武汉地区结核分枝杆菌异烟肼耐药性的基因分析
2015-12-24孙战强,陈高瞻,余晓丽
通信作者:余晓丽(1963-),女,教授,E-mail:Liyan1181@126.com.
武汉地区结核分枝杆菌异烟肼耐药性的基因分析
孙战强1,2,陈高瞻1,余晓丽1
(1.武汉轻工大学 生物与制药工程学院,湖北 武汉430023;2. 上海凯地生物科技有限公司,上海 20025)
摘要:探讨武汉地区结核分枝杆菌临床分离株异烟肼耐药相关基因突变特征。通过PCR直接序列分析法对异烟肼耐药相关基因katG基因及mabA-inhA启动子进行序列分析。20株异烟肼敏感株未检测到突变,49株耐药株中有40株(81.6%)存在突变,其中katG的S315突变率占主导优势,突变率为73.5%(36/49),mabA-inhA启动子区核苷酸置换率为8.2% (4/49),其中一株同时检测到了katG和mabA-inhA启动子区的突变。此外,还检测到了一种国内尚未报道的katG突变类型P232S(CCG→TCG)。研究证实katG基因和mabA-inhA启动子的突变与异烟肼耐药相关,为进一步研究结核分枝杆菌异烟肼耐药性奠定了基础。
关键词:结核分枝杆菌;异烟肼;耐药; katG基因; mabA-inhA启动子
收稿日期:2015-06-05.
作者简介:孙战强(1971-),男,主管检验师,E-mail:sunzhanqiang12@126.com.
基金项目:上海市科委科技支撑项目(12441903300).
文章编号:2095-7386(2015)03-0037-04
DOI:10.3969/j.issn.2095-7386.2015.03.008
中图分类号:R 378
Genotypic analysis of isoniazid-resistantMycobacterium
tuberculosisisolates recovered from Wuhan
SUNZhan-qiang1, 2,CHENGao-zhan1,YUXiao-li1
(1.School of Biology and Pharmaceutical Engineering,Wuhan Polytechnic University,Wuhan 430023,China;
2.Shanghai Kaidi Biological Technology Co., LTD, Shanghai 20025, China)
Abstract:To evaluate the characteristic of genes associated with isoniazid-resistant Mycobacterium tuberculosis clinical isolates collected from Wuhan, China. Methods The genotypes of katG and mabA-inhA promoter that associated with isoniazid-resistance were analyzed with PCR direct sequencing. Results Of the 49 isoniazid-resistant clinical isolates, 43 strains(81.6%) carried mutations in the target genes, and no mutations was found in the 20 isoniazid- sensitive strains. The mutations in katG gene at codon 315 was the most frequent mutations up to 73.5%(36/49)in all. The nucleotide replacement rate of mabA-inhA promoter was 8.2% (4/49).One isolate strain was detected mutations both in katG mabA-inhA promoter. Moreover, we found a new mutant type P232S(CCG→TCG). Conclusions This study confirmed that mutations in the katG and mabA-inhA promoter related with isoniazid-resistance, and provided clues to explore the mechanism of isoniazid -resistant Mycobacterium tuberculosis.
Key words:Mycobacteriumtuberculosis(MTB); isoniazid (INH); drug resietance;katGgene;mabA-inhApromoter
1引言
结核病是单一致病菌引起死亡最主要的原因,而耐药及耐多药结核分枝杆菌的出现,使结核病的治疗更加困难。异烟肼( INH)是结核病治疗中最主要的一线药物,同利福平、吡嗪酰胺一样是现代结核病控制策略(DOTS)短期化疗的基础药物[1]。研究表明在所有结核分枝杆菌(Mycobacteriumtuberculosis, MTB)耐药分离株中INH的耐受率最高,因此阐明INH的耐药机制可以为MTB耐药快速检测和个体化治疗提供关键理论基础。INH耐药机制十分复杂,受多个基因调控,国内外研究表明,MTB中80%-90%的INH耐药主要是由于katG基因和mabA-inhA启动子区发生突变所致[2]。本研究旨在探讨武汉地区结核分枝杆菌临床分离株异烟肼耐药相关基因突变特征,为进一步深入地研究阐明结核分枝杆菌异烟肼的耐药机制奠定基础。
2材料与方法
2.1材料
结核分枝杆菌标准菌株H37Rv(ATCC 27294)购自中国药品生物制品检验所。69株临床分离株依据《结核病诊断实验室检验规程》[3]在武汉医疗救治中心进行分枝杆菌分离培养、菌种鉴定、绝对浓度法药敏试验(异烟肼,利福平,链霉素,乙胺丁醇),共收集到20株全敏感株,49株INH耐药株。
2.2方法
2.2.1DNA提取
分离菌株接种在改良罗氏(L-J)培养基上,37 ℃培养至对数生长期,根据文献[4]报道,采用酶解和传统酚/ 氯仿抽提法提取各菌株DNA。运用Thermo NANODROP 2000微量分光光度计测定DNA的浓度和纯度,-20℃保存备用。
2.2.2聚合酶链反应(PCR)及测序
参照文献[5]报道,设计菌种鉴定引物INS-F(5’- CGTGAGGGCATCGAGGTGGC -3’)和INS-R(5’- GCGTAGGCGTCGGTGACAAA-3’),该引物可以特异性地扩增结核分枝杆菌的IS6110的保守区域。根据结核分枝杆菌标准株H37Rv的katG基因和mabA-inhA启动子区序列设计引物katG-F(5’- TCGGCGATGAGCGTTACAG-3’) 、katG-R(5’- CGTCCTTGGCGGTGTATTG-3’) ,产物长度为459 bp;mabA-inhA-F(5’-CGTCCTTGGCGGTGTATTG-3’) 及mabA-inhA-R(5’-ATCCCCCGGTTTCCTCCGGT-3’) ,产物长度为248 bp。PCR体系(50 μl):10×PCR Buffer(5 μl),四种dNTP 各0.2 mmol/L,上下游引物各0.5 μmol/L,Taq酶2.5 U,DNA模板2 μg;PCR扩增条件:94 ℃预变性5 min,94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸1 min,30个循环,72 ℃延伸10 min。PCR 产物经1%的琼脂糖凝胶电泳后,目的基因片段阳性产物经Omega-PCR纯化试剂盒纯化后送上海美吉生物医药科技有限公司测序。
2.2.3DNA序列分析
测序数据通过Clustal W软件并结合Blast数据库(http://blast.ncbi.nlm.nih.gov/Blast.cgi?CMD=Web&PAGE_TYPE=BlastHome)进行比对分析。
3结果
3.1PCR产物
以结核分枝杆菌标准株H37Rv为对照,应用引物INS-F和INS-R引物扩增结核分枝杆菌临床分离株均得到245bp片段,进一步确认这些分离株为结核分枝杆菌。此外,其他耐药基因也均扩出相应的DNA片段(见图1)。

M:DNA Marker;1:标准株H37Rv;2~8:临床分离株 图1 katG基因PCR扩增产物电泳图
3.2katG基因的突变检测
20株INH敏感株未检测到突变,49株耐药株中有40株(81.6%)发生了突变(见表1)。katG S315突变占主导优势,突变率为73.5%(36/49)。S315最主要的突变类型为:S315T(AGC→ACC)见图2,另外还包括S315N(AGC→AAC)突变。此外,本实验中还发现了katG基因232位点突变,类型为P232S( CCG→TCG),未发现缺失突变。
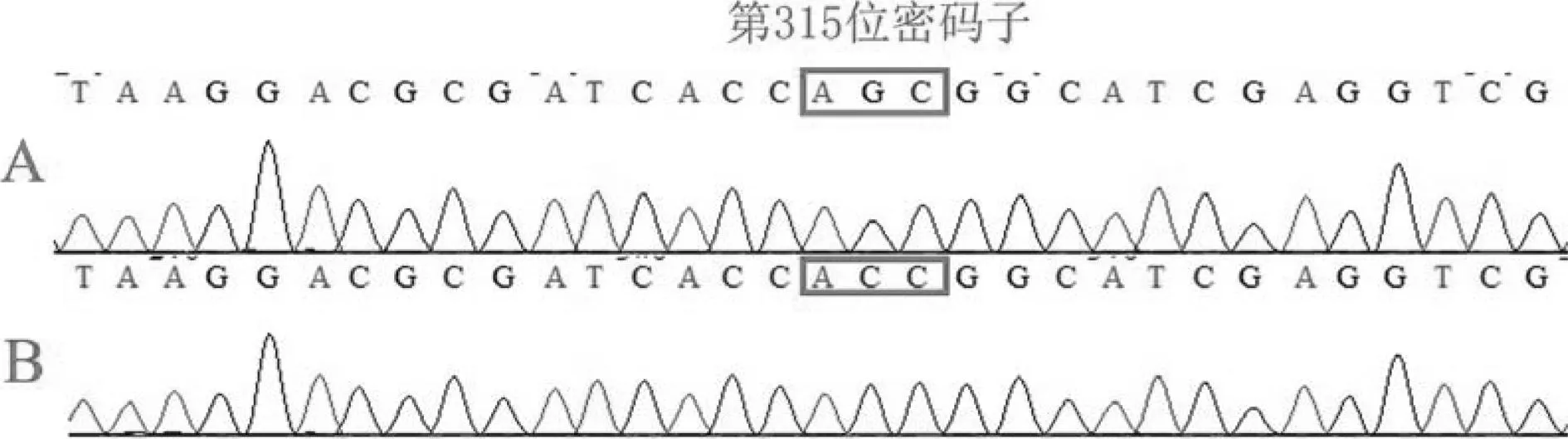
A.结核分枝杆菌H37Rv标准株的katG基因片段序列; B.耐INH临床分离株的katG基因点突变S315T(AGC→ACC) 图2 临床分离菌株katG基因315位点突变情况
表1INH耐药株katG及mabA-inhA突变情况 (n=49)

基因基因突变特征位点核苷酸氨基酸突变菌株数目占比katG232CCG→TCGPro(P)→Ser(S)1/49315AGC→ACCSer(S)→Thr(T)35/49AGC→AACSer(S)→Asn(N)mabA-inhA启动子-15C→T4/49
3.3mabA-inhA启动子序列突变检测
mabA-inhA启动子区的序列分析结果:有4株(8.2%)检测到了碱基替换,均为C-15T突变。其中一株与katG基因S315联合突变。
4讨论
Zhang[6]等最早研究MTB的耐药的分子机制,提示是由于katG基因的完全缺失造成了过氧化氢-过氧化物酶的活性丧失从而导致MTB对INH的高耐药。近年来,INH的耐药机制引起了人们的广泛关注,研究发现INH耐药分子机制与某些基因突变有关。但是由于地理区域的不同,INH耐药基因多态性及其突变频率存在差异。研究报道称,katG315的突变率范围为:40%—93.6%,常见的katG315突变有AGC→ACC, AGC→AGG, AGC→AAC, AGC→GGC等[7-8]。本实验中来自武汉地区的49例INH耐药株katG315(Ser→Thr)突变占主导优势,突变率为71.4%(35/49),与国内外绝大多数报道一致[9-10];35株katG315突变中97.1%(34/35)的突变类型为AGC→ACC,仅一例为 AGC→AAC,上述结果表明katG315突变是武汉地区结核分枝杆菌INH耐药的主要原因。
本次研究中在katG基因232位点发现突变类型是CCG→TCG(Pro→Ser)。Cardoso[11]等首次报道了该突变位点232位Pro→Arg,且和mabA-inhA启动子联合突变,表现为INH高耐药;国内陈杨[12]等报道了该位点的突变(CCG→CAG ,Pro→Gln),且与inhA基因的 16位(CGT→GGT)联合突变,表现为耐多药。Ando[13]等新近研究发现katG基因232位点另一突变类型Pro→Ser,并称其与INH低耐药有关。而本实验中的232(CCG→TCG,Pro→Ser)突变并未与检测中的其他几个INH耐药基因发生联合突变。有研究表明,结核分枝杆菌katG基因的缺失与耐INH有关,且多为高耐药,但该基因缺失的报道并不多见[14]。本研究未发现耐药菌株中katG基因的缺失。
由inhA编码的烯酰基乙酰载体蛋白(inhA)是INH的作用靶点之一,研究表明inhA上游启动子区域mabA-inhA突变能引起inhA表达的增加,从而提高药靶浓度并产生INH耐药性[15]。多数研究发现INH耐药株中mabA-inhA突变率为4.3%—34.4%[16]。本研究中耐药株mabA-inhA突变率为8.1%(4/49),且都为单独的C-15T替换,可见在武汉地区mabA-inhA基因突变率相对较低,而且mabA-inhA和katG联合突变率更小,可能只需要katG基因一个突变就足以满足耐INH。
此外,本研究49株表型耐药株中有6例在以上检测基因中均未发生突变,这些菌株可能在这些基因的其他区段发生突变或者还存在其他耐药机制目前正在进一步研究之中。
本研究首次阐明了武汉地区结核分枝杆菌INH耐药株的耐药分子机制主要为katG基因发生点突变,为进一步研究结核分枝杆菌INH的耐药机制奠定了一定的基础,为MTB耐药分子诊断积累区域差异性数据。但是完全阐明武汉地区结核分枝杆菌INH耐药的机制还需要大规模检测样本,并且进行多基因的研究。
参考文献:
[1]WHO. Multidrug and extensively drug-resistant TB (M/XDR-TB): 2010 global report on surveillance and response[R]. Geneva:World Health Organization,2010.
[2]Wade M M, Zhang Y. Mechanisms of drug resistance in Mycobacterium tuberculosis[J]. Front Biosci,2004,9:975-994.
[3]中国防痨协会基础专业委员会. 结核病诊断实验室检验规程[M]. 北京:中国教育文化出版社,2006:49-51.
[4]张舒林, 伍学强, 祖燕, 等. 应用PCR-SSCP检测耐异烟肼结核分支杆菌KatG基因突变[J]. 中国卫生检验杂志,2000,10:185-187.
[5]Zhang S L, Qi H, Qiu D L, et al. Genotypic analysis of multidrug-resistant Mycobacterium tuberculosis isolates recovered from central China[J]. Biochem Genet,2007,45:281-290.
[6]Zhang Y, Heym B, Allen B,et al. The catalase-peroxidase gene and isoniazid resistance of Mycobacterium tuberculosis[J]. Nature,1992,358:591-593.
[7]Mokrousov I, Narvskaya O, Otten T, et al. High prevalence of KatG Ser315Thr substitution among isoniazid-resistant Mycobacterium tuberculosis clinical isolates from northwestern Russia, 1996 to 2001[J]. Antimicrob Agents Chemother,2002,46:1417-1424.
[8]Boonaiam S, Chaiprasert A, Prammananan T, et al.Genotypic analysis of genes associated with isoniazid and ethionamide resistance in MDR-TB isolates from Thailand[J]. Clin Microbiol Infect,2010,16:396-399.
[9]乐军, 张曼, 张红梅, 等. 结核分枝杆菌临床分离株异烟肼耐药相关基因突变的分子特征[J]. 中华微生物学和免疫学杂志,2006,26:950-955.
[10]Yao C, Zhu T, Li Y, et al.Detection of rpoB, katG and inhA gene mutations in Mycobacterium tuberculosis clinical isolates from Chongqing as determined by microarray[J]. Clin Microbiol Infect,2010,16:1639-1643.
[11]Cardoso R F, Cooksey R C, Morlock G P, et al. Screening and characterization of mutations in isoniazid-resistant Mycobacterium tuberculosis isolates obtained in Brazil[J]. Antimicrob Agents Chemother,2004,48:3373-3381.
[12]陈杨, 陈玲, 张泓, 等. 耐异烟肼结核分枝杆菌及其katG与inhA基因突变的研究[J]. 中国抗生素杂志,2010,35:788-792.
[13]Ando H, Kondo Y, Suetake T, et al. Identification of katG mutations associated with high-level isoniazid resistance in Mycobacterium tuberculosis[J]. Antimicrob Agents Chemother,2010,54:1793-1799.
[14]Varela G, Gonzalez S, Gadea P, et al.Prevalence and dissemination of the Ser315Thr substitution within the KatG enzyme in isoniazid-resistant strains of Mycobacterium tuberculosis isolated in Uruguay[J]. J Med Microbiol,2008,57:1518-1522.
[15]Muller B, Streicher E M, Hoek K G, et al. inhA promoter mutations: a gateway to extensively drug-resistant tuberculosis in South Africa? [J].Int J Tuberc Lung Dis,2011,15:344-351.
[16]Yumin H, Sun Y J, Sinyew W,et al. Contribution of dfrA and inhA mutations to the detection of isoniazid-resistant Mycobacterium tuberculosis isolates[J]. Antimicrob Agents Chemother,2009,53:4010-4012. C, Zhu T, Li Y, et al.Detection of rpoB, katG and inhA gene mutations in Mycobacterium tuberculosis clinical isolates from Chongqing as determined by microarray[J]. Clin Microbiol Infect,2010,16:1639-1643.
[11]Cardoso R F, Cooksey R C, Morlock G P, et al. Screening and characterization of mutations in isoniazid-resistant Mycobacterium tuberculosis isolates obtained in Brazil[J]. Antimicrob Agents Chemother,2004,48:3373-3381.
[12]陈杨, 陈玲, 张泓, 等. 耐异烟肼结核分枝杆菌及其katG与inhA基因突变的研究[J]. 中国抗生素杂志,2010,35:788-792.
[13]Ando H, Kondo Y, Suetake T, et al. Identification of katG mutations associated with high-level isoniazid resistance in Mycobacterium tuberculosis[J]. Antimicrob Agents Chemother,2010,54:1793-1799.
[14]Varela G, Gonzalez S, Gadea P, et al.Prevalence and dissemination of the Ser315Thr substitution within the KatG enzyme in isoniazid-resistant strains of Mycobacterium tuberculosis isolated in Uruguay[J]. J Med Microbiol,2008,57:1518-1522.
[15]Muller B, Streicher E M, Hoek K G, et al. inhA promoter mutations: a gateway to extensively drug-resistant tuberculosis in South Africa? [J].Int J Tuberc Lung Dis,2011,15:344-351.
[16]Yumin H, Sun Y J, Sinyew W,et al. Contribution of dfrA and inhA mutations to the detection of isoniazid-resistant Mycobacterium tuberculosis isolates[J]. Antimicrob Agents Chemother,2009,53:4010-4012.
