乙型肝炎患者外周血CD4 + T 细胞Runx3 的表达及意义
2015-12-31臧国庆王洁玲汤正好余永胜
张 毅,臧国庆,王洁玲,汤正好,江 红,奚 敏,余永胜
上海交通大学附属第六人民医院感染病科,上海200233
乙型肝炎病毒(hepatitis B virus,HBV)感染是一个严重的公共卫生问题。据统计,全球每年约有100万人死于HBV 感染所致的肝硬化、肝衰竭和原发性肝癌[1]。我国现有慢性HBV 感染患者约9 300 万人,其中慢性乙型肝炎(chronic hepatitis B,CHB)患者约2 000万人[2]。研究发现CHB 患者体内Th 细胞分化出现偏移,存在Th1、Th2 细胞及其分泌的细胞因子比例失衡[3]。Th1 型免疫应答低下使机体不能产生足量的细胞毒性T 淋巴细胞(cytotoxic T lymphocytes,CTL)和Th1 型细胞因子以杀伤、破坏HBV 感染的靶细胞,HBV 复制不能得到抑制;而Th2 型细胞因子又可抑制Th1 细胞的免疫应答,使机体不能及时清除HBV,最终导致HBV 慢性持续感染[3]。Th1/Th2 失衡是CHB 发生、发展和不同转归的重要原因[4-6]。Runx3(runtrelated transcription factor 3)是一种肿瘤抑制基因,在机体正常生长、发育和肿瘤发生、发展过程中起着重要作用[7-8]。研究发现Runx3 是CD4+T 细胞分化过程中重要的转录因子,可以调节Th 细胞的分化,从而影响Th1/Th2 的平衡[9-11]。但目前尚无Runx3 与乙型肝炎的相关研究。本文旨在探讨乙型肝炎患者外周血CD4+T 细胞Runx3 的表达及意义。
1 资料与方法
1.1 一般资料 选取2011 年6 月-2012 年12 月上海交通大学附属第六人民医院感染病科诊治的CHB患者37 例(CHB 组),男23 例,女14 例,年龄21 ~58岁,平均年龄(38.1 ±16.5)岁,诊断符合2010 年《慢性乙型肝炎防治指南》[12],其中轻度20 例、中度11例、重度6 例。急性乙型肝炎(acute hepatitis B,AHB)患者11 例(AHB 组),男7 例,女4 例,年龄20 ~55岁,平均年龄(36.9 ±15.7)岁,诊断符合2000 年《病毒性肝炎防治方案》[13]。所有患者均排除其他肝炎病毒、人免疫缺陷病毒、EB 病毒、巨细胞病毒等病原体感染,排除自身免疫性疾病、肿瘤等疾病。所有患者就诊前6 个月内均未使用过干扰素、核苷(酸)类似物等抗病毒药物和(或)胸腺肽等免疫调节药物。健康对照者19 名(健康对照组),男12 名,女7 名,年龄23 ~56岁,平均年龄(37.8 ±14.6)岁,排除各系统疾病,6 个月内未使用过免疫调节药物。CHB 组、AHB 组、健康对照组之间性别、年龄相比,差异无统计学意义(P >0.05),具有可比性。
1.2 方法
1.2.1 主要试剂:RosetteSep 富集人CD4+T 细胞抗体(StemCell 公司),人淋巴细胞分离液(StemCell 公司),总RNA 提取试剂盒(Invitrogen 公司),反转录试剂盒(Promega 公司),荧光定量PCR 试剂盒(Takara 公司)。
1.2.2 外周血CD4+T 细胞分离:无菌条件下采集CHB 患者、AHB 患者和健康对照者肘静脉血各10 ml置于肝素抗凝管中;加入500 μl RosetteSep 富集人CD4+T 细胞抗体混合物,充分混匀,室温孵育20 min;用等体积含2% FBS 的PBS 稀释血液并缓慢混匀;将稀释血液按稀释血液:淋巴细胞分离液=2∶1 的比例在10 ml 淋巴细胞分离液的上面缓慢加入稀释的血液样品,室温1 200 r/min 离心20 min;收集位于淋巴细胞分离液与血浆界面之间的灰色细胞层;加入5 倍体积含2%的PBS 混匀,1 200 r/min 离心10 min,弃上清,再重复洗涤富集细胞1 次,分离得到CD4+T 细胞;流式细胞仪检测CD4+T 细胞纯度达90%以上,-80 ℃保存备用。
1.2.3 引物设计与合成:根据NCBI GenBank 公布的人Runx3 基因编码区序列,应用Primer Premier 5.0 软件设计引物,以GAPDH 基因为内参照。引物由上海生工生物工程技术服务有限公司合成。引物序列如下:Runx3 上游引物5'-ACTGTGATGGCAGGCAATG-3';Runx3 下 游 引 物5'-GTGATGGTCAGGGTGAAACTC-3';GAPDH 上游引物5'-GCACCGTCAAGGCTGAGAAC-3';GAPDH 下游引物5'-GCCTTCTCCATGGTGGTGAA-3'。
1.2. 4 总RNA 提取与反转录合成cDNA:按照Invitrogen 公司的总RNA 提取试剂盒说明书进行操作提取总RNA;按照Promega 公司的反转录试剂盒说明书进行操作反转录合成cDNA,反应条件(20 μl 反应体系):42 ℃孵育60 min,70 ℃灭活10 min。 -80 ℃保存备用。
1.2.5 实时定量PCR 检测Runx3 mRNA 表达水平:在ABI 7500 PCR 仪上进行实时定量PCR 检测,按照Takara 公司的荧光定量PCR 试剂盒说明书进行操作,每份cDNA 做3 个重复管。PCR 反应体系:SYBR Premix Ex Taq 12.5 μl,上下游引物各0.5 μl,cDNA 模板1. 0 μl,ddH2O 10. 5 μl。PCR 反应条件:预变性95 ℃5 min,变性95 ℃15 s,退火60 ℃1 min,共40个循环。PCR 循环结束后对扩增曲线进行分析,3 个重复管取平均Ct 值计算,并对所扩增的PCR 产物熔解曲线进行分析。应用2-△△Ct法计算Runx3 mRNA的相对表达水平。
1.3 统计学处理 采用SPSS 16.0 统计软件进行分析,计量资料用±s 表示,组间比较采用LSD-t 检验,P <0.05 为差异有统计学意义。
2 结果
2.1 各组外周血CD4+T 细胞Runx3 mRNA 表达水平 CHB 组Runx3 mRNA 的表达水平明显低于AHB组和健康对照组(P <0.01),AHB 组Runx3 mRNA 的表达水平明显高于健康对照组(P <0.05,见表1)。
表1 各组外周血CD4 +T 细胞Runx3 mRNA 表达水平(±s)Tab 1 Runx3 mRNA expression in peripheral CD4 +T cells of each group (±s)
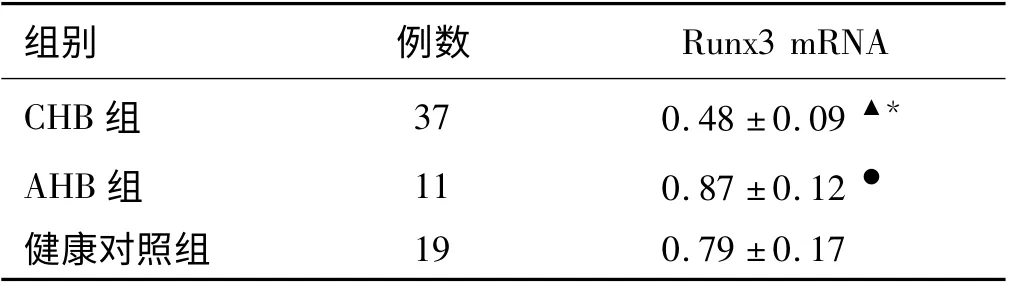
表1 各组外周血CD4 +T 细胞Runx3 mRNA 表达水平(±s)Tab 1 Runx3 mRNA expression in peripheral CD4 +T cells of each group (±s)
注:与健康对照组比较,▲P <0.01,●P <0.05;与AHB 组比较,* P <0.01。
组别 例数Runx3 mRNA CHB 组 37 0.48 ±0.09▲*AHB 组 11 0.87 ±0.12 ●健康对照组19 0.79 ±0.17
2.2 CHB 患者轻、中、重度组外周血CD4+T 细胞Runx3 mRNA 表达水平 CHB 重度组Runx3 mRNA的表达水平明显高于CHB 轻度组(P <0.05);CHB 轻度组与CHB 中度组、CHB 中度组与CHB 重度组Runx3 mRNA 的表达水平相比,差异无统计学意义(P >0.05,见表2)。
表2 CHB 患者外周血Runx3 mRNA 表达水平(±s)Tab 2 Runx3 mRNA expression in CHB patients (±s)

表2 CHB 患者外周血Runx3 mRNA 表达水平(±s)Tab 2 Runx3 mRNA expression in CHB patients (±s)
注:与轻度组比较,* P <0.05。
组别 例数Runx3 mRNA轻度组20 0.39 ±0.18中度组 11 0.55 ±0.23重度组 6 0.65 ±0.27*
3 讨论
乙型肝炎自然感染率高,80% ~90% 的儿童和5% ~10%的成年人感染HBV 后会转变为慢性HBV携带者,成为慢性肝炎、肝硬化和肝细胞癌发病的高危人群[14-15]。CHB 发病机制非常复杂,目前尚未完全阐明。
在HBV 特异性抗原的刺激下,初始CD4+T 细胞在不同细胞因子的作用下分化为Th1、Th2 等细胞亚群。Th1、Th2 细胞的分化状态及其相互之间的平衡在机体抗HBV 的免疫应答中起着重要作用[4-6]。Kasahara 等[16]研究发现Th1 细胞依靠其分泌的多种细胞因子发挥直接抗HBV 或间接免疫调节作用,同时其分泌的细胞因子是机体产生HBV 特异性CTL 的必要条件。Penna 等[17]研究发现Th1 型细胞因子可造成肝细胞的损伤,但其在AHB 的恢复过程中发挥了重要作用。Maruyama 等[18]研究发现Th2 细胞及其细胞因子与HBV 持续感染有关。当Th1 细胞亚群占优势时,将促进细胞免疫反应,增强CD8+T 细胞的活性,从而清除肝细胞内的病毒,但同时增加了肝脏的炎症反应而加重肝细胞的损伤;当Th2 细胞亚群占优势时,将促进体液免疫反应,并抑制细胞免疫反应,CTL 活性减弱,肝细胞损伤减轻[19]。
Runx3 基因于1994 年由Levanon 等[7,20]研究发现,曾先后被命名为AML2、PEBP2αC 和CBFA3。人Runx3 基因位于染色体1p36.1,基因全长约67 kb,含有2 个启动子、6 个外显子和1 290 bp 的开放阅读框[20-21]。Runx3 是调节Th 细胞分化重要的转录因子,其可以通过增强T-bet(T-box expressed in T cells)或减弱GATA3(GATA binding protein 3)的转录活性,促使CD4+T 细胞向Th1 细胞分化[9-11]。Djuretic 等[10]研究发现Runx3 在Th1 细胞中以T-bet 依赖的方式被诱导,Th1 细胞中IFN-γ 的大量产生和IL4 基因的沉默需要T-bet 和Runx3 的共同作用;在Runx3 缺失的Th2细胞中,T-bet 不能抑制IL4,但T-bet 和Runx3 共同作用却能显著抑制IL4。Kohu 等[11]研究发现Runx3 在Th2 细胞中也能被诱导,它可以与GATA3 相互作用并削弱GATA3 的转录活性,Runx3 过表达能促使Th 细胞向Th1 细胞分化,并下调Th2 细胞的免疫应答。
本研究显示,CHB 组Runx3 mRNA 的表达水平明显低于AHB 组和健康对照组,而AHB 组Runx3 mRNA 的表达水平明显高于健康对照组。CHB 患者Runx3 低水平表达不能有效促进Th1 细胞免疫应答,无法清除体内病毒,从而使机体处于HBV 持续感染状态。Brooks 等[22]研 究 发 现CHB 患 者 体 内 存 在Th1/Th2失衡,Th1 细胞反应不足,Th1 型免疫应答低下而Th2 型免疫应答亢进,Th1 细胞减少是乙型肝炎保护性抗体不能产生的重要原因。Maini 等[23]研究发现急性自限性HBV 感染主要是细胞免疫反应的结果,细胞因子多为Th1 型,而慢性HBV 感染细胞免疫反应很弱或测不出,体液免疫却相对较强,细胞因子多为Th2 型。
本研究还显示,CHB 重度组Runx3 mRNA 的表达水平明显高于CHB 轻度组。Jiang 等[24]研究发现慢性HBV 感染者Th1 细胞随肝脏炎症活动的加剧而明显增多,而Th2 细胞比例在慢性HBV 感染的不同阶段则较为恒定,但均明显高于健康对照组。Löhr 等[25]研究发现急性重症乙型肝炎患者Th1 细胞的增殖反应比急性非重症乙型肝炎患者强,类似结果在慢性重症乙型肝炎患者中也得到证实。
综上所述,Runx3 在CHB 的发病过程中可能发挥了重要作用,CHB 患者Runx3 表达水平降低可能与Th1/Th2 失衡有关,但能否通过上调Runx3 的表达水平促使CHB 患者外周血Th 细胞向Th1 细胞分化和Th1 型细胞因子的分泌从而促进HBV 的清除还有待进一步研究。
[1] Ganem D,Prince AM. Hepatitis B virus infection-natural history and clinical consequences [J]. N Engl J Med,2004,350 (11):1118-1129.
[2] Lu FM,Zhuang H. Management of hepatitis B in China[J]. Chin Med J (Engl),2009,122(1):3-4.
[3] Marinos G,Rossol S,Carucci P,et al. Immunopathogenesis of hepatitis B virus recurrence after liver transplantation[J]. Transplantation,2000,69(4):559-568.
[4] Jiang YF,Ma ZH,Zhao PW,et al. Effect of thymosin-α1 on T-helper 1 cell and T-helper 2 cell cytokine synthesis in patients with hepatitis B virus e antigen-positive chronic hepatitis B[J]. J Int Med Res,2010,38(6):2053-2062.
[5] Jiang Y,Ma Z,Xin G,et al. Th1 and Th2 immune response in chronic hepatitis B patients during a long-term treatment with adefovir dipivoxil[J]. Mediators Inflamm,2010,2010:143026.
[6] Zheng Y,Huang Z,Chen X,et al. Effects of telbivudine treatment on the circulating CD4+T-cell subpopulations in chronic hepatitis B patients[J]. Mediators Inflamm,2012,2012:789859.
[7] Cohen MM Jr. Perspectives on RUNX genes:an update[J]. Am J Med Genet A,2009,149A(12):2629-2646.
[8] Bangsow C,Rubins N,Glusman G,et al. The RUNX3 gene-sequence,structure and regulated expression[J]. Gene,2001,279(2):221-232.
[9] Wong WF,Kohu K,Chiba T,et al. Interplay of transcription factors in T-cell differentiation and function:the role of Runx[J]. Immunology,2011,132(2):157-164.
[10] Djuretic IM,Levanon D,Negreanu V,et al. Transcription factors Tbet and Runx3 cooperate to activate Ifng and silence Il4 in T helper type 1 cells[J]. Nat Immunol,2007,8(2):145-153.
[11] Kohu K,Ohmori H,Wong WF,et al. The Runx3 transcription factor augments Th1 and down-modulates Th2 phenotypes by interacting with and attenuating GATA3 [J]. J Immunol,2009,183 (12):7817-7824.
[12] Chinese Society of Hepatology and Chinese Society of Infectious Diseases,Chinese Medical Association. The guideline of prevention and treatment for chronic hepatitis B (2010 Version)[J]. Zhonghua Gan Zang Bing Za Zhi,2011,19(1):13-24.中华医学会肝病学分会,中华医学会感染病学分会. 慢性乙型肝炎防治指南(2010 年版)[J]. 中华肝脏病杂志,2011,19(1):13-24.
[13] Chinese Society of Infections Diseases and Parasitology and Chinese Society of Hepatology,Chinese Medical Association. The program of prevention and treatment for viral hepatitis[J]. Zhonghua Gan Zang Bing Za Zhi,2000,8(6):324-329.中华医学会传染病与寄生虫病学分会,中华医学会肝病学分会.病毒性肝炎防治方案[J]. 中华肝脏病杂志,2000,8(6):324-329.
[14] Chisari FV,Ferrari C. Hepatitis B virus immunopathogenesis[J].Annu Rev Immunol,1995,13:29-60.
[15] Te HS,Jensen DM. Epidemiology of hepatitis B and C viruses:a global overview[J]. Clin Liver Dis,2010,14(1):1-21.
[16] Kasahara S,Ando K,Saito K,et al. Lack of tumor necrosis factor alpha induces impaired proliferation of hepatitis B virus-specific cytotoxic T lymphocytes[J]. J Virol,2003,77(4):2469-2476.
[17] Penna A,Del Prete G,Cavalli A,et al. Predominant T-helper 1 cytokine profile of hepatitis B virus nucleocapsid-specific T cells in acute self-limited hepatitis B[J]. Hepatology,1997,25(4):1022-1027.
[18] Maruyama T,McLachlan A,Iino S,et al. The serology of chronic hepatitis B infection revisited [J]. J Clin Invest,1993,91(6):2586-2595.
[19] You J,Zhuang L,Ma YL,et al. Research advances in the imbalance of helper T lymphocyte subpopulations and cytokine network in patients with chronic hepatitis B[J]. World Chinese Journal of Digestology,2007,15(8):791-799.游晶,庄林,马永良,等. 慢性乙型肝炎的Th 细胞亚群及相关细胞因子网络失衡[J]. 世界华人消化杂志,2007,15(8):791-799.
[20] Puig-Kröger A,Corbí A. RUNX3:a new player in myeloid gene expression and immune response[J]. J Cell Biochem,2006,98(4):744-756.
[21] Bae SC,Takahashi E,Zhang YW,et al. Cloning,mapping and expression of PEBP2 alpha C,a third gene encoding the mammalian Runt domain[J]. Gene,1995,159(2):245-248.
[22] Brooks DG,Teyton L,Oldstone MB,et al. Intrinsic functional dysregulation of CD4 T cells occurs rapidly following persistent viral infection[J]. J Virol,2005,79(16):10514-10527.
[23] Maini MK,Boni C,Lee CK,et al. The role of virus-specific CD8+cells in liver damage and viral control during persistent hepatitis B virus infection[J]. J Exp Med,2000,191(8):1269-1280.
[24] Jiang R,Feng X,Guo Y,et al. T helper cells in patients with chronic hepatitis B virus infection[J]. Chin Med J(Engl),2002,115(3):422-424.
[25] Löhr HF,Krug S,Herr W,et al. Quantitative and functional analysis of core-specific T-helper cell and CTL activities in acute and chronic hepatitis B[J]. Liver,1998,18(6):405-413.
