MMP-2与TIMP-2在豚鼠形觉剥夺性近视眼脉络膜表达的变化及调控机制
2015-12-29吴振凯,刘双珍,夏晓波等
MMP-2与TIMP-2在豚鼠形觉剥夺性近视眼脉络膜表达的变化及调控机制
吴振凯1刘双珍夏晓波毛俊峰许惠卓
(中南大学湘雅医院眼科,湖南长沙410000)
摘要〔〕目的观察豚鼠形觉剥夺性近视眼脉络膜组织中基质金属蛋白酶(MMP)-2及其特异性组织抑制剂(TIMP-2)表达的变化及其调控机制。方法将42只3周龄的三色豚鼠随机分为A、B、C组,A组为对照组,共12只,将其随机分为A1组和A2组,A1组6只,饲养14 d后处死,A2组6只,饲养21 d后处死;B组为遮盖组,共18只,随机分为B1、B2、B3组,B1组6只,将半透明眼罩缝于右眼遮盖14 d后处死,B2组6只,将半透明眼罩缝于右眼遮盖21 d后处死,B3组6只,将半透明眼罩缝于右眼遮盖14 d后去除遮盖7 d后处死;C组为注药组,随机分为C1、C2组,C1组6只,予右眼球周注射20 μl浓度为10-6 mmol/L的全反式视黄酸(RA)溶液,C2组6只,予右眼球周注射全反式视黄酸的助溶剂二甲基亚砜(DMSO)溶液作为对照,C1组、C2组均饲养21 d后处死;各组豚鼠左眼不做干预,A1组、B1组、B2组、B3组在饲养14 d后检测眼球屈光度及眼轴长度,A2组、B2组、B3组、C1组、C2组在饲养21 d后检测眼球屈光度及眼轴长度。豚鼠在处死后摘除眼球,对脉络膜组织进行MMP-2和TIMP-2特异性组织抑制剂免疫组化染色,采用免疫组化吸光度测量软件对脉络膜组织阳性着色区进行吸光度测定。结果B1组右眼在遮盖14 d后眼轴延长,近视形成,脉络膜组织中MMP-2表达增强,与自身左眼、对照组A1组右眼相比差异均具有统计学意义(P<0.05)。B1组右眼脉络膜组织中TIMP-2染色阳性区灰度值与对照组相比差异无统计学意义(P>0.05);B2组右眼在遮盖21 d后眼轴延长,近视形成,脉络膜组织中MMP-2表达增强,与自身左眼、对照组A2组右眼、B1组右眼相比,差异均具有统计学意义(P<0.05)。B2组脉络膜组织中TIMP-2染色阳性区灰度值与对照组相比差异无统计学意义(P>0.05);B3组右眼在遮盖14 d后眼轴延长,近视形成,与自身左眼、A1组右眼相比,差异均具有统计学意义(P<0.05),与B2组右眼相比,差异不具有统计学意义(P>0.05);去除遮盖7 d后,近视屈光度较B3组右眼在遮盖14 d时降低,眼轴较B3组右眼在遮盖14 d时缩短(均P<0.05)。脉络膜组织中MMP-2与自身左眼、对照组A2组右眼、B1组右眼相比,差异均具有统计学意义(P<0.05)。B3组右眼脉络膜TIMP-2染色阳性区灰度值与对照组相比差异无统计学意义(P>0.05);C1组右眼球周注射全反式视黄酸溶液21 d后,眼轴延长、近视形成,脉络膜MMP-2染色阳性区灰度值与A2组、C2组右眼、自身左眼相比,差异均有统计学意义(P<0.05),C1组脉络膜组织中TIMP-2染色阳性区灰度值差异无统计学意义(P>0.05)。结论剥夺豚鼠的形觉可增强脉络膜中MMP-2表达并造成豚鼠近视,在豚鼠眼球旁一次性注射全反式视黄酸溶液可导致脉络膜中MMP-2表达增强并造成豚鼠近视,脉络膜组织中的视黄酸可能是调控基质金属蛋白酶-2 的表达最终导致近视的上游调控因子。
关键词〔〕MMP-2;TIMP-2;RA;近视
中图分类号〔〕R7〔文献标识码〕A〔
基金项目:湖南省自然科学基金资助项目(08JJ3047)
通讯作者:刘双珍(1951-),女,主任医师,教授,博士生导师,主要从事斜视、弱视、屈光不正、肿瘤整形的研究。
1常德市第一人民医院眼科
第一作者:吴振凯(1982-),男,硕士,主治医师,主要从事白内障、眼底病、屈光不正及斜弱视的研究。现工作于常德市第一人民医院。
随着对近视发病机制研究的深入,脉络膜在调控眼球生长中的作用日益受到重视。有研究发现视觉信息的变化可引起脉络膜厚度的代偿性改变〔1〕,但其具体机制尚未阐明。脉络膜组织富含胶原成分,而基质金属蛋白酶(MMP)-2能降解Ⅳ、Ⅴ、Ⅸ、Ⅺ型胶原,但尚未明确脉络膜中MMP-2在近视发生发展过程中的变化及其调控机制,本研究拟观察形觉剥夺性近视眼脉络膜中MMP-2及其特异性组织抑制剂(TIMP-2)表达的变化,并球周注射全反式视黄酸(RA)溶液探讨其调控机制。
1材料与方法
1.1分组与干预将42只3周龄的豚鼠(动物均购于中南大学动物部动物研究所)随机分A 、B、C组,A组12只,作为对照组,将其随机分为A1组和A2两组,其中A1组6只,饲养14 d后拉颈处死,A2组6只,饲养21 d后拉颈处死;B组18只,为遮盖组,将其随机等分为B1、B2、B3三组,其中B1组6只,将自制的半透明眼罩缝于右眼的眶周对右眼进行遮盖,饲养14 d后拉颈处死,B2组6只,将半透明眼罩缝于右眼遮盖21 d后拉颈处死,B3组6只,将半透明眼罩缝于右眼遮盖14 d后再去除遮盖的眼罩,再饲养7 d后拉颈处死;C组:注药组,分为C1、C2两组,每组6只,在实验开始第1天予C1组豚鼠的右眼球周注射20 μl浓度为10-6mmol/L的RA溶液,C2组球周注射20 μl RA的助溶剂二甲基亚砜(DMSO)溶液,均为单次注射,21 d后处死,豚鼠左眼为自身对照眼。
1.2屈光度与眼轴的检测屈光度:0.15%托吡卡胺眼液点眼3次将瞳孔充分散大,带状检影验光检查豚鼠屈光度数。眼轴长度:以1%丁卡因滴眼液滴眼3次进行表面麻醉,表面麻醉成功后,以A超测定豚鼠的眼轴长度。屈光度及眼轴长度均为检查3次后取平均值。
1.3组织切片行脉络膜脱黑色素HE染色按预定时间处死豚鼠后剖取眼球,沿赤道部环形剪开眼球后将所取眼球后极部组织用4%多聚甲醛固定24 h,梯度酒精脱水,制作成石蜡切片,切片常规脱蜡至水,0.25%高锰酸钾处理1~2 min使组织呈均匀的棕黄色时进行水洗,以2%的草酸溶液漂白组织2 min后在显微镜下进行检查以确定黑色素是否去除,将黑色素被完全去除的组织切片进行充分水洗,PBS冲洗;3%H2O2处理10 min,以去除内源性过氧化物酶的活性,行HE染色,于光镜下观察脉络膜微结构。
1.4免疫组化染色兔抗人多克隆抗体MMP-2、兔抗人多克隆抗体TIMP-2、兔二步法检测试剂盒、DAB显色试剂盒均购自北京中杉金桥公司,石蜡切片进行脱蜡处理,水化后脱黑色素处理,以3%H2O2溶液阻断内源性的过氧化物酶,对抗原进行热修复,将切片置于温度为95℃~98℃、pH值为6.0的枸橼酸盐溶液中处理20 min,在室温下进行自然冷却;一抗(兔抗人多克隆抗体MMP-2、兔抗人多克隆抗体TIMP-2,PBS 1∶100稀释)4℃过夜;二抗(PowerVision免疫组化检测试剂)37 ℃孵育,在显微镜下控制DAB的显色,以苏木素进行复染,脱水后透明处理,以中性树胶封切片处理。在光镜下,随机选取脉络膜中央断面区域,采用免疫组化吸光度测量软件对脉络膜组织阳性着色区进行吸光度测定。
1.5统计学分析采用SPSS18.0软件进行t检验和单因素方差分析。
2结果
由表1,2可见,B1组右眼眼轴长度、近视屈光度数以及脉络膜MMP-2染色阳性区灰度值与自身左眼、A1组右眼相比,差异均具有统计学意义(P<0.05)。脉络膜TIMP-2染色阳性区灰度值与自身左眼、A1组右眼相比,差异无统计学意义(P>0.05);B2组右眼长度、近视屈光度数以及脉络膜MMP-2染色阳性区灰度值与自身左眼、A2组右眼、B1组右眼相比,差异均具有统计学意义(P<0.05)。脉络膜TIMP-2染色阳性区灰度值与自身左眼、A2组右眼、B1组右眼相比,差异无统计学意义(P>0.05);B3组右眼眼轴长度以及近视屈光度数与自身左眼、A1组右眼相比,差异均具有统计学意义(P<0.05),与B2组右眼相比,差异不具有统计学意义(P>0.05);去除遮盖7 d后,近视度为(-0.15±0.46)D,眼轴长度为(8.16±0.17)mm,近视屈光度较B3组右眼在遮盖14 d时降低,眼轴缩短(P<0.05)。脉络膜MMP-2染色阳性区灰度值与自身左眼、A2组右眼、B1组右眼相比,差异均具有统计学意义(P<0.05)。脉络膜组织TIMP-2染色阳性区灰度值与自身左眼、A2组右眼、B1组右眼相比,差异无统计学意义(P>0.05)。C1组右眼球周注射RA溶液21 d后,眼轴延长、近视形成,近视度,眼轴长度,脉络膜MMP-2染色阳性区灰度值,与A2组、C2组右眼、自身左眼相比,差异均有统计学意义(P<0.05)。脉络膜组织TIMP-2染色
阳性区灰度值与A2组、C2组右眼、自身左眼相比,差异无统计学意义(P>0.05)。HE染色后随机双盲阅片发现经过形觉剥夺或者球周注射RA溶液后实验眼后极部脉络膜较对照眼相同部位的脉络膜萎缩变薄,见图1。
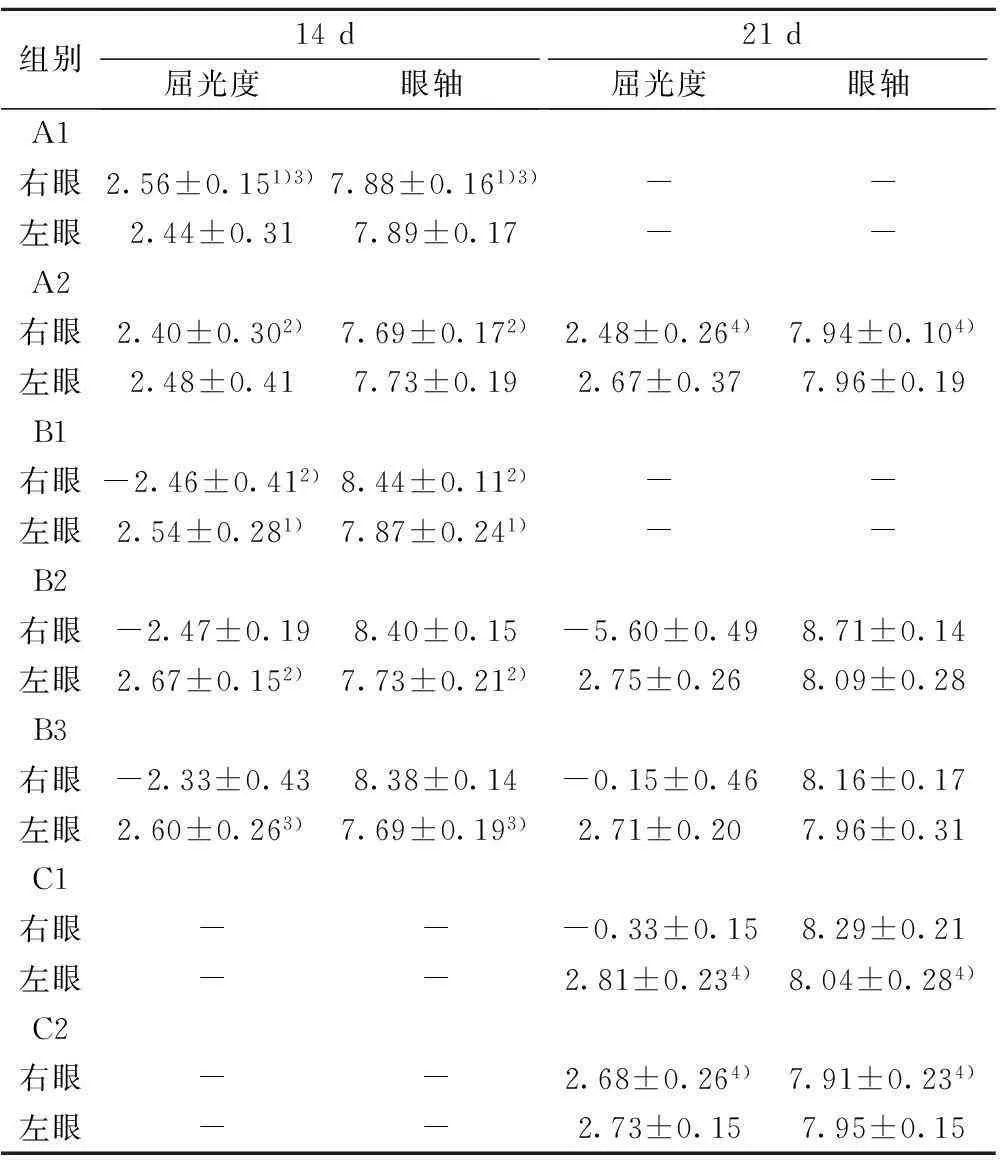
表1 各组屈光度及眼轴长度的比较( n=6,mm)
与B1组比较:1)P<0.05;与B2组比较:2)P<0.05;与B3组比较:3)P<0.05;与C1组比较:4)P<0.05;下表同
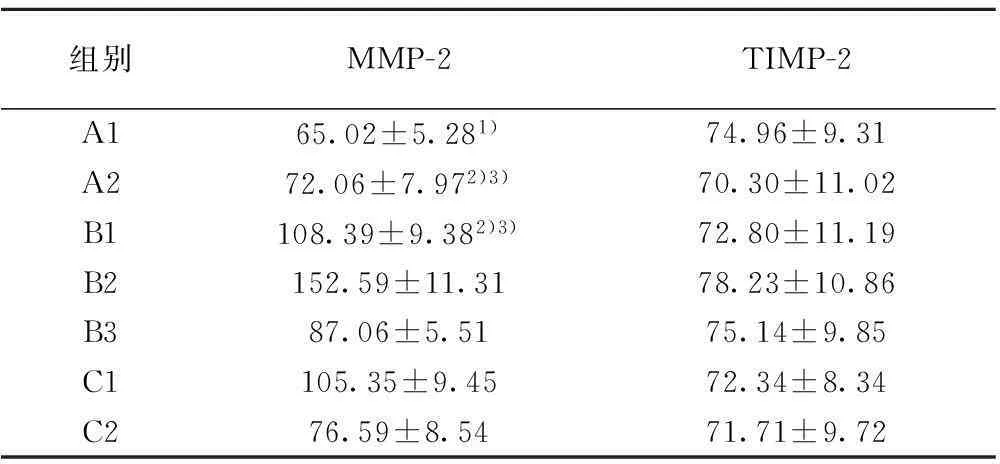
表2 各组脉络膜MMP-2及TIMP-2染色阳性区
3讨论
MMP-2为一种明胶酶A,通常以酶原的形式存在于组织中,在脉络膜的黑色素细胞〔2〕、视网膜色素上皮细胞〔3〕等细胞内均有表达。在特定的情况下MMP-2可被激活而具有降解Ⅳ、Ⅴ、Ⅸ、Ⅺ型胶原的作用。TIMP-2是MMP-2的特异性抑制因子,可以以非共价键形式紧密结合MMP-2并与之形成复合物,可抑制MMP-2降解细胞外基质的作用。MMP-2和TIMP-2之间的平衡决定了MMP-2的活性,当两者之间的平衡关系被打破时,细胞外基质成分的降解和重组之间的动态平衡也会随之而被打破。
目前关于近视眼脉络膜厚度调控机制尚未被阐明,据实验报道〔4〕,剥夺形觉可以导致近视的发生和发展,在近视进展时,眼球后极部脉络膜的厚度变薄,而当形觉剥夺被去除后,脉络膜厚度随之部分恢复,这说明外界的视觉信息的变化可通过某些目前未知的途径导致脉络膜厚度的变化,而脉络膜厚度的变化最终导致了眼轴长度的变化。本研究发现正常豚鼠脉络膜中有MMP-2和TIMP-2的少量表达,在剥夺豚鼠的形觉2 w后,豚鼠眼脉络膜组织中MMP-2表达的明显增强,且其表达随着形觉剥夺的时间延长而增加。在形觉剥夺被去除一段时间后,脉络膜组织中的MMP-2表达较去形觉剥夺之前明显减弱,而在此期间MMP-2的表达却始终未见明显变化,即脉络膜中MMP-2和TIMP-2之间的平衡在近视产生的过程中被打破,而在形觉剥夺被去除后,近视程度在一定程度上减轻,在此过程中MMP-2和TIMP-2之间的平衡又趋向于恢复。豚鼠眼球脉络膜组织中MMP-2的表达在剥夺形觉后增强、去剥夺后减弱,这与形觉剥夺性近视眼中脉络膜厚度的变化方向是一致的。由于玻璃膜的内胶原层、弹力层、外胶原层含有丰富的胶原纤维组织,且脉络膜上腔主要由起源于巩膜、脉络膜、基质层中疏松的胶原纤维组成框架,因此豚鼠形觉剥夺性近视眼脉络膜组织中基质金属蛋白酶-2表达的增强极可能增加脉络膜组织细胞外基质成分的降解,导致脉络膜厚度的变化,我们进行的组织切片显微镜下观察到的豚鼠形觉剥夺眼脉络膜的萎缩变薄证实了这种推论,这也符合临床上所观察到的人类近视眼脉络膜萎缩变薄的现象。
据研究报道,豚鼠形觉剥夺后,随着近视的发生和眼轴的延长,脉络膜及视网膜组织中视黄酸的含量也随之增加,而在去形觉剥夺后,随着近视程度的减轻和眼轴长度的缩短,脉络膜及视网膜RA含量也随之而下降〔5〕,这种脉络膜及视网膜组织中视黄酸的含量随近视程度变化而变化的趋势与我们研究所发现的形觉剥夺与去形觉剥夺时脉络膜中MMP-2变化的趋势高度一致。有研究报道〔6〕,视黄酸可以通过调控生长因子如TGF-β等而间接调控MMP家族的表达,而且我们前期的研究〔7〕也证实了外源性RA可增强后极部巩膜MMP-2的表达,据此,我们推测形觉剥夺性近视眼后极部组织中RA水平的变化可能会影响脉络膜组织中MMP-2与TIMP-2之间的平衡。眼组织具有强有力的吸聚视黄酸的能力,Mertz等〔8〕以24 mg/kg 视黄酸喂养小鸡时,鸡的视网膜组织中视黄酸的浓度达到普通内源性视黄酸浓度水平的2倍,而在脉络膜组织中则达到惊人的70倍。另外,有研究发现仅仅只给予极少量的外源性视黄酸,即可造成动物眼视网膜组织中视黄酸含量的升高,同时可观察到动物眼轴的延长〔9〕,据此我们可以断定,在眼球球旁局部注射的视黄酸溶液可被眼局部组织如巩膜、脉络膜或视网膜吸收并聚集。本研究提示形觉剥夺导致的眼球后极部组织中视黄酸水平的上升可能同样是以相同的途径增强了脉络膜组织中MMP-2的表达,进而导致MMP-2与TIMP-2之间的平衡被打破,即视黄酸可能是形觉剥夺性近视眼脉络膜中一个重要的调控MMP-2与TIMP-2之间平衡的上游因子。外界视觉环境的变化可能是通过调控眼球后极部组织中视黄酸的含量而来调控脉络膜组织中MMP-2的表达,进而影响脉络膜的厚度,最终导致了动物眼屈光度和眼轴的变化,当然,巩膜组织中的视黄酸含量增加导致的巩膜变薄和生物力学的变化也可能同时参与了近视的发生和发展。
4参考文献
1Hung LF,Wallman J,Smith EL.Vision-dependent changes in the choroidal thickness of macaque monkeys〔J〕.Invest Ophthalmol Vis Sci,2000;41(6):1259-69.
2Chu SC,Hu DN,Yang SF,etal. Uveal melanocytes produce matrix metalloproteinases-2 and-9 in vitro〔J〕.Pigment Cell Res,2004;17:636-42.
3Alexander JP,Bradley JM,Gabourel JD,etal. Expression of matrixmetalloproteinases and inhibitor by human retinal pigment epithelium〔J〕.Invest Ophthalmol Vis Sci,1990;31(12):2520-8.
4Fitzgerald ME,Wildsoet CF,Reiner A.Temporal relationship of choroidal blood flow and thickness changes during recovery from form deprivation myopia in chicks〔J〕.Exp Eye Res,2002;74(5):561-70.
5McFadden SA,Howlett MHC,Mertz JR.Retinoic acid signals the direction of ocular elongation in the guinea pig eye〔J〕.Vis Res,2004;44(2):643-53.
6林文生,张农,张颂文,等.全反式维甲酸可增强大鼠系膜细胞基质金属蛋白酶2-表达〔J〕.中华病理学杂志,1999;28(6):456-7.
7王剑锋,刘双珍.外源性视黄酸对鸡后巩膜基质MMP-2表达作用的动态观察〔J〕.国际眼科杂志,2004;4(5):815-9.
8Mertz JR,Howlett MHC.Retinoic acid from both the retinaand choroids influences eye growth〔J〕.Invest Ophthalmol Vis Sci,1999;40(4):849-55.
9Mertz JR,Wallman J.Choroidal retinoic acid synthesis:a possible mediator between refractive error and compensatory eye growth〔J〕.Exp Eye Res,2000;70(4):519-27.
〔2014-10-11修回〕
(编辑李相军)
