Critical Care Med, 2006, 34(4):972-979.
[9] Shimazawa M,Nakajima Y,Mashima G,et al. Docosahexaenoic acid (DHA)has neuroprotective effects against oxidative stress in retinal ganglion cells [J]. Brain Res,2009,1251(28):269-275.
[10] Burri L, Thoresen GH, Berge RK. The role of PPARα activation in liver and muscle.[J]. Ppar Res, 2010,2010(1):1-7.
[11] 张震,富文俊,邢宇锋,等.L-FABP/PPARα信号通路与非酒精性脂肪性肝炎关系的研究进展[J].山东医药,2016,56(8):98-100.
[12] 李金莲,刘军锋,李娜,等.PPAR-α激动剂Wy14643减轻肝细胞系氧化应激损伤[J].基础医学与临床,2012,32(11):1284-1287.
[13] 王南南,徐力致,王亚平.肝型脂肪酸结合蛋白研究进展[J].生命科学,2012(2):3360-3362.
[14] 欧阳冬生,王珍珊,袁浩泳,等.肝脏脂肪酸结合蛋白抗氧化作用研究进展[J].中国现代医学杂志,2011,21(26):3284-3287.
Repair function of ω-3 fish oil fat emulsion on oxidative damage in liver cells and its mechanism
LIZhiqiang,YANGDagang,WANGHuiqun
(GuizhouMedicalUniversity,Guiyang550004,China)
Objective To observe the repair function of ω-3 fish oil fat emulsion on oxidative damage in liver cells and to explore its mechanism. Methods Human hepatic cell line HL7702 was cultured in vitro. HL7702 cells were divided into the control group, model group and observation group, and each group was divided into 9 holes. The control group was cultured with conventional culture medium, in the model and the observation groups, we added 500 μmol/L hydrogen peroxide (H2O2) to the culture medium, after 1 hour, the observation group was added with 0.5% ω-3 fish oil fat emulsion, and then we set a blank control group which was only added with the culture medium without cells. After 4 h of culture, the cells in each group were collected. We observed the intracellular lipid droplets by oil red O staining and calculated the cell survival rate by CCK-8, measured reactive oxygen species (ROS) in the cells of each group by DCFH-DA fluorescence probe, detected the alanine aminotransferase (ALT) and aspartate aminotransferase (AST) in the supernatant of each group by colorimetric assay, the triglyceride (TG) and total cholesterol (TC) by COD-PAP single reagent colorimetric method, malondialdehyde (MDA) by thiobarbituric acid and superoxide dismutase (SOD) by xanthine oxidase, the relative expression of peroxisome proliferator-activated receptor-α (PPAR-α) mRNA by real-time fluorescence quantitative PCR, and the relative expression of liver fatty acid-binding protein (L-FABP) protein by Western blotting. Results Under microscope, the intracellular lipid droplets were orange. The HL7702 liver cells in the control group arrayed closely, and only a few orange red lipid droplets appeared. In the model group, a large number of orange red lipid droplets were observed and accompanied by lipid droplets. In the observation group, the liver cells showed more orange lipid droplets, but they were significantly decreased as compared with those of the model group. The cell survival rates of the control group, the model group and the observation group were 100%, 67.25%±4.68%, and 71.72%±2.47%. The survival rate of the observation group was higher than that of the model group (P<0.05), but was lower than that of the control group (P<0.05). Compared with the control group, the intracellular ROS of the model group was increased (P<0.05), the ALT and AST in cell culture supernatants were increased (allP<0.05), and the intracellular TG, TC, MDA were increased (allP<0.05), the SOD was decreased (P<0.05), the PPARα mRNA was decreased (P<0.05) and the relative expression of L-FABP protein was decreased (P<0.05). Compared with the model group, the intracellular ROS of the observation group was decreased (P<0.05), the ALT and AST in cell culture supernatants were decreased (allP<0.05), the intracellular TG, TC, MDA were decreased (allP<0.05), the SOD was increased (P<0.05), the PPARα mRNA was increased (P<0.05) and the relative expression of L-FABP protein was increased (P<0.05). Conclusions ω-3 fish oil fat emulsion can repair oxidative damage in human hepatic cell line HL7702, and its mechanism may be related to the up-regulation of the expression of L-FABP/ PPARα signaling pathway and the maintenance of lipid homeostasis in liver cells.
ω-3 fish oil fat emulsion; hepatic cells; oxidative damage; cell survival; peroxisome proliferator-activated receptor-α; liver fatty acid-binding protein
贵州省国际科技合作计划项目(20137023)。
李智强( 1984- ) ,男,硕士研究生,主要研究方向为肝脏疾病。E-mail: 969951219@qq.com
简介:杨大刚( 1971- ) ,男,硕士,副教授,主要研究方向为肝胆胰疾病。E-mail: ydg435888@126.com
10.3969/j.issn.1002-266X.2016.47.007
R575
A
1002-266X(2016)47-0025-04
2016-08-15)
