不同潮位下角果木的种群发展趋势分析
2015-12-21钟军弟成夏岚周贤熙刘锴栋袁长春
钟军弟,成夏岚,周贤熙,刘锴栋,袁长春,陈 燕
(1.岭南师范学院 生命科学与技术学院,广东 湛江524048;2. 华南农业大学 动物科学学院,广东 广州510642)
不同潮位下角果木的种群发展趋势分析
钟军弟1,成夏岚1,周贤熙2,刘锴栋1,袁长春1,陈 燕1
(1.岭南师范学院 生命科学与技术学院,广东 湛江524048;2. 华南农业大学 动物科学学院,广东 广州510642)
为了阐明不同潮位下角果木Ceriops tagal种群的发展趋势,合理利用与保护现有的红树物种资源,通过野外样方调查与室内分析的统计学方法对不同潮位下的角果木种群进行调查,综合比较分析不同潮位生境下角果木的年龄结构,存活曲线与及静止生命表的差异,探究不同潮位生境下角果木的种群结构变化规律与发展趋势。构建高潮位生境下的种群动态的时间序列预测模型,预测了角果木种群其随着时间的变化种群的发展状况。结果表明,高潮位的生境下角果木种群呈现典型的“金字塔”增长型结构,种群处于不断增长趋势,存活曲线表现为凹形,即为DeeveeyⅢ。在低潮位的生境下,角果木的幼苗较多,但在生长发育过程中出现生长“瓶颈”现象,幼苗较难发育至成体树,种群呈现衰退的趋势。时间序列模型表明,高潮位生境下的角果木处于健康稳定的增长状态,种群的生长发育随着时间的变化而呈现出稳定的增长态势。
角果木;年龄结构;静止生命表;存活曲线;潮位;时间序列模型
红树林是分布于热带和亚热带海岸潮间带的木本植物群落,为海陆交界的特殊湿地生态系统,其不仅能够防风消浪、保护堤岸、净化环境 ,还是鱼、虾、蟹等海洋生物生长繁殖的场所和栖息地,而且能够有效的保护生物物种的多样性,其对人类具有重要的生态学及社会经济价值[1]。然而随着沿海地区人口增长、工业和城市的发展,我国海岸带被过度地开发利用,越来越多的红树林生境遭到破坏,一些红树物种已濒临灭绝。因此,保护和恢复红树林生态系统是当前亟需解决的重要问题。
角果木Ceriops tagal红树科角果木属,为红树林的主要组成物种,主要分布于海南省和广东省徐闻县两地,分布面积不大,数量较少。随着经济的发展及海岸的过度利用与开发,角果木受到人为破坏较为严重。据报道,由于人为的破坏,广西壮族自治区和台湾省两地的角果木已绝迹[1]。因此,亟需对角果木采取必要的保护措施。植物种群数量动态变化是植物个体生存能力与外界环境相互作用的结果。不同生境条件下植物种群年龄结构、静止生命表以及存活曲线不仅可以反映种群现实状况,还可以展现植物种群与环境抗争的关系,这对于濒危植物的保护和利用研究具有重要意义[2-4]。目前,关于角果木方面的研究主要侧重于化学成分分析及应用[5-8]和植物生理生态[9-10]方面的研究,而对角果木的种群生态学领域方面的研究却较少。因此,我们通过野外样方调查与室内统计学的方法研究角果木的种群生态,综合比较分析角果木在不同潮位生境下的种群年龄结构,静止生命表,存活曲线的变化规律与及分析种群时间序列模型,研究角果木在不同潮位下的生长状况与种群发展趋势,了解角果木对异质环境的生态适应机制,以进一步保护现有红树资源以及为人工造林提供理论基础。
1 研究地自然概况
研究区位于广东省西南部雷州半岛的最南端—徐闻县迈陈镇官曹海岸红树林区域内,地处东经 109°95′750"~109°96′945",北 纬20°56′970"~20°59′335" 之间。全境属亚湿润季风气候,夏季高温多雨,年平均气温24.1 ℃。极端最高温38.8 ℃,极端最低温2.2℃[11],无霜期360 d以上。太阳辐射强年平均日照时数为2 078.7 h,年蒸发量达1 934 mm。年平均降雨量1 163.1 mm,年均相对湿度83 %。4~9月为雨季,10~3月为干季。全年盛行东风和东南风,5~10月间受到热带风暴和台风影响。该区域成土母质主要为玄武岩、古浅海沉积物和河流冲积物等,土壤类型以砖红壤为主,另有滨海沙土、潮砂泥土等[12]。研究区红树林面积约40 hm2,主要由角果木、红海榄Rhizophora stylosa、白骨壤Aricennia marina等种类组成。角果木可形成单优群落或与红海榄、白骨壤形成混合群落。群落外貌呈黄绿色或黄绿与深绿镶嵌,群落高近2 m,郁闭度约70 %,群落林下幼苗和幼树较多,天然更新较好。
2 研究方法
2.1 样地设置与调查
潮位是指受潮汐影响周期性涨落的水位。根据当地的实际情况,在没有涨潮的情况下,我们将距离海水边缘的100 m以内作为高潮位,在距离海水边缘100 m以外作为低潮位。我们选择的研究样地设于祖国大陆的最南端湛江市徐闻县迈陈镇的红树林区域内。经踏查后,我们选择生境条件具有代表性和一致性、植物个体生长良好、人为破坏较少的样地进行调查,相邻样方之间的距离至少相隔100 m,以便获得更为准确的有代表性的实验数据。不同潮位生境下各选取5 m×5 m大小样方8个,并在每个样方内对角果木进行每木检尺,记录不同潮位样方内的角果木的数量,基径,冠幅,盖度。同时记录每个样方的经纬度,海拔等环境因子。
2.2 种群年龄结构分析方法
由于木本植物生长周期长,不可能也没有必要追踪所有的个体命运。通过现实不同年龄阶段的个体数量可以推测种群时间上的动态过程。植物中稳定的径级结构类似于稳定的年龄结构,所以使用胸径或者基茎结构代替年龄结构来分析乔木种群的结构及其动态特征[3,13]。鉴于角果木树种植株较矮,分枝部位较低等特点,我们选择按其基径大小进行分级:基径≤2. 5的植株为幼苗,定为Ⅰ级,2. 5 cm以上的植株则基径每增加5 cm为一级。据此,根据我们样方数据可以将角果木种群分为7级。把从小到大的树木径级顺序看作时间顺序关系,即第1径级对应第1龄级,第2径级对应第2龄级,如此一一对应, 最后统计各龄级株数。
2.3 静止生命表编制
生命表是研究种群数量动态的重要工具,其不仅可以反映种群的数量动态及发展趋势,还可以反映种群与环境间的相互关系以及它们在群落中的作用和地位[14-15]。因此,编制种群生命表和生存分析是研究种群数量动态的有效方法,
编制静态生命表要满足三个假设:一是种群的数量是静态不变的,即密度不变;二是年龄组合是稳定的,即种群的年龄结构与时间无关,各年龄的比例不变; 三是个体的迁移是平衡的,即迁入与迁出的数量是一致的。特定时间生命表一般包含如下栏目:
x:单位时间年龄等级的中值;ax:在x龄级内现有个体数;
lx:在x龄级开始标准化存活个体数(一般转化为1 000);
ln(lx):存活数标准化的对数;ax(%):存活百分数;
dx:各区间死亡数标准化;qx:死亡率 ;
Lx:从x到x+1龄级间隔期间还存活的个体数;
Tx:从x龄级到超过x龄级的个体总数;
ex:进入x龄级个体的生命期望寿命;
各项都是相互关联的, 其关系如下:

由于研究的角果木种群为天然林,而且是由“ 空间推时间”,“横向导纵向”,因此调查所得的数据并不完全满足编表的所有假设。因此,在生命表的编制中会出现死亡率为负值的情况。对这种情况,吴承祯等认为虽然生命表分析中产生的一些负的dx 值与数学假设技术不符,但仍能提供有用的生态学记录,即表明种群并非静止不动,而是在迅速发展或衰落之中[4]。因此,我们参照江洪[16](1992)生命表编制过程中采用的匀滑修正技术对统计数据进行处理得到了生命表。
2.4 存活曲线的绘制
存活曲线是研究种群动态的主要手段之一,通过对存活曲线的分析,可以了解掌握种群动态的本质及其内在规律[17-18]。存活曲线绘制的方法有两种,一是以存活量的对数值lglx 为纵坐标,以年龄为横坐标作图;另一种方法是用存活数量对年龄作图,但年龄用平均寿命期望的百分离差来表示[10,19]。本文以存活量lx 为纵坐标,以年龄级为横坐标作图。
2.5 种群动态的时间序列预测模型
时间序列分析是时间数列预测和回归预测方法的综合形式。该方法并不是用于分析因果关系,而是侧重于根据时间序列过去的变化规律推测种群今后的发展趋势。参照一次性移动平均法对角果木的种群龄级结构进行模拟和预测[20],模型构建的公式为:
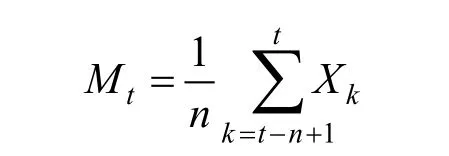
式中:n表示需要预测的未来时间年限,Mt表示未来n年时t龄级的种群大小,Xk为当前k龄级的种群大小,分别对未来2个和4个龄级时间后进行模型构建。
3 结果与分析
3.1 不同潮位生境下角果木种群年龄结构
由图1可知,高潮位的角果木的种群年龄结构属于典型的“金字塔”增长型,幼苗发育成成体树较多,种群发展处于稳定增长的趋势,说明高潮位是适合角果木生长的生境。在低潮位的生境下,虽然幼苗很多,但幼苗只发育至第三阶段,即角果木的生长几乎只停留于幼苗与幼树阶段,没有发育成为成体树,角果木的生长呈现“瓶颈”状态,这说明低潮位的环境不适宜角果木的生长发育。可见,潮位影响着角果木的生长状况,低潮位不利于角果木的生长,高潮位有利于角果木的生长。
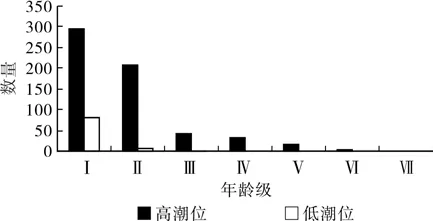
图1 不同潮位生境下角果木种群年龄结构Fig. 1 Different tide population age structure of the Ceriops tagal
3.2 不同潮位生境下角果木种群的静态生命表
由表1可知,在低潮位生境下角果木种群的期望寿命变化不大,即表示Ⅰ级至Ⅲ级的角果木的期望寿命是基本相同的,角果木的生长发育过程中,其种群Ⅰ级至Ⅲ级的死亡率几乎不变,说明角果木生长发育后期的不稳定。高潮位生境下角果木从Ⅰ级至Ⅶ级的期望寿命是呈不断上升的趋势,表明随着角果木的生长发育,期望寿命单调性增高,这说明角果木的死亡率越来越低,角果木种群生长发育后期相当的稳定。
3.3 不同潮位生境下角果木种群的存活曲线
图2可知,低潮位的生境下,角果木的种群生长多停留在幼苗、幼树阶段,种群内幼苗、幼树死亡率较高,较难向成树发育过渡,呈现生长“瓶颈”状态,种群发展表现衰退的趋势。而在高潮位的生境下,种群在各个年龄阶段均有分布,并在一定的年龄范围内,种群存活率不变。超过一定年龄范围后,种群存活率不断上升。因此,角果木的存活曲线是属于典型的凹形增长模式,即DeeveeyⅢ。由此可以看出,高潮位适合角果木的生长发育,有利于角果木的种群发展,而低潮位不利于角果木的种群发展。
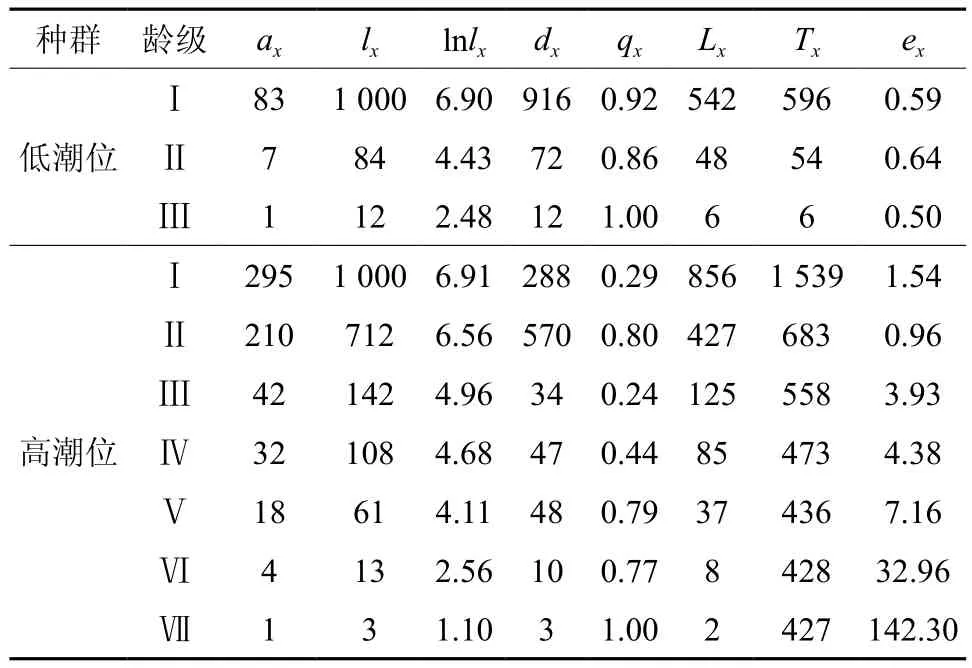
表1 不同潮位生境下种群角果木静止生命表Table 1 Life table of Ceriops tagal population in different tides
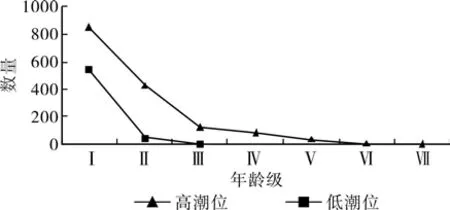
图2 不同潮位生境下角果木存活曲线Fig. 2 Survival curve of Ceriops tagal
3.4 种群动态的时间序列预测模型
为了更深入了解高潮位生境下今后的变化趋势,我们建立了高潮位生境下的时间序列预测模型。由于低潮位生境下的角果木年龄级较少,没法建立四龄级后的时间序列模型,两龄级后的数据只有两个,所建立的模型不具有可靠性,故没有建立低潮位的时间序列模型。高潮位时间序列分析(图3)表明,随着时间序列的变化,无论是两龄级后,还是四龄级后,角果木种群中所有老龄树木均高于前一年龄级的种群数量,均呈典型的增加趋势,说明高潮位生境下的角果木种群的幼中龄植株数所占整体的比例高,能使后期个体得到一定的补充,此生境下的角果木的种群处于健康稳定的增长状态。但是我们同样看到,无论是两龄级后还是四龄级后,Ⅵ和Ⅶ年龄级的数量都呈不变的趋势,这表明处于这两个年龄级的角果木数量有限,树木很难往更高级的年龄级生长发育。因此我们需要加大对Ⅵ和Ⅶ年龄级的保护措施,维持种群的持续发展。
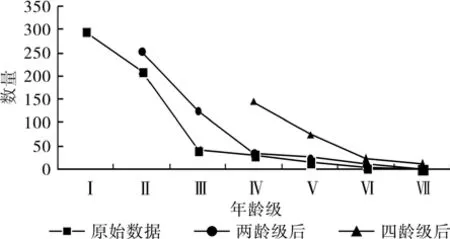
图3 高潮位生境下角果木种群数量动态时间序列预测Fig.3 Time sequence prediction of Ceriops tagal population in high tide
4 结论与讨论
平均期望寿命又称“生命期望值”或“平均余命”,是一种对种群的某一阶段年龄种群今后尚能生存的平均寿命的有效的预测[21-22]。不同生境下植物种群表现不同的生长发育状况。研究不同潮位生境下的角果木种群数量动态,可以反映角果木在不同生境下其生长状况及种群发展趋势。有研究表明,高潮位生境高程较高,土壤盐度较低潮位的低,土壤N、P养分及含水量较低潮位生境的高[23]。高潮位生境下的角果木种群密度较大,种群生长发育状况较好,低潮位生境下角果木种群生长发育较差,且出现幼苗难向成树过度的生长“瓶颈”现象。主要原因可能为高潮位生境下,海水淹浸时间较短,频率较低,生境光照时间较长,生境内生物类群丰富,滩涂沉积物较多,土壤有机质、N和P等养分丰富。另外,高潮位生境下沟渠淡水注入较多,土壤含水量较高,生境盐度较低,角果木的生长渗透压与环境渗透压相似,其并没受到环境渗透压的胁迫抑制。因此,嗜热、喜光、中等耐盐特性[24]的角果木种群生长发育较好,各个年龄阶段个体数量较多,种群呈稳定增长趋势。相反,低潮位生境下,海水淹浸时间较长,频率较高,生境光照时间较短,生境内盐度和渗透压较高,土壤水分、有机质、N和P等养分较低,角果木个体生长受到生境渗透压和养分的胁迫抑制。因而,角果木种群生长发育状况较差,幼苗虽具有一定数量,但成树个体数较少,种群呈“瓶颈”的衰退状态。由于角果木为“胎生”植物,其胚轴在一定的时期内角果木幼苗的生长提供所需要的养分,因此,角果木幼苗生长受到的外界干扰相对较少,即使在高盐度、低养分的低潮位下仍具有一定的数量。不同潮位下的角果木有着不同的生长趋势的现象是由于多种原因综合作用的结果,但是具体是为哪种原因在起着主要主导的作用,这还需要更为深入与详细的研究。
角果木种群在不同潮位生境下其生长发育趋势是不同的。在低潮位的生境下,角果木幼苗的生长遇到“瓶颈”状态,幼苗只能发育至Ⅲ级,生命期望寿命呈不变的趋势,幼苗无法发育成为成体树,种群呈衰退的发展趋势。在高潮位的生境下,角果木的生长态势很好,幼苗的存活率较高,种群年龄结构是典型的“金字塔”增长型,生命期望寿命是呈不断上升的趋势,死亡率不断降低,幼苗发育成成体树的可能性很大,生长发育趋势良好。角果木时间序列趋势分析也表明高潮位生境下的种群是处于健康成熟的状态下。因此,我们要重点保护高潮位的生境环境,防止过度破坏高潮位环境,影响角果木的生存。同时,在适宜种植的高潮位生境上引种造林,扩大角果木的分布面积,维持其种群的延续。
[1]王文卿, 王 瑁. 中国红树林[M]. 北京 : 科学出版社, 2007.
[2]李俊清. 森林生态学[M]. 北京:高等教育出版社, 2006.
[3]张文辉, 王延平, 康 永.濒危植物太白红杉种群年龄结构及其种群发展趋势分析[J].生物多样性, 2004, (3): 361-369.
[4]吴承祯, 洪 伟. 珍稀濒危植物长苞铁杉种群生命表分析[J].应用生态学报, 2000, 13(4): 333-336.
[5]张 炎, 邓志威, 高大翔, 等.中国红树植物角果木的化学成分 [J]. 药学学报 , 2005, 40(10): 935-939.
[6]王 慧,吴 军. 十雄角果木树皮的二萜类化学成分研究[J].中草药 , 2013, 44(5): 532-536.
[7]王子熹. 角果木提取物对人卵巢癌裸鼠皮下移植瘤的作用[J]. 齐鲁医学杂志 , 2013, 28(3): 199-202.
[8]孙 蓓,王秀美,邱文生,等.红树植物角果木Dolabrane型二萜对Lewis荷瘤小鼠的抗肿瘤作用[J].临床肿瘤学杂志,2010, 15(2): 127-131.
[9]臧 剑,符秀梅,王鹤鸣. 角果木根系盐胁迫的防御机理[J].热带生物学报, 2013, 4(2): 160-164.
[10]陈 燕, 谢正生, 刘锴栋. 角果木和红海榄对低温胁迫的生理响应差异研究[J].西北农林科技大学学报:自然科学版,2013, (3): 69-74.
[11]杨彩福, 焦学志. 近50年来湛江市年极端气温统计分析[J].广东气象 , 2008,30(3):14-16.
[12]钟来元, 郭良珍. 雷州半岛南部土壤肥力综合评价及空间变异[J]. 广东海洋大学学报, 2012, 32(3):76-81.
[13]李先琨,苏宗明,向悟生.濒危植物元宝山冷杉种群结构与分布格局[J]. 生态学报,2002(12): 2246-2254.
[14]程红梅. 大蜀山大叶榉种群数量动态研究[J].中南林业科技大学学报 , 2009, 29(3) : 65-69.
[15]陈远征, 马祥庆,冯丽贞,等.濒危植物沉水樟的种群生命表和谱分析[J].生态学报,2006, 26(12):4267- 4272.
[16]江 洪.云杉种群生态学[M].北京:中国林业出版社,1992.
[17]Edward S. Deevey J R. Life tables for natural populations of animals[J]. The Quarterly Review of Biology, 1947,22(4):283-314.
[18]闫桂琴, 赵桂仿, 胡正海. 秦岭太白红杉种群结构与动态的研究[J].应用生态学报,2011 ,32(8): 824-828.
[19]Edward S. Deevey J R. Life tables for natural populations of animals[J]. The Quarterly Review of Biology, 1947,22(4):283-314.
[20]申仕康, 马海英, 王跃华, 等. 濒危植物猪血木自然种群结构及动态[J].生态学报, 2008, 25(5): 2404-2412.
[21]Edward S. Deevey J R. Life tables for natural populations of animals[J]. The Quarterly Review of Biology, 1947,22(4):283-314.
[22]于大炮, 周 莉, 董百丽, 等. 长白山北坡岳桦种群结构及动态分析[J]. 生态学杂志, 2004, (5): 30-34.
[23]潘 宇. 芦苇与互花米草种群沿潮位梯度的镶嵌分布格局及其与生境关系研究[D]. 上海:华东师范大学, 2012.
[24]陈 燕,刘锴栋, 陈粤超,等. 角果木的育苗与造林技术[J].广东林业科技,2013, 29(3): 1-5.
The analysis of Ceriops tagal development trend in different tide level
ZHONG Jun-di1, CHENG Xia-lan1, ZHOU Xian-xi2, LIU Kai-dong1, YUAN Chang-chun1, CHEN Yan1
(1. School of Life Science and Technology, Lingnan Normal University, Zhanjiang 524048, Guangdong, China;2. College of Animal Science, South China Agricultural University, Guangzhou 510642, Guangdong, China)
In order to clarify the trend of theCeriops tagalpopulation age structure and future development, protect and make full use of the existing resources, we gathered datum under different tidal field quadrat randomly, explored different tidal habitatsCeriops tagalpopulation structure variation with the age structure ofCeriops tagal, survival curve and still life table, and predicted the population development ofCeriops tagalwith the time series prediction model of population dynamics.The result shows thatCeriops tagalpopulations are increasing with the structure of typical pyramid and the concave survivorship curves, namely for Deeveey Ⅲ. In the low tide level there are a certain number ofCeriops tagalseedlings, however, it is more impossible for the seedlings to develop into mature trees, showing a growth bottlenecks and a trend of population decline. Time sequence prediction indicates thatCeriops tagalpopulation is in the stable development in high tide environment.
Ceriops tagal; Age structure ofCeriops tagalpopulation; Life table; Survival curve; Tide level; Time sequence prediction
S718.54
A
1673-923X(2015)10-0074-05
10.14067/j.cnki.1673-923x.2015.10.013
2014-02-10
国家星火计划项目(2013GA780090,2011GA780061);广东省林业科技创新项目(2013KJCX011-03,2015KJCX025)
钟军弟,硕士
陈 燕,副教授;E-mail:yanch211@163.com
钟军弟,成夏岚,周贤熙,等. 不同潮位下角果木的种群发展趋势分析[J].中南林业科技大学学报,2015, 35(10): 74-78.
[本文编校:吴 彬]
