香灰菌与银耳混合培养过程中酶系的相互作用*
2015-12-24赖淑芳郑珠霜王丽芬陈桂珍朱传进万春和孙淑静胡开辉
林 辉,赖淑芳,郑珠霜,王丽芬,陈桂珍,朱传进,万春和,孙淑静,胡开辉**
(1.福建农林大学生命科学学院,福建 福州 350002;2.福建农林大学(古田)菌业研究院,福建 古田 352200;3.古田食用菌研发中心,福建 古田 352200)
〈生理生化〉
香灰菌与银耳混合培养过程中酶系的相互作用*
林 辉1,2,赖淑芳1,郑珠霜1,王丽芬2,3,陈桂珍2,3,朱传进2,3,万春和1,孙淑静1,2,胡开辉1,2**
(1.福建农林大学生命科学学院,福建 福州 350002;2.福建农林大学(古田)菌业研究院,福建 古田 352200;3.古田食用菌研发中心,福建 古田 352200)
该实验对纯香灰菌(Hypoxylon sp.)培养及其与银耳(Tremella fuciformis)混合培养过程中的纤维素酶、淀粉酶、漆酶等培养原料基质降解相关酶变化规律进行了比较与分析。结果表明,香灰菌长满菌袋后淀粉酶与漆酶活力随着培养时间的延长逐渐下降,木聚糖酶活力在培养至15 d达到最大值5 000 U·mL-1,纤维素酶活力培养至5 d后,稳定在40 U·mL-1左右,可以有效降解木料培养基。银耳与香灰菌混合培养后,在固体培养过程中,其中胞外可溶性蛋白浓度和木聚糖酶、纤维素酶活力变化规律与香灰菌纯培养相似,而淀粉酶在培养至第10天达到最大值2 253 U·mL-1,其胞外可溶性蛋白浓度与木聚糖酶、纤维素酶和淀粉酶活力显著提高,漆酶活力培养至第5天达到最大值71.33 U·mL-1,随后迅速下降,蛋白酶活力变化情况与淀粉酶相似,第15天酶活力达到最大值3.05 U·mL-1。说明混合培养中两者具有互作效应,可以有效促进胞外蛋白的分泌,提高基质降解相关酶活力,促进菌体对基质的降解,提高养分的供应。该结果可为银耳优良菌种的选育,两者的配比以及栽培料的选择与优化提供科学依据。
香灰菌;银耳;基质;降解;酶活
银耳(Tremella fuciformis)生活史复杂,自身没有分解木质纤维素与淀粉基质的能力,不能单独在木质纤维素上生长,只有与香灰菌进行共生培养时才能正常生长发育[1-2]。香灰菌(Hypoxylonsp.)是银耳的伴生菌,在银耳的培养过程中,可以分泌基质降解酶对栽培料基质进行分解,为银耳的生长发育提供营养。目前有学者[3-4]就香灰菌菌液对银耳菌丝生长作用以及不同香灰菌传代次数对银耳栽培的影响进行了研究,表明了香灰菌的代谢产物对银耳菌丝的稳定生长具有促进作用,并且随着香灰菌传代次数的增加羧甲基纤维素酶以及漆酶活性逐渐下降,混合培养过程中银耳的生长周期延长,产量与品质下降。但纯香灰菌与银耳混合培养过程中的基质降解相关酶活测定还未见报道。为寻找银耳栽培过程中银耳菌种的质量与稳定性以及与相关酶活的变化规律,建立银耳菌种质量的内控指标,以及为栽培料的优化提供判断依据,本研究对纯香灰菌培养及其与银耳混合培养过程中的纤维素酶、漆酶体系(漆酶、锰过氧化物酶和木质素过氧化物酶)、木聚糖酶、淀粉酶和蛋白酶活力的变化进行了测定,探明香灰菌培养过程中自身的基质分解相关酶活性,旨在揭示香灰菌在银耳培养中在基质分解方面的作用,找出香灰菌基质降解酶活变化与银耳生长发育的相关性,从而为优良菌种的选育和银耳栽培料的优化提供借鉴。
1 材料与方法
1.1菌种
银耳纯菌丝和香灰菌纯菌丝,均为福建农林大学(古田)菌业研究院保藏菌种。
1.2试供培养基
1.2.1 PDA培养基(加富)
土豆(去皮)200g、葡萄糖20g、蛋白胨3g、酵母提取粉3g、硫酸镁1.5g、磷酸二氢钾1.5g、维生素B10.1g、琼脂20g,加水定容至1000 mL,pH自然。
1.2.2 栽培料
木屑培养基:木屑78%、麦麸20%、蔗糖1%、石膏1%(含水量60%)。
1.3试验方法
1.3.1 培养方法
栽培料加水混匀后过夜堆料,次日装袋(35 mm×90 mm),每袋湿重60%。121℃灭菌120 min,冷却后按5%接入原种,接种完的菌袋放入25℃恒温培养箱进行培养,一定时间测定菌袋失重情况。
1.3.2 样品前处理
自菌袋长满菌丝开始(25 d),每隔5 d将整袋栽培料倒入不锈钢锅,充分混匀即可取样,每次取样3袋。准确称取20 g栽培料置于60℃烘干箱烘干,另取10 g用于制备粗酶液。
1.3.3 粗酶液的制备
栽培料全部取出搅拌均匀后,称取10 g于100 mL三角瓶中,加入50 mL双蒸水,150 r·min-1,25℃恒温振荡摇床培养90 min后,经纱布过滤后,9 000 r·min-1离心15 min,取上清液即为粗酶液。
1.4pH值与总糖的测定
将制备好的粗酶液,使用pH计测定溶液pH值。准确称取0.15 g烘干后的栽培料,加入1.5 mL浓盐酸与5 mL双蒸水沸水浴3 h。8 000 r·min-1离心10 min,取上清后将沉淀重复2次操作,汇合3次溶液后定容至50 mL,使用硫酸-苯酚法测定糖浓度。
1.5酶活测定
相关酶活测定:纤维素酶活力参照文献 [5];木聚糖酶活力参照文献 [6];中温-α-淀粉酶活力测定参照文献 [7];中性蛋白酶活力测定参照文献 [8];漆酶活力测定参照文献 [9];木质素过氧化物酶活力测定参照文献 [10];锰过氧化物酶活力测定参照文献 [11]。
2 结果
2.1香灰菌满袋后的总糖与pH值变化分析
香灰菌满袋后培养过程中栽培料总糖和pH值的变化情况见图1。
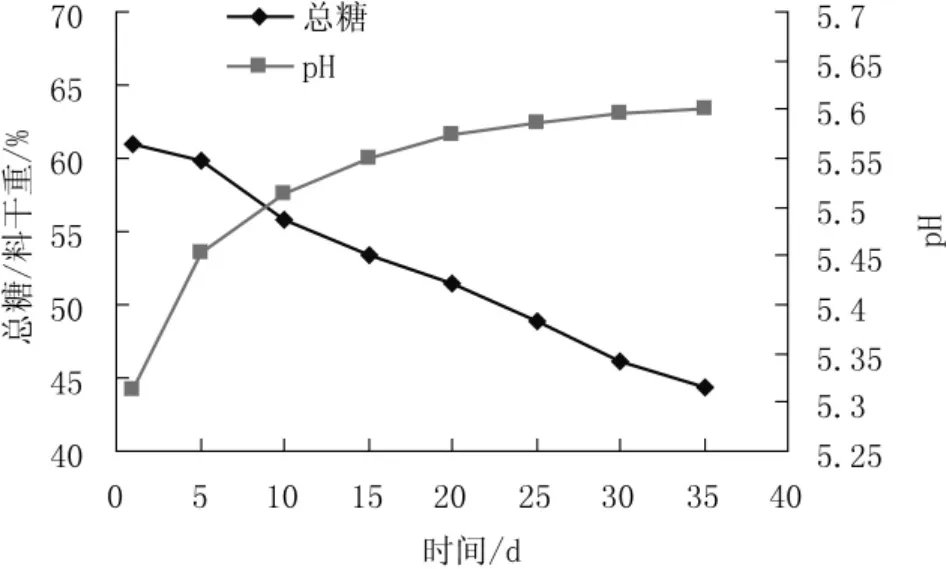
图1 香灰菌培养过程中栽培料总糖与pH变化曲线Fig.1 Changing of total sugar and pH in cultivation material duing the Hypoxylon sp.cultivation
由图1可以看出,香灰菌菌袋长满后,栽培料中的总糖随着培养时间的延长,以0.5%(总糖/料干重)左右的速率逐渐下降。培养35 d,总糖共下降18%。其总糖用于菌丝的生长、呼吸作用以及合成相关物质,菌袋重量也有所减轻。栽培料的pH略有上升,从菌袋长满到第10天,pH值由5.3上升到5.5,栽培10 d以后,菌袋中栽培料的pH值基本不变,维持在5.5~5.6。pH值的略有上升可能是香灰菌在生长过程中对栽培料中的酸性物质的利用所致。
2.2香灰菌满袋后与银耳混合培养过程中的酶活力变化
2.2.1 纤维素酶活力变化分析
香灰菌满袋后与银耳混合培养过程中纤维素酶活力变化情况见图2。

图2 香灰菌与银耳混合培养过程中纤维素酶活力变化Fig.2 Activity of cellulase produced by Hypoxylon sp.and Tremella fuciformis mixed cultivation incultivation material
木屑、棉籽壳是银耳栽培的主要原料,具有丰富的纤维素与半纤维素,在纤维素酶的作用下纤维素分解成葡萄糖,为碳水化合物和氨基酸构成细胞物质和供给食用菌生长发育所需的能量,因此纤维素酶活的变化可反映食用菌对培养基质的降解能力。
由图2可以看出随着香灰菌菌袋菌丝长满,纤维素酶酶活力从20 U·mL-1至长满菌袋5 d后稳定在40 U·mL-1左右,持续的降解纤维素为菌丝生长和出菇提供所需营养。香灰菌与银耳混合培养后,培养至20 d纤维素酶活力达到210 U·mL-1,纤维素酶活力显著提高,相对于香灰菌纯培养酶活力提高了5倍,由此可见银耳与香灰菌混合培养后,大大提高了银耳对原料中纤维素的降解能力。
2.2.2 淀粉酶和木聚糖酶活力变化分析
香灰菌满袋后与银耳混合培养过程中淀粉酶和木聚糖酶酶活力变化情况见图3。

图3 香灰菌与银耳混合培养过程中淀粉酶、木聚糖酶活力变化Fig.3 Activity of amylase and xylanase produced by Hypoxylon sp. and Tremella fuciformis mixed cultivation in cultivation material
淀粉酶可以降解栽培料中的淀粉类多糖为单糖作为菌丝生长、代谢的能量来源,木聚糖酶可有效降解培养料中如麸皮等糠麸类饲料中的木聚糖(木聚糖是半纤维素的主要成分)。
由图3可以看出香灰菌在生长过程中具有很高的淀粉酶活力。当菌丝长满菌袋后,淀粉酶活力高达900 U·mL-1,随着培养时间的增加,栽培料中的淀粉酶活力逐渐减低,培养至25 d后降低到500 U·mL-1左右。香灰菌与银耳混合培养后淀粉酶酶活力大大提高,培养至第10天淀粉酶活力稳定在2 253 U·mL-1左右。香灰菌在长满菌袋后木聚糖酶活力高达1000 U·mL-1,随着培养时间的延长,其酶活力继续升高,培养至15 d后达到最大值5 000 U·mL-1;混合培养后的木聚糖酶活力明显提高,培养至25 d酶活力高达8 575 U·mL-1。可见混合培养后,淀粉酶与木聚糖酶酶活力相对香灰菌单纯培养提高明显,加强了银耳培养过程中对栽培料中淀粉类和半纤维素多糖的利用。
2.2.3 中性蛋白酶活力与胞外可溶性蛋白浓度变化
香灰菌满袋后与银耳混合培养过程中胞外可溶性蛋白浓度和中性蛋白酶酶活力变化情况见图4。
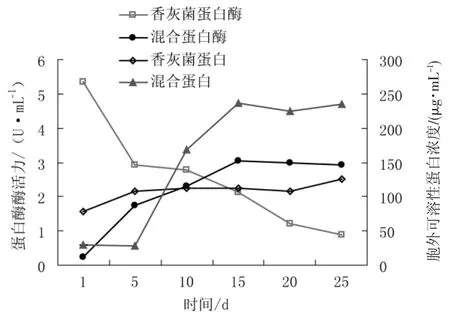
图4 香灰菌与银耳混合培养过程中胞外可溶性蛋白浓度和蛋白酶酶活力变化Fig.4 Extracellular soluble protein and activity of protease produced by Hypoxylon sp.and Tremella fuciformis mixed cultivation in cultivation material
蛋白酶可以将蛋白质分解成氨基酸、尿素、氨等小分子化合物后进行吸收并将其用于合成蛋白质和核酸,为菌丝的生长提供原料。其中纤维素酶、淀粉酶和木聚糖酶等基质降解酶均为胞外可溶性降解酶,基质降解酶是胞外酶的重要组成成分之一。
由图4可以看出香灰菌在培养过程中蛋白酶活力很低,菌丝刚长满袋后仅达到5 U·mL-1;随着培养时间的延长,栽培料中蛋白酶活力逐渐降低,长满25 d后其酶活力仅在1 U·mL-1左右。香灰菌与银耳混合培养后,胞外蛋白酶活力最大仅达到3.05 U·mL-1。香灰菌菌袋长满后胞外可溶性蛋白浓度由80 μg·mL-1增长到110 μg·mL-1,而混合培养后,栽培料中的胞外可溶性蛋白浓度培养至第15天后稳定在235 μg·mL-1左右。可见相对香灰菌纯培养,香灰菌与银耳的混合培养可以降低胞外蛋白酶的合成,提高胞外酶的分泌。
2.2.4 漆酶体系(漆酶、木质素过氧化物酶、锰过氧化物酶)酶活力
香灰菌满袋后与银耳混合培养过程中漆酶、锰过氧化物酶和木质素过氧化物酶活力变化情况见图5、表1。
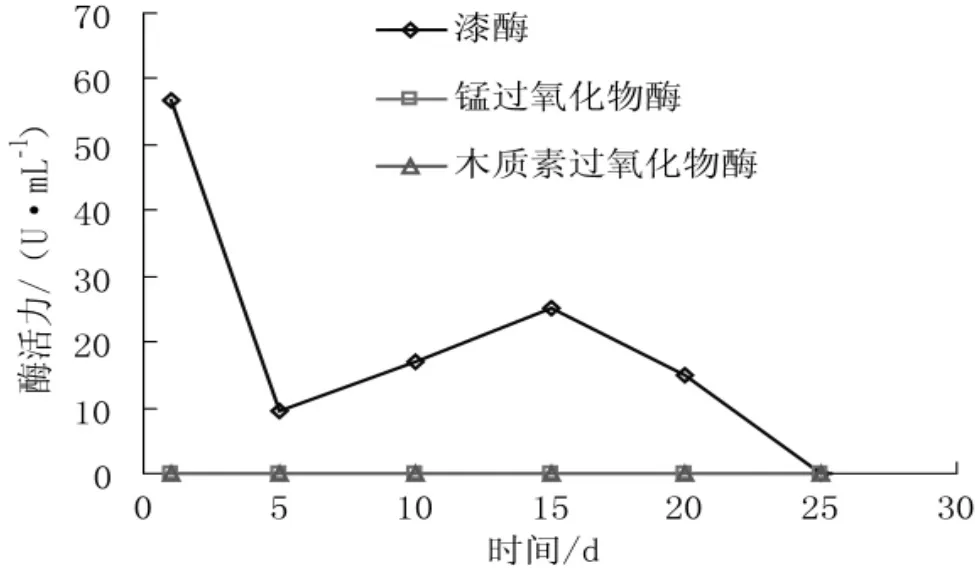
图5 香灰菌培养过程中漆酶、锰过氧化物酶与木质素过氧化物酶酶活力变化Fig.5 Activity of laccase,manganese peroxidase and lignin peroxidase produced by Hypoxylon sp.in cultivation material
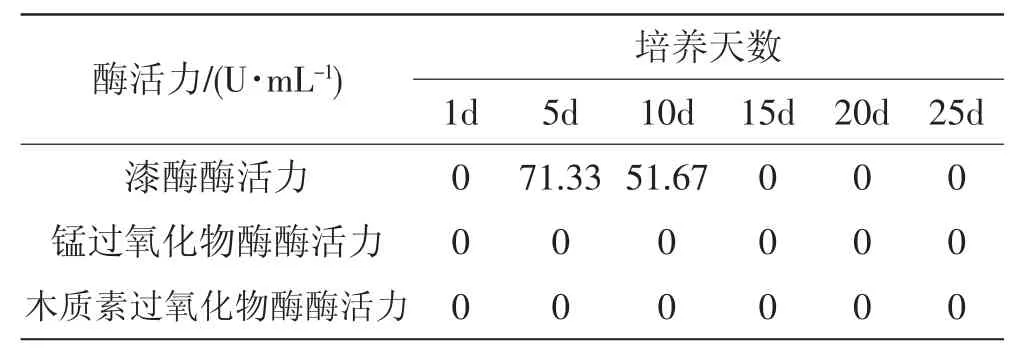
表1 混合培养过程胞外蛋白浓度和基质降解相关酶活力Tab.1 Activity of laccase,manganese peroxidase and lignin peroxidase produced by Hypoxylon sp.and Tremella fuciformis mixed cultivation in cultivation material
漆酶是一种与木质素分解相关的多酚氧化酶,在食用菌生长周期中分解木质素,为菌丝提供营养物质,另外,漆酶的代谢产物醌能增强菌种的抗病能力,抑制杂菌的污染。
通过测定表明,香灰菌菌丝长满菌袋后随着培养时间的增加漆酶活性迅速降低,5 d~15 d酶活力提高到25 U·mL-1后又迅速降低,培养至25 d后检测不到漆酶的活性。混合培养后,漆酶活力在培养第5天达到最大值71.33 U·mL-1后迅速下降,培养至15 d后检测不到漆酶的活性。两种培养均检测不到锰过氧化物酶与木质素过氧化物酶酶活性。
3 小结与讨论
王玉万和梁勤等[12-13]对银耳研究结果表明,银耳自身在培养过程中并不能产生纤维素酶并对栽培料中的纤维素进行降解,混合培养后栽培料中的纤维素酶酶活有了明显的提高。本实验研究结果与这一结论一致:香灰菌纯培养的栽培料中纤维素酶活力达到40 U·mL-1,混合培养后的酶活提高到210 U·mL-1。菌袋长满后的栽培料中,木聚糖酶与淀粉酶活力最为明显,其中木聚糖酶酶活力高达5 000 U·mL-1以上,淀粉酶酶活力也达到2 000 U·mL-1。其中淀粉酶活力随着培养时间的延长而逐渐下降。这一结果与对香菇与金针菇等食用菌的研究结果相似[14-15]。而木聚糖酶活力达到最高后随着培养天数延长逐渐下降,这一现象又与对平菇研究结果相似[16-17]。本文漆酶活性迅速下降后,又有所上升,这一规律与对金针菇的培养过程研究相似[15]。因此香灰菌在培养过程中可以很好地在栽培过程中降解木质素、木聚糖和淀粉等基质中的碳源,有效地为银耳的生长提供营养。相对香灰菌纯培养,混合培养栽培料中除了蛋白酶酶活力外,纤维素酶酶、木聚糖酶、漆酶和淀粉酶酶活力均显著提高,加快对基质的降解速率。这一现象也与王玉万研究结果相同[12]。从胞外可溶性蛋白浓度可以看出,混合培养栽培料的蛋白浓度高达香灰菌纯培养的2倍,因此混合培养可以明显促进菌丝胞外酶的分泌,提高基质降解酶的活力。以上的结果可以说明香灰菌在基质降解方面对银耳有重要作用,两者之间存在着明显的相互作用关系,可以为两者优良菌种的选育,栽培过程中两者之间的配比以及培养料的优化提供重要的参考。
[1]汪国莲,陈立国,陈明.银耳菌丝生长营养条件的初步研究[J].食用菌,2000,22(4):12-14.
[2]邓优锦,王庆福,陈介斌,等.具有拮抗关系的18个香灰菌株遗传差异性分析[J].食用菌学报,2014(2):25-31.
[3]贺元川,陈仕江,杨勇,等.香灰菌传代对银耳栽培的影响[J].江苏农业科学,2014,42(4):201-202.
[4]陈明.香灰菌浸出液对银耳菌丝体生长的影响[J].食用菌,2000,22(4):10-11.
[5]郭艳艳,阮玲云,冯宏昌,等.不同营养条件下斑玉蕈菌丝生长及产酶特性[J].菌物学报,2014,33(3):697-705.
[6]李科,孙彦平,蔡小波,等.黄曲霉产木聚糖酶条件优化及酶解产物初步分析[J].生物技术通报,2009(S1):348-351,363.
[7]孙静,耿慧莉,莫德馨.中温α-淀粉酶活性的定量测定[J].教学仪器与实验,2009(11):44-45.
[8]SB/T 10317-1999,蛋白酶活力测定法[S].
[9]胡开辉,刘建忠,孙淑静,等.斑玉蕈育种中漆酶转化体系建立的初步研究[J].菌物学报,2010,29(4):528-535.
[10]田林双.木质素降解相关酶类测定标准方法研究[J].畜牧与饲料科学,2009,20(10):13-14.
[11]徐建中,胡开辉,孙淑静,等.不同类型食用菌产木素氧化酶系能力比较与分析[J].江西农业大学学报,2011,33(2):375-380.
[12]王玉万.银耳及其伴生菌营养生理生态研究[J].应用生态学报,.1993,4(1):59-64.
[13]梁勤,温文婷,叶小金,等.银耳种质资源遗传多样性及酶学特性分析[J].西南农业学报,2010,23(4):1194-1198.
[14]闫训友,史振霞,杨静,等.壳聚糖对香菇菌丝生长代谢关键酶的影响[J].浙江农业学报,2012,24(3):455-458.
[15]高金权,刘朝贵,李成琼,等.稻草秸秆栽培金针菇基质降解特性研究[J].中国农学通报,2005,21(12):260-264.
[16]庄庆利,李冠军,申进文,等.平菇栽培种培养过程中胞外酶活性变化的研究[J].河南农业大学学报,2010,44(2):163-166.
[17]刘朝贵,高金权,李成琼,等.糙皮侧耳(Pleurotus ostreatus)降解转化稻草秸秆研究[J].西南农业大学学报:自然科学版,2006,28(2):258-263.
紫芝基因组精细图完成发布
据悉,教育部长江学者创新团队负责人陈士林领导的课题组日前完成了紫芝基因组精细图,该精细图含有12条染色体,序列总长度达到48.86兆碱基,编码大约1.5万个基因。相关研究成果已发表在自然出版集团旗下期刊《科学报道》上。
课题组研究表明,紫芝在生长发育中具有多种防御机制,包括基因组防御机制和化学防御机制,紫芝的化学防御功能可能受到多种基因调控机制影响。基因组防御对于维持物种遗传物质稳定性具有重要意义,而化学防御与其次生代谢产物的合成与调控具有密切关系。
灵芝基因组图谱的公布为开展灵芝三萜等有效成分的合成研究提供了便利,随着这些合成途径的逐步解析,使得通过合成生物学合成灵芝有效成分成为可能。同时,对灵芝生长发育和抗病抗逆关键基因的研究探索,将推动灵芝的基因组辅助育种研究,加速灵芝新品种的培育,为灵芝产业的蓬勃发展提供了科技支撑。
中国食用菌商务网
2015.07.03
Interaction Rules of Enzyme System between Hypoxylon sp.and Tremella fuciformis
LIN Hui1,2,LAI Shu-fang1,ZHENG Zhu-shuan1,WANG Li-fen2,3,CHEN Gui-zhen2,3, ZHU Chuan-jin2,3,WAN Chun-he1,SUN Shu-jing1,2,HU Kai-hui1,2
(1.College of Life Sciences,Fujian Agriculture and Forestry University,Fuzhou 350002,China; 2.Edible Fungi Research Institute (Gutian),Fujian Agriculture and Forestry University,Gutian 350002,China; 3.Edible Fungi Research and Development Center of Gutian,Gutian 350002,China)
In order to study the interaction rules of enzyme system betweenHypoxylonsp.andTremella fuciformis,the substrate degradation related enzymes about theHypoxylonsp.and mixed strain cultivation (T.fuciformisandHypoxylonsp.)were determined and analyzed.The results showed that the amylase and laccase activity ofHypoxylonsp.reduced during the culture period.However,the activity of xylanase reached the peak of 5 000 U·mL-1at the 15thday,while the cellulase activity increased and reached the peak of 40 U·mL-1after the 5thday,which could degrade the wood culture medium effectively.Thechanging rule of soluble extracellular protein concentration,xylanase and cellulase in medium by mixed strain cultivation had similar trend withHypoxylonsp.,while the amylase reached the peak of 2 253 U·mL-1at the 10thday and soluble extracellular protein concentration and xylanase,cellulose,amylase activity increased obviously.The laccase activity reached the peak of 71.33 U·mL-1at the 5thday and then declined sharply.The changing rule of proteinase activity was similar with amylase which reached the peak of 3.05 U·mL-1at the 15thday.The results indicated that the interaction in mixed strain cultivation not only promote the secretion of extracellular protein,but also it improve the substrate degradation related enzyme activity,and increase the ability of degradation for the substrate.The results lay good foundation for theT.fuciformisbreeding,such as the ratio and the culture medium for theHypoxylonsp.and theT.fuciformis.
Hypoxylonsp;Tremella fuciformis;substrate;degradation;enzyme activity
S646.9
A
1003-8310(2015)04-0057-05
10.13629/j.cnki.53-1054.2015.04.014
福建省高校产学研重大项目(2013N5004);珍稀食用菌品种(真姬菇、绣球菌) 创新与自动化生产技术产业化工程项目(2014S1477-8);福建省食用菌产业技术重大研发平台(2014N2101)。
林辉(1990-),男,硕士,助理研究员,主要从事食用菌开发与利用方面研究。E-mail:13489943760@163.com
**通信作者:胡开辉(1962-),男,硕士,教授,主要从事真姬菇的应用、开发与工厂化方面研究。E-mail:2692609765@qq.com
2015-04-20
