山西霍山植物群落谱系结构的空间格局
2016-02-06姜晓燕梁林峰毕润成
姜晓燕,梁林峰,毕润成,闫 明*
(1 山西师范大学 生命科学学院,山西临汾 041004;2 山西省林业调查规划院,太原 030012)
山西霍山植物群落谱系结构的空间格局
姜晓燕1,梁林峰2,毕润成1,闫 明1*
(1 山西师范大学 生命科学学院,山西临汾 041004;2 山西省林业调查规划院,太原 030012)
以山西霍山七里峪植物群落为研究对象,用R语言分别对9个海拔梯度(1 200~2 000 m)、5个空间尺度(100、400、900、1 600和2 500 m2)和5个空间尺度下划分的6个径级(Ⅰ. DBH<5 cm、Ⅱ. 5 cm≤DBH<10 cm、Ⅲ. 10 cm≤DBH<15 cm、Ⅳ. 15 cm≤DBH<20 cm、Ⅴ. 20 cm≤DBH<25 cm、Ⅵ. DBH≥25 cm)的植物群落谱系结构及其在不同研究方向上的变化规律进行分析,探讨其群落构建成因。结果显示:(1)在所有海拔梯度上,霍山七里峪植物群落都表现出一定的谱系结构,在不同海拔梯度上,群落谱系结构表现出一定的差异性,且在中低海拔群落的谱系结构是发散的,在较高海拔群落的谱系结构是聚集的。(2)5个空间尺度下研究区植物群落都具有谱系结构,且随着空间尺度的增加,谱系聚集程度越来越大。(3)不同径级下的群落都有一定的谱系结构,随着径级的增大,谱系聚集程度逐渐降低,趋向于发散结构,但在Ⅵ径级时群落聚集程度较Ⅳ、Ⅴ径级的高。研究表明,山西霍山七里峪植物群落在不同海拔梯度、不同空间尺度和不同径级下都表现出显著的谱系结构,与中性理论所预测的谱系随机并不相符,证明生态位理论在霍山植物群落构建过程中的作用更加重要。
霍山; 植物群落; 谱系结构; 空间格局
群落构建研究对于解释现有物种共存和物种多样性的维持是至关重要的,因此一直是群落生态学研究中的重点内容[1]。分析群落内物种间的亲缘关系可以反映现有群落形成的历史过程,更加全面的了解进化和生态学进程对群落结构形成的影响[2]。随着分子系统学和谱系群落生态学的迅速发展,群落谱系结构正成为群落生态学研究中的热点内容[3-4]。使用DNA序列进化关系表示物种间的亲缘关系,一方面可以有效衡量群落物种组成,另一方面可以分析群落的谱系结构[5]。谱系结构在进化历史上反映了群落的物种组成[6],通过对其充分的研究可以有效推测群落构建的主要生态过程,进而从全新的角度了解群落构建的成因[7-8]。
谱系群落生态学中的一个基本内容就是群落的谱系结构[9]。谱系结构分为3种基本形式:谱系聚集、谱系发散和谱系随机[3]。物种的中性理论和生态位理论是解释群落现有谱系结构的2个主要理论[10]。根据中性理论,由于随机的生态漂变,所有个体是没有差别的,群落是随机构建的,因而无谱系结构[11]。根据生态位理论,生境过滤作用由于相似的生境特点将会筛选适应能力相似,亲缘关系近的物种共存,会产生谱系聚集的群落。而竞争排斥作用是生态位相似的物种会因为竞争相同的资源而互相排斥产生谱系发散的群落[12]。因此,关于群落谱系结构的研究,有助于探讨中性理论和生态位理论在群落构建中的相对重要性。
在热带森林研究中,群落谱系结构在群落构建方面得到了广泛的应用[6]。Kembel等[4]研究巴拿马样地植物群落谱系结构后,发现随着空间尺度的增大(100 m2及更大的空间尺度),谱系结构会逐渐趋向于聚集。Silva等[13]的研究发现小空间抑制了亲缘关系近的物种共存,呈现出显著的谱系发散。Letcher[14]研究了不同演替阶段下植物群落谱系结构时发现随着演替的深入,谱系结构更趋于发散,并且谱系发散更趋于大径级的群落。黄建雄等[15]在研究谱系结构、环境因子及空间因子对群落动态变化的影响时发现,整体上空间因子是影响群落动态的主要因素。不同研究方向下的群落谱系结构不同,反映出群落构建机制存在一定的差异。鉴于上述情况,本研究对不同海拔梯度、不同空间尺度和不同径级下植物群落谱系结构的变化进行比较分析,以探讨在植物群落构建过程中,生态位理论和中性理论的相对重要性。
1 材料和方法
1.1 研究区域概况
七里峪林场境内植物资源丰富,种类较繁多,植被地带性分布较为明显。境内乔木层的物种有辽东栎(Quercuswutaishanica)、油松(Pinustabuliformis)、白桦(Betulaplatyphylla)等,灌木层的有榛子(Corylusheterophylla)、土庄绣线菊(Spiraeapubescens)、黄刺玫(Rosahugonis)等,草本层以羊胡子草(Eriophorumvaginatum)、野艾蒿(Artemisialavandulaefolia)、唐松草(Thalictrumaquilegifolium)等为主[20]。
1.2 取样方法
在霍山七里峪林场进行踏查,根据人为破坏、生境条件、群落组成等因素的不同选择较好的地段作为样地。采用系统取样的方法,从1 200~2 000 m,海拔每升高100 m为一个梯度,在每个梯度上选择林相良好的典型地段设置样地,在每个样地做4~6个大小为10 m×10 m的乔木样方,在每个乔木样方内分别做2个5 m×5 m的灌木样方和4个1 m×1 m的草本样方,共调查乔木样方48个,灌木样方96个,草本样方192个。另外,在中海拔地段另选一块样地,利用网格法,设置10 m×10 m的样方25个,20 m×20 m的样方16个,30 m×30 m的样方9个,40 m×40 m的样方4个,50 m×50 m的样方2个;调查样地中所有胸径1 cm的个体,记录其树名、株数、胸径、树高、冠幅等。同时记录各样地的基本情况包括海拔、坡向和坡度等[21]。
1.3 谱系树的构建
将所有样地中包含的所有187个物种及其科属信息输入植物谱系库软件Phylomatic[22]中,软件以被子植物分类系统(APG)为基础数据,自动输出由输入物种所构建的谱系树。使用软件Phylocom[23]提供的算法BLADJ,利用分子及化石定年数据[24],计算出谱系树中的每一个分化节点发生的时间。该算法首先将已知分化时间的节点固定,对所有未知节点采用平均的方法处理。这种方式获得的分枝长度代表连续2次物种分化间隔的时间(以百万年为单位),这比简单地将分枝长度设置为1包含更多的可靠数据,因而结果将更加可靠。由于样地中的所有物种隶属61科146属,来源相对广泛,所构建的谱系树足以区分群落之间的谱系结构[25]。
1.4 谱系指数的选择
选择近年来广泛被使用的净谱系亲缘关系指数(net relatedness index,NRI)[3]来代表群落谱系结构,和另一种谱系指数净最近种间亲缘关系指数(net nearest taxa index,NTI)[3]相比,NRI更加侧重从整体上反映研究群落的谱系结构[26]。因此本研究仅以NRI作分析。假定样地中调查到的所有物种组成一个局域物种库,该指数首先计算出样方中所有物种对的平均谱系距离MPD(mean phylogenetic distance),保持物种数量与物种个体数不变,将小样方中物种的种名从物种库中随机抽取999次,从而获得该样方中物种在随机零模型下的MPD的分布,最后利用随机分布结果将观察值标准化,从而获得NRI。其计算公式为[23]:
进而,从同一角度也可很好地理解这样一个建议:“立足专业成长,关注基本问题.”[24]因为,作为一线教师,就只有采取这一立场,面对多变的指导思想才不会无所适从,并切实避免这样一个现象,即是忙了一辈子却不知道忙了什么,乃至一再地重复别人的错误,却没有任何真正的进步.
式中,MPDsample代表观察值,MPDrandsample代表物种在谱系树上通过随机后获得的平均值,SD为标准偏差。若NRI>0,说明小样方的物种在谱系结构上聚集;若NRI<0,说明小样方的物种在谱系结构上发散;若NRI=0,说明小样方的物种在谱系结构上随机。
1.5 分析方法
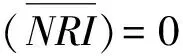
2 结果与分析
2.1 不同海拔梯度下群落谱系结构
沿海拔梯度所设置的48个样方中,共有21个(43.75%)样方的净谱系亲缘关系指数(NRI)大于0,表明样方所在的小局域群落谱系结构是聚集的,有27个(56.25%)样方的净谱系亲缘关系指数(NRI)小于0,则表明样方所在的小局域群落谱系结构是发散的。由表1可知,不同海拔梯度的NRI均值与0均具有显著性差异,均表现出一定的谱系结构。海拔1 700 m~2 000 m的NRI均值极显著大于0,表明植物群落谱系结构聚集,在其余海拔下的NRI均值极显著小于0,说明植物群落谱系结构是发散的。同样由图1可知,谱系结构发散的样方主要集中在中低海拔(1 200~1 600 m)处,而谱系结构聚集的样方主要是在较高海拔(1 700~2 000 m)的地区。总体上,不同海拔梯度下群落的净谱系亲缘关系指数呈现出先下降后上升的趋势(图1)。在海拔1 500 m处,谱系发散指数达到了最大值,竞争排斥作用最为激烈,在海拔2 000 m处,聚集指数达到了最大值,说明生境过滤对群落的影响达到了最大。
2.2 不同空间尺度下的群落谱系结构

度的增大,聚集程度是逐渐增加的。通过方差比较分析可知,10 m×10 m尺度下的群落NRI与其他尺度相比有显著性差异,其他4个尺度下的群落谱系聚集程度较大,而且它们之间无显著差异(图2)。
2.3 不同径级的群落谱系结构
分别计算6个径级群落总体NRI发现,群落谱系结构整体上都表现为谱系聚集,但是随着径级的增大,群落谱系结构越来越趋向于发散,径级增加到Ⅴ径级(20 cm≤DBH<25 cm)时,谱系指数值达到最低,谱系聚集程度最低,但是当DBH≥25 cm时,群落的谱系结构指数值比Ⅳ、Ⅴ径级的大,谱系聚集程度相对较高(图3)。并且在不同空间尺度下观察不同径级的群落谱系结构时发现,各个空间尺度下各径级群落的谱系结构均表现为谱系聚集,而且随着径级的增大,谱系指数值逐渐变小,群落的谱系结构越来越趋向于发散结构,当DBH≥25cm时,相比群落谱系指数值较前2个径级的大,也就是聚集程度比Ⅳ径级、Ⅴ径级的大,这与6径级群落总体的NRI值的变化趋势相符。10 m×10 m空间尺度下的各径级的NRI值均小于其他空间尺度下相对应径级的NRI值,其余尺度下各径级的NRI值是随着空间尺度的增加而增大的(图4)。
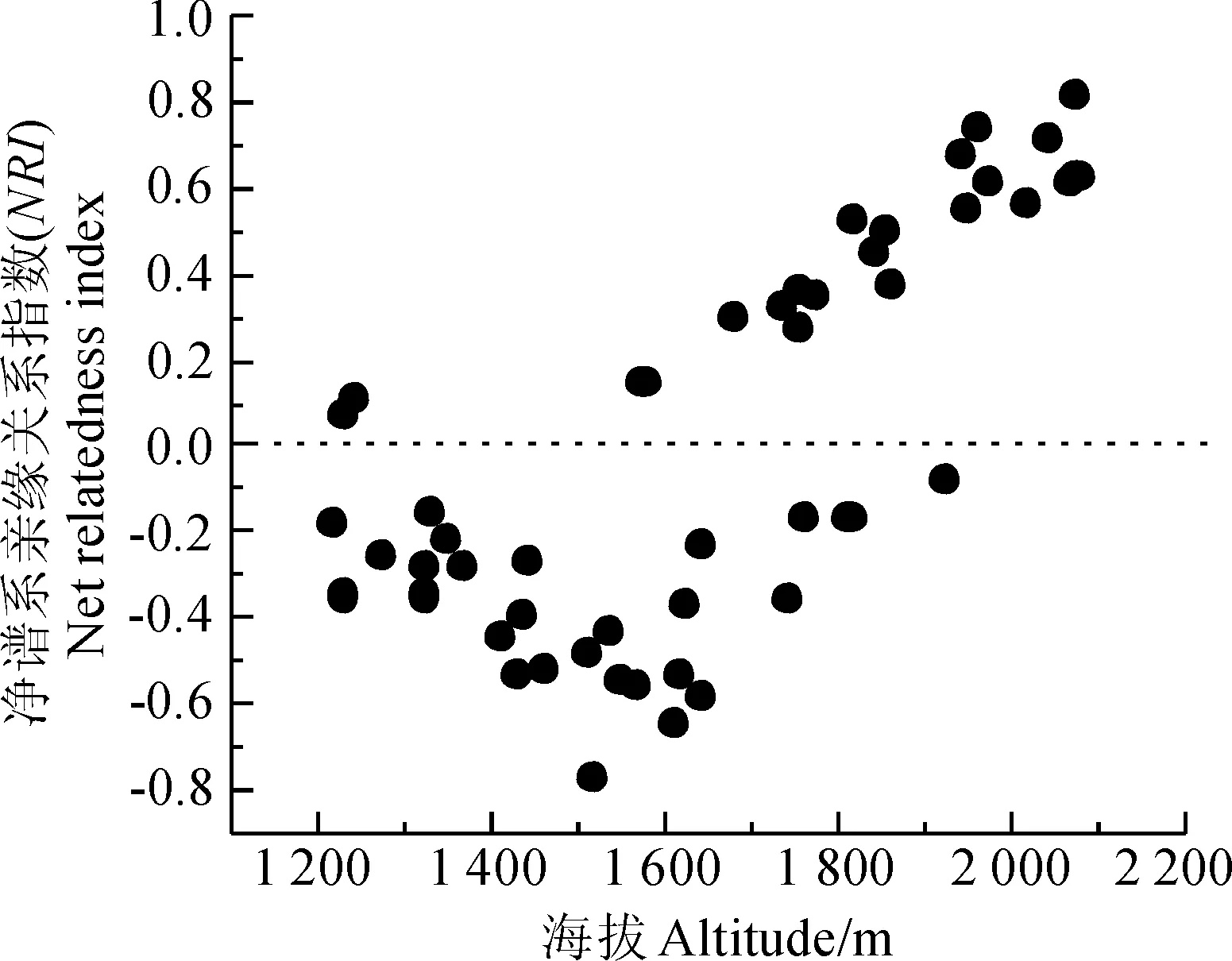
图1 净谱系亲缘关系指数(NRI)与海拔梯度散点图Fig.1 Plot of NRI versus altitude

海拔梯度Altitudegradient/m平均值MeanofNRI标准差StandarddeviationtP1200-0.1147200.203927-1.2580.01∗∗1300-0.2486200.754288-7.3700.01∗∗1400-0.4230600.111646-8.4730.01∗∗1500-0.4327670.310139-3.4180.01∗∗1600-0.4319330.282169-3.7930.01∗∗17000.2497170.2674292.2870.01∗∗18000.3551000.2692072.9500.01∗∗19000.5085800.3344133.4010.01∗∗20000.6766800.10093114.9910.001∗∗∗
Note:**P<0.01,***P<0.001
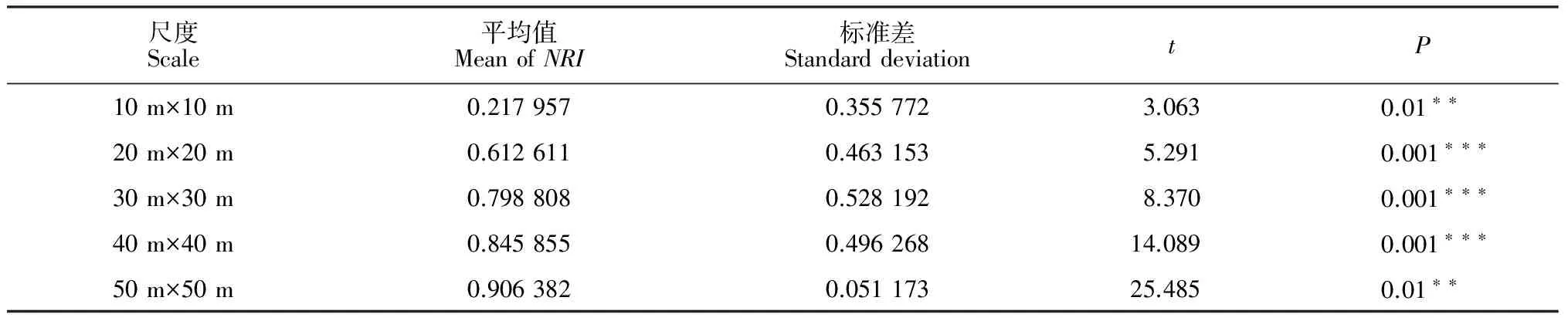
表2 5个空间尺度上群落NRI分布及平均值为0的t检验
Note:**P<0.01,***P<0.001
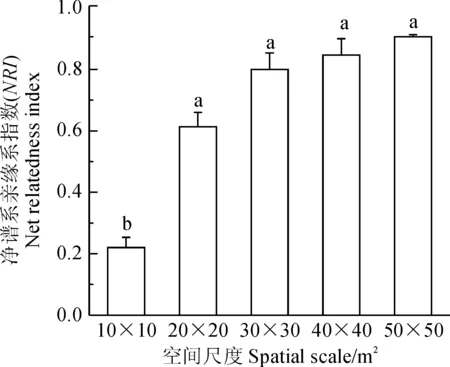
不同小写字母表示0.05水平差异显著图2 不同空间尺度下群落NRI的平均值The normal letters mean significant difference at 0.05 levelFig.2 Mean of NRI of communities at different spatial scales

Ⅰ.DBH<5 cm;Ⅱ. 5 cm≤DBH<10 cm;Ⅲ.10 cm≤DBH<15 cm;Ⅳ.15 cm≤DBH<20 cm;Ⅴ. 20 cm≤DBH<25 cm;Ⅵ. DBH≥25 cm; 下同. The same as below图3 不同径级群落总体上的NRIFig.3 Overall NRI of communities at different diameter class

图4 不同空间尺度下不同径级的群落NRI分布Fig.4 Distributions of NRI of different diameter class at different spatial scales
3 讨 论
一般来说,谱系距离可以用来表示物种亲缘关系,谱系距离小的物种亲缘关系近,生态特征就有可能越相似,对类似环境的适应能力就越一致,也就是说生态位越相似[26]。如果在一个样方中所观察到的物种间的亲缘关系与从物种库中随机抽取的相同物种形成的种间关系显著相近,说明样方内的物种是谱系聚集的,相反,如果观察到的物种亲缘关系与随机物种组成显著地更加疏远,说明样方内物种的谱系结构是发散的,如果观察值与随机值没有显著性差异,说明样方内的物种无谱系结构。生境过滤和竞争排斥是生态位理论的两种主要机制,用来解释形成群落现有谱系结构的成因。生境过滤机制指亲缘关系相近的物种有相似的生境偏好,因此生态特征相似的物种更有机会共存,谱系结构表现为谱系聚集,也就是生境能够筛选亲缘关系相近的物种共存;而竞争排斥机制是指生态特征相似的物种由于竞争相同的资源与环境而会互相排斥,因此会使亲缘关系较远的物种共存,谱系结构表现为谱系发散[3]。随着研究的深入,人们发现不同海拔梯度、空间尺度或者谱系尺度下群落的谱系结构并不同,反映出群落构建成因会由于尺度不同而不同[27]。
海拔梯度是影响群落谱系结构的一个重要因素,研究表明,随着海拔的升高,群落谱系结构由发散逐渐转变为聚集。中低海拔处群落的谱系结构是发散的,表明竞争排斥机制影响着现有群落的谱系结构,中低海拔地段的样方所处位置地势较平缓,区域温度和降水量配置适宜,环境和资源条件优越,气候环境条件较好,生态胁迫作用不明显[21],主要是物种间的竞争作用较大,导致群落内物种间亲缘关系较远的物种可以共存;而在较高海拔处,群落谱系结构是聚集的,表明生境过滤占主导作用,因为高海拔地段环境条件严峻,生境异质性高[28],生理胁迫压力较大,强烈的生境压力驱使具有相同生物学特性即亲缘关系较近的物种聚集在一起形成谱系聚集的结构。从中低海拔谱系发散格局到高海拔聚集格局的转变表现了环境压力逐渐加强,竞争作用逐渐减弱的趋势。这与刘巍等[2]在研究长白山植物群落谱系结构和Kembel等[4]在研究巴拿马大样地内植物群落谱系结构的研究结论类似。
对不同空间尺度下的群落谱系结构的研究发现,在所设置的5个空间尺度下,群落谱系结构都表现为谱系聚集,并且随着空间尺度的增大,谱系结构的聚集程度也增大。谱系结构随着空间尺度变化的原因可能是因为在小的空间尺度下,物种的生境比较均质,各种资源相对有限,导致群落中物种之间的竞争排斥作用占主导,密度制约是竞争排斥的一种作用机制,而且在小尺度上密度制约是维持生物多样性的一个非常重要的作用因子[29-30],随着所研究空间尺度的增大,密度制约作用逐渐减弱,环境变量逐渐增多,空间异质性越来越大,生境过滤成为影响群落谱系结构的主导因素,从而使较大尺度上的群落谱系结构越来越聚集[5]。本研究结论与黄健雄等[6]在研究古田山常绿阔叶林不同尺度下植物群落谱系结构和Swenson等[26]关于5个样地不同空间尺度下植物群落谱系结构的研究结论相一致。
植物群落的谱系结构与树种的径级水平有一定的相关性,树种径级大小是依据空间代时间的方法进行划分的,6个径级整体上的谱系结构都是聚集的,随着径级的增大,聚集程度越来越低,但是当DBH≥25 cm时,NRI值相对前2个径级的大,也就是聚集程度比前2个径级的大,而且在研究5个不同空间尺度下不同径级群落的谱系结构时发现,在每一个空间尺度下,随着径级的增大,群落谱系结构的聚集程度逐渐减少,这与6个径级整体上的变化趋势是相似的,也就是说随着径级的增大,群落越来越趋向于谱系发散结构,这可能是由于大树种子扩散受到限制,导致小树聚集生长,整体上表现为谱系聚集,随着物种径级的增加,同种物种间的竞争排斥作用加强,存活的物种间的地理距离会越来越大,整体表现为谱系发散[5]。这与Swenson等[26]的研究结果相一致,即小径级的谱系聚集或随机,而大径级的谱系结构则越来越趋向于发散的格局。但是本研究树木径级在Ⅵ径级时,群落NRI比Ⅳ、Ⅴ径级的大,聚集程度也相对较高,可能是因为DBH≥25 cm的物种在整个群落中比较单一。这与张奎汉等[31]的研究结果有较高相似。
在目前已有的群落谱系结构研究中,尽管出现了各种研究结果,如谱系聚集、谱系发散、谱系随机,但是在大多数的植物群落内部,物种间的亲缘关系往往呈现出的是非随机的格局[32]。在本次所调查的植物群落中,不同海拔梯度,不同空间尺度和不同径级下的群落类型均表现出一定的谱系结构,而与中性理论所预测的物种的分布趋向于随机,群落构建是随机的,呈现谱系随机的结构[3]并不一致,更加表明生态位理论相对于中性理论在暖温带落叶阔叶林植物群落构建过程中的重要作用。
研究群落的谱系结构对探讨群落构建成因、群落发展动态有着重要的作用。通过研究群落的谱系结构来探讨某种机制对群落构建的主导作用已经被广泛应用。关于谱系指数NRI现在也被很多研究所采用,一方面是由于其设计科学[6],另一方面,计算该指数的R软件中的picante程序包以及Phylocom软件的发展也为该指数的应用提供了有力的支持。由于谱系结构受到不同尺度、环境因子和生态特征等多方面的影响,而现在关于谱系结构的研究还主要是用生境过滤、竞争排斥和中性理论来解释,因此以后通过谱系结构的研究判断群落构建时,需要全面考虑这几方面的因素[33]。作为群落谱系生态学中的一项重要内容,谱系结构的研究会随着分子生物学的发展和一些相关软件的完善在生态学中得到广泛的应用。本研究应用NRI指数对七里峪植物群落的谱系结构进行了分析,探讨了其在不同研究方向上的变化规律,并分析了生态位理论和中性理论在植物群落构建过程中的相对重要性,但是由于谱系结构的研究开展起来的时间并不久,在很多方面需要进一步的完善,如果想要更加深入探讨群落构建成因,还需结合生态特征、环境变量、谱系结构和多种生态过程进行共同分析。
[1] ROSINDELL J, HUBBELL S P, ETIENNE R S. The unified neutral theory of biodiversity and biogeography at age ten[J].TrendsinEcology&Evolution, 2011,26(7): 340-348.
[2] 刘 巍, 曹 伟. 长白山植物群落谱系结构及环境因子对其的影响[J]. 干旱区资源与环境, 2013,27(5): 63-68.
LIU W, CAO W. Phylogenetic structure and influence of environmental factors on phylogenetic structure of plant community in Changbai Mountains[J].JournalofAridLandResourcesandEnvironment, 2013,27(5): 63-68.
[3] WEBB C O, ACKERLY D D, MCPEEK M A,etal. Phylogenies and community ecology[J].AnnualReviewofEcologySystematics, 2002,33(1): 475-505.
[4] KEMBEL S W, HUBBELL S P. The phylogenetic structure of a neotropical forest tree community[J].Ecology, 2006,87(S7): S86-S89.
[5] 牛红玉, 王峥峰, 练琚愉, 等. 群落构建研究的新进展: 进化和生态相结合的群落谱系结构研究[J]. 生物多样性, 2011,19(3): 275-283.
NIU Y H, WANG Z F, LIAN J Y,etal. New progress in community assembly : community phylogenetic structure combining evolution and ecology[J].BiodiversityScience, 2011,19(3): 275-283.
[6] 黄建雄, 郑凤英, 米湘成. 不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响[J]. 植物生态学报, 2010,34(3): 309-315.
HUANG J X, ZHENG F Y, MI X C. Influence of environmental factors on phylogenetic structure at multiple spatial scales in an evergreen broad-leaved forest of China[J].ChineseJournalofPlantEcology, 2010,34(3): 309-315.
[7] MERWIN L, HE T, LAMONT B B. Phylogenetic and phenotypic structure among Banksia communities in south-western Australia[J].JournalofBiogeography, 2012,39(2): 397-407.
[8] 冷海楠, 崔福星, 张荣涛, 等. 兴安落叶松林植物群落谱系结构研究[J]. 黑龙江科学, 2014,5(12): 10-12.
LENG H N, CUI F X, ZHANG R T,etal. Research of forest community phylogeny based on phylomatic fromLarixgmeliniiforest[J].HeilongjiangScience, 2014,5(12): 10-12.
[9] 宋 凯, 米湘成, 贾 琪, 等. 不同程度人为干扰对古田山森林群落谱系结构的影响[J]. 生物多样性, 2011,19(2): 190-196.
SONG K, MI X C, JIA Q,etal. Variation in phylogenetic structure of forest communities along a human disturbance gradient in Gutianshan forest, China[J].BiodiversityScience, 2011,19(2): 190-196.
[10] 柴永福, 岳 明. 植物群落构建机制研究进展[J]. 生态学报, 2016,36(15): 1-8.
CAI Y F, YUE M. Research advances in plant community assembly mechanisms[J].ActaEcologicaSinica, 2016,36(15): 1-8.
[11] GETSCHKE G. The unified neutral theory of biodiversity and biogeography[J].PlantSystematics&Evolution, 2002,85(11): 3 172-3 174.
[12] WIENS J J, GRAHAM C H. Niche conservatism: integrating evolution, ecology and conservation biology[J].AnnualReviewofEcology,EvolutionandSystematics, 2005,36(1): 519-539.
[13] SILVA I A, BATALHA M A. Phylogenetic overdispersion of plant species in southern Brazilian savannas[J].BrazilianJournalofBiology, 2009,69(3): 845-851.
[14] LETCHER S G. Phylogenetic structure of angiosperm communities during tropical forest succession[J].ProceedingsoftheRoyalSocietyB:BiologicalSciences, 2010,277(1 678): 97-104.
[15] 黄建雄, 叶万辉, 练琚愉, 等. 谱系结构、环境因子及空间因子对群落动态变化的影响[J]. 科学通报, 2014,59(35): 3 471-3 478.
HUANG J X, YE W H, LIAN J Y,etal. Detecting the influence of phylogenetic structure, environmental factors and PCNM factors in population dynamics in a subtropical forest community in Guangdong, China[J].Chin.Sci.Bull., 2014,59(35): 3 471-3 478.
[16] 茹文明, 张金屯, 毕润成, 等. 山西霍山森林群落林下物种多样性研究[J]. 生态学杂志, 2005,24(10): 1 139-1 142.
RU W M, ZHANG J T, BI R C,etal. Species diversity of undergrowths in Huoshan Mountain of Shanxi Province[J].ChineseJournalofEcology, 2005,24(10): 1 139-1 142.
[17] 白 聪, 闫 明, 毕润成, 等. 山西太岳山兴唐寺红柄白鹃梅群落优势种的空间格局分析[J]. 植物生态学报, 2014,38(12): 1 283-1 295.
BAI C, YAN M, BI R C,etal. Spatial pattern analysis of dominant species inExochordagiraldiicommunity in Xingtang Temple of Taiyue Mountains, Shanxi, China[J].ChineseJournalofPlantEcology, 2014,38(12): 1 283-1 295.
[18] 吴征镒. 中国植被[M]. 北京: 科学出版社, 1980: 279-280.
[19] 闫 明, 毕润成. 山西霍山植被分类及不同演替阶段群落物种多样性的比较分析[J]. 植物资源与环境学报, 2009,18(3): 56-62.
YAN M, BI R C. Vegetation classification and comparative analysis of species diversity of community at different succession stages in Huoshan Mountain of Shanxi Province[J].JournalofPlantResourcesandEnvironment, 2009,18(3), 56-62.
[20] 田慧霞, 闫 明. 山西七里峪油松种群空间格局的分形特征[J]. 生态学杂志, 2015,34(2): 326-332.
TIAN H X, YAN M. Fractal properties of spatial pattern ofPinustabuliformispopulation in Qiliyu, Shanxi[J].ChineseJournalofEcology, 2015,34(2): 326-332.
[21] 何艳华, 闫 明, 张钦弟, 等. 五鹿山国家级自然保护区物种多样性海拔格局[J]. 生态学报, 2013,33(8): 2 452-2 462.
HE Y H, YAN M, ZHANG Q D,etal. Altitudinal pattern of plant species diversity in the Wulu Mountain Nature Reserve, Shanxi, China[J].ActaEcologicalSinica, 2013,33(8): 2 452-2 462.
[22] WEBB C O, DONOGHUE M J. Phylomatic: tree assembly for applied phylogenetics[J].Mol.Ecol.Notes, 2005,5(1): 181-183.
[23] WEBB C O, ACKERLY D D, KEMBEL S. Phylocom: software for the analysis of phylogeneitic community structure and character evolution (with phylomatic and ecovolve) - user’s manual Version4.1[EB/OL]. http://phylodiversity. net/phylocom/, 2009.
[24] WIKSTROM N, SAVOLAINEN V, CHASE M W. Evolution of the angiosperms: calibrating the family tree[J].ProceedingsoftheRoyalSocietyB:BiologicalSciences, 2001,268(1 482): 2 211-2 220.
[25] WEBB C O, CANNON C H, DAVIES S J. Ecological organization, biogeography, and the phylogenetic structure of tropical forest tree communities[M]//Carson W P, Schnizer S A eds. Tropical Forest Community Ecology. Oxford: Wiley Blackwell, 2008.
[26] SWENSON N G, ENQUIST B J, THOMPSON J,etal. The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities[J].Ecology, 2007,88(7): 1 770-1 780.
[27] PRINZING A, DURKA W, KLOTZ S,etal. The niche of higher plants: evidence for phylogenetic conservatism[J].ProceedingsoftheRoyalSocietyB:BiologicalSciences, 2001,268(1 483): 2 383-2 389.
[28] LAI J S, MI X C, REN H B,etal. Species-habitat associations change in a subtropical forest of China[J].JournalofVegetationScience, 2009,20(3): 415-423.
[29] WEBB C O, GILBERT G S, DONOGHUE M J. Phylodiversity-dependent seeding mortality, size structure, and disease in a bornean rain forest[J].Ecology, 2006,87(7 Suppl): S123-S131.
[30] WRIGHT J S. Plant diversity in tropical forests: a review of mechanisms of species coexistence[J].Oecologia, 2002,130(1): 1-14.
[31] 张奎汉, 鲍大川, 郭屹立, 等. 后河自然保护区珍稀植物群落谱系结构的时空变化[J]. 植物科学学报, 2013,31(5): 454-460.
ZHANG K H, BAO D C, GUO Y L,etal. Phylogenetic structure of a rare plant community in Houhe Nature Reserve over Time and Space[J].PlantScienceJournal, 2013,31(5), 454-460.
[32] VAMOSI S M, HEARD S B, VAMOSI J C,etal. Emerging patterns in the comparative analysis of phylogenetic community structure[J].MolecularEcology, 2009,18(4): 572-592.
[33] PAUSAS J G, VERDU M. The jungle of methods for evaluating phenotypic and phylogenetic structure of communities[J].Bioscience, 2010,60(8): 614-625.
(编辑:潘新社)
Spatial Pattern of Phylogenetic Structure of Plant Community in Shanxi Huoshan Mountain
JIANG Xiaoyan1, LIANG Linfeng2, BI Runcheng1, YAN Ming1*
(1 College of Life Sciences, Shanxi Normal University, Linfen, Shanxi 041004, China; 2 Shanxi Forest Inventory and Planning Institute, Taiyuan 030012, China)
Qiliyu region is located between the eastern margin of the Lingkong mountain and northern margin of the Mianshan Mountain, and it shows distinct transitional vegetation types and high species richness. The phylogenetic structure of a community is composed of the genetic relationships between species across the community. Phylogenetic structure is a synthetical indicator reflecting community construction’s three ecological processes which are habitat filtering, competitive exclusion and stochastic processes. Understanding of the phylogenetic structure of a community will provide new insights into the different ecological processes in constructing the community. Our objective was to examine the effect of spatial factor on phylogenetic structure and determine the relative importance of neutral theory and niche theory in building the plant community in Huoshan mountain. We compared the phylogenetic structure of the community along different altitude gradient using data from 1 200 to 2 000 m, and examined the effects of five spatial scales(100, 400, 900, 1 600 and 2 500 m2) and six DBH classes(Ⅰ. DBH<5 cm,Ⅱ. 5 cm≤DBH<10 cm,Ⅲ. 10 cm≤DBH<15 cm,Ⅳ. 15 cm≤DBH<20 cm,Ⅴ. 20 cm≤DBH<25 cm,Ⅵ. DBH≥25 cm) at five spatial scales on phylogenetic structure of the community in Qiliyu region. The results demonstrated that: (1) there were certain phylogenetic structure at all different altitudinal gradient. The phylogenetic structure of community was dispersed at middle-low altitude and clustered at higher altitude. (2) Communities were phylogenetically clustered at all spatial scales. With increasing spatial scale, the phylogenetic structure tended to cluster. (3) The effects of six DBH classes at five spatial scales on phylogenetic structure of the community were examined, which shows the result that the degree of phylogenetic clustering decreased with increasing DBH classes. However, the degree of phylogenetic clustering at the scale of DBH Ⅵ was higher than the Ⅳ and Ⅴ. It turns out from our research that the prediction of niche theory is more important than the prediction of neutral theory in building the plant community in Huoshan Mountain of Shanxi.
Huoshan mountain; plant community; phylogenetic structure; spatial pattern
1000-4025(2016)12-2505-08
10.7606/j.issn.1000-4025.2016.12.2505
2016-09-10;修改稿收到日期:2016-11-02
国家自然科学基金(31270461);山西师范大学校基金(ZR1211)
姜晓燕(1994-),女,在读硕士研究生,主要从事群落生态学研究。E-mail:794746952@qq.com
*通信作者:闫 明,副教授,硕士生导师,主要从事植物生态学研究。E-mail: mycorrhiza@sina.com
Q948.15+7
A
