冀北山区不同林龄落叶松人工林林分结构及更新
2015-12-21王冬至张冬燕张志东牟洪香张玉珍
王冬至,张冬燕,张志东,牟洪香,张玉珍
(1.河北农业大学 林学院,河北 保定 071000;2.河北农业大学 商学院,河北 保定 071000)
冀北山区不同林龄落叶松人工林林分结构及更新
王冬至1,张冬燕2,张志东1,牟洪香1,张玉珍1
(1.河北农业大学 林学院,河北 保定 071000;2.河北农业大学 商学院,河北 保定 071000)
林分结构在很大程度上决定了林分稳定性和更新苗的存活与分布,因此,研究林分结构与更新格局至关重要。本文运用角尺度、大小比数、趋势面分析对不同林龄落叶松人工林结构及更新苗分布格局进行了统计分析。结果表明:27 a生落叶松林为随机分布,32 a和39 a生落叶松林均呈均匀分布;更新苗在不同林龄落叶松林下均呈聚集分布,且与林龄呈负相关关系。不同林龄林分结构随着林龄的增加林木分化呈递减的趋势,随着林龄增加林木在空间结构上的分化趋于稳定;不同林龄更新苗在空间结构上呈现出随着林龄的增加更新苗分化不明显;草本植物对更新苗的数量和分布影响最大,在人工林经营过程中应结合抚育间伐并对林下灌草及枯落物进行及时清理,促进更新苗生长发育。
人工林;空间结构参数;趋势面分析;更新
林分空间结构一般指林木分布格局及其属性在空间上的排列方式,它不仅反映了种群的空间属性和数量特征[1-3],还对物种生长、繁殖、死亡以及森林资源管理和利用等都具有显著影响[4-6],而且在很大程度上决定了林分的稳定性及发展的可能性和林分经营空间的大小[7]。有学者[8-10]利用
角尺度、大小比数和混交度3个林分空间结构参数对栓皮栎林、蒙古栎林等林分空间结构特征进行了研究,表明不同的林分结构很可能会影响林下树种的分布[11-13]和林下幼苗更新[14-15],而林下幼苗分布格局将决定未来森林群落的结构[16-17],因此研究林分结构与更新苗格局对实现森林可持续经营至关重要。
林分结构决定着林分功能,有什么样的林分结构,就有相应的林分功能[18]。因此,林分结构与更新研究对森林经营技术研究、森林结构与功能研究具有重要意义。林分结构及更新苗分布格局不仅取决于环境因子的影响,还受其它人为干扰等物理因子的影响,近年来研究林分空间结构的方法迅速发展,如空间自相关分析、空间异质性分析、趋势面分析、地统计学方法和分形方法等[19-20]。林分结构是影响华北落叶松林天然更新的重要因素,而更新苗的空间分布又将决定未来的林分结构[21-22],在研究区不同林龄落叶松人工林内几乎看不到天然更新的落叶松大苗或幼树的存在表明其天然能力较差,本文旨在探索不同林龄落叶松人工林林分结构及更新苗空间分布格局和天然更新规律有助于揭示不同林龄华北落叶松天然更新过程,为可持续经营华北落叶松人工林提供科学依据。
1 研究区概况
研究区位于河北省塞罕坝林场,是阴山山脉与大兴安岭余脉交汇处,典型的山地地形,属寒温带大陆性季风气候,经纬度为118°19′09″~116°50′29″E,41°07′14″~42°58′17″N,海 拔 高度1 022 m~1 884 m,年均气温-1.4~4.7℃,极端温度为30.9℃~-42.8℃,年均降水量380~560 mm,年蒸发量1 350~1 500 mm,年均无霜期59~126 d,年均日照时数为2 368 h,成土母质主要为残坡积母质、坡积母质、冲洪积母质、洪积母质、冲击母质和风积母质等,随着海拔升高其土壤分布为棕壤-褐土-风砂土-草甸土-沼泽土-灰色森林土-黑土等。研究区主要乔木树种有油松Pinus tabulaeformis、白桦Betula platyphylla、 蒙 古 栎Quercus mongolica、五 角 枫Acer elegantulumFang et P. L. Chiu、山杨Populus davidiana、等,林下主要灌木树种有山刺玫Rosa daverica、稠李Prunus padusL.、华北忍冬Lonicera tatarinowii、大叶小檗Berberis ferdinandi-coburgii、栓翅卫矛Euonymus phellomanusLoes等,主要草本植物有草地老鹳草Geanium daharicumDC. Var. alpinum Bat、藜芦Radix RhizomaVeratri Veratrum nigrum L.、地榆Sanguisorba officinalis、唐松草Radix et RhizomaThalictri Sibirici、蒲公英Herba Taraxaci、野蔷薇Rosa multifloraThunb.、苔Carex tristachya等。
2 研究方法
2.1 样地设置
经过对不同林龄落叶松林分实地踏查,在塞罕坝千层板林场和北曼甸林场分别选取不同林龄(27 a、32 a、39 a)落叶松人工林为研究对象,并在0~5°的缓坡上对不同林龄落叶松林分别设置3块50 m×50 m(0.25 hm2)样方作为调查样地并将样地划分为25个10 m×10 m网格进行乔木、灌木及草本调查,主要调查内容乔木层:胸径、树高、冠幅、枝下高、郁闭度、坐标、枯落物厚度及更新苗株数、年龄、地径、苗高及坐标等;在样地内沿对角线分别设置8个2 m×2 m小样方调查灌木及草本的多度、盖度、基径、高度及频度等。
2.2 数据统计
2.2.1 角尺度
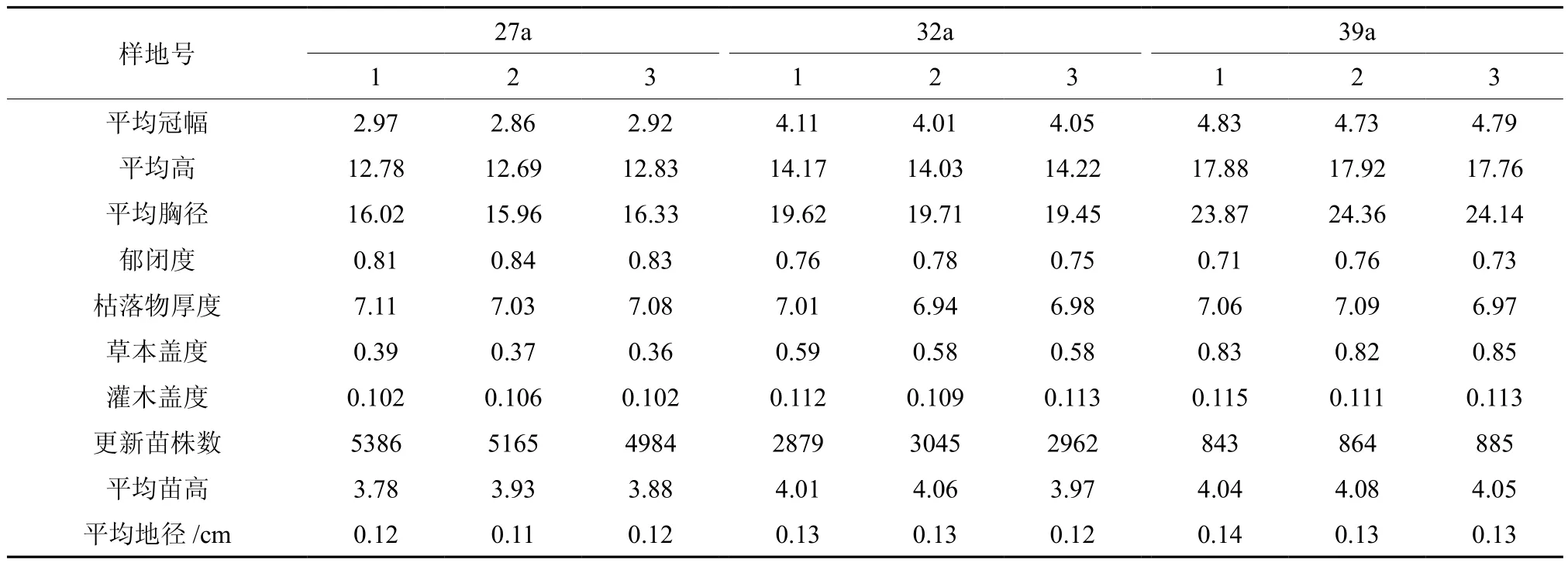
表1 落叶松样地特征Tabbe 1 Stand characteristics of Larix principis-rupprechtii
角尺度用来描述相邻树木围绕参照树i的均匀性,对参照树i的n个相邻最近树而言,均匀分布时其位置分布角应各为360°/n,定义a0(a0=360°/n±360°/10 n)为标准角[23],角尺度 (wi) 定义为 a 角小于标准角a0的个数占所考察的相邻最近树的比例,用下式表示[24]:

式中当第j个a角小于标准角a0则zij= 1,否则zij=0,0<zij<1,zij值越小,分布越均匀。
2.2.2 大小比数
大小比数[23]主要用来描述林木的大小分化程度,其公式为:

式中:kij=1,则参照树i比相邻木j小;kij= 0,则相反,n为最近邻木数Ui值越高,表明周围比参照树大的相邻木越多,参照树的优势度越小;Ui值越低表明周围比参照树大的相邻木就越少,参照树的优势度就越大。
2.2.3 聚集度指数
区域空间实体分布的聚集度指数 J 可以用最邻近距离来[25]计算:

式中:为最邻近更新苗之间平均距离,n为更新苗株数,A为调查的区域面积,当J值越小时,表明更新苗在空间区域分布上越集中,个体关联程度越高 ,更新苗间的屏蔽效应或竞争效应就越大,反之就越小。
2.2.4 趋势面分析
更新苗趋势面分析是通过普通线性模型用最小二乘法来拟合所观测数据,用多元回归方法拟合出观测变量与地理因子的曲面方程,地理因子主要指经纬度或样方空间坐标[20],趋势面分析可直观反应更新苗的空间位置,空间坐标数据包含x2、y2、xy项或更高次项,以保证观测数据能以多变量高次多项式( 趋势面方程) 拟合空间坐标[26]。二次曲面方程模型表达式为V=a1+a2X+a3Y+a4X2+a5XY+a6Y2式中a为回归系数,XY为空间坐标,拟合精度为回归平方和U占总平方和S的比值百分数,即C = U/ S*100,趋势面回归方程包含地理数据表的所有项:通过趋势面分析可以得到更新苗在真实空间中的分布图,使更新苗空间分布和真实空间中的群落趋势图进行对比,可以分析更新苗空间结构产生的原因[20,27]。
3 结果与分析
3.1 林分径阶与更新苗苗龄分布
不同林龄落叶松人工林株数按径阶分布如图1所示,其中27 a 落叶松以16 cm径阶株数分布最多约占林分32%,其次分布在14 cm和18 cm径阶分别占22%、25%;32 a落叶松以20 cm径阶株数分布最多约占林分39%,其次分布在18 cm和22 cm径阶分别占31%、20%;39a落叶松以24 cm径阶株数分布最多约占林分30%,其次分布在22 cm和28 cm径阶分别占16%、20%;在华北落叶松人工林经营过程中不同林龄林分胸径随林龄增加而增大。不同林龄落叶松林下同龄更新苗地径基本没有差异(P>0.05),不同苗龄株数分布在不同林龄林分内差异较大(P<0.05),林分更新幼苗株数随幼苗等级的增大而减少,从图2可以看出幼苗株数随苗龄增大幼苗分布呈倒J型[28],一年生林下更新苗数量与林龄呈负相关关系,在华北落叶松人工林内几乎看不到落叶松幼树的存在。
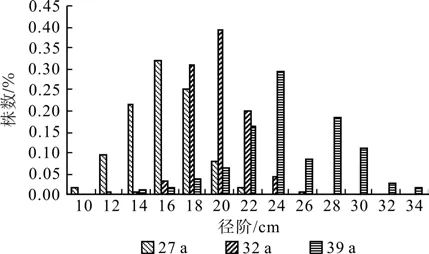
图1 不同林龄径阶株数Fig.1 The number of trees with different diameter classes

图2 不同苗龄更新苗分布Fig.2 Distribution of seedlings with different ages
3.2 林分及更新苗空间格局
不同林龄落叶松人工林及更新苗角尺度分布如图3所示,不同林龄落叶松林平均角尺度分别为:0.486(27 a)、0.431(32 a)、0.421(39 a),不同林龄落叶松林林下更新苗平均角尺度分别为:0.698(27 a)、0.633(32 a)、0.593(39 a),根据随机分布的角尺度取值范围是 0.475<<0.517,小于 0.475 为均匀分布,大于0.517则为集聚分布,27 a 生落叶松林为随机分布,32 a和39 a落叶松林生均呈均匀分布;更新苗在不同林龄落叶松林下均呈聚集分布,且聚集程度较高并与林龄呈负相关关系的趋势。
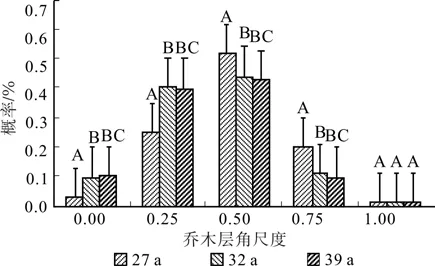
图3 不同林龄落叶松林及更新苗角尺度Fig.3 Uniform angel index of Larix principis-rupprechtii plantations with different ages larch and seedlings
经过数据处理发现不同林龄落叶松人工林小比数分布在0.124~0.264之间,从图4可知不同林龄落叶松林大小比数概率均接近于均匀分布,林分平均大小比数分别为0.521(27 a)、0.493(32 a)、0.471(39 a),表明在不同林龄的林分类型中其空间结构上存在较大差异(P<0.05),随着林龄的增加林木分化呈减弱的趋势,在空间结构上趋于稳定;不同林龄更新苗地径大小比数概率均以绝对劣态苗比率最大分别为0.308(27 a)、0.326(32 a)、0.331(39 a),更新苗平均大小比数为0.614(27 a)、0.621(32 a)、0.624(39 a),更新苗大小比数在空间结构上随着林龄的增加分化趋势并不明显(P>0.05)。
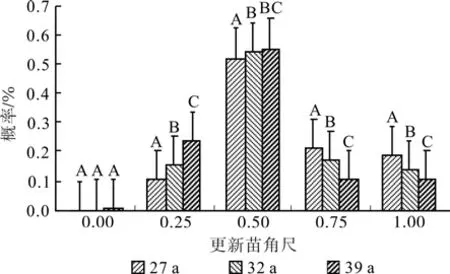
图4 不同林龄落叶松林及更新苗大小比数Fig.4 Neighborhood of Larix principis-rupprechtii plantations with different ages and seedlings
3.3 林分及更新苗趋势面分析
研究中以落叶松胸径和更新苗地径为因变量,以空间坐标XY为自变量分布对不同林龄落叶松及林下更新苗进行了趋势面回归分析并对落叶松和更新苗的空间分布格局进行了模拟,在模拟方程过程中一次多项式和三次多项式拟合精度较低均在0.4以下,二次多项式拟合精度较高(大于0.5)且具有更好的拟合效果,因此选用二次趋势面空间分析结果进行分析,不同林龄落叶松及更新苗趋势面回归方程如表2所示。

表2 趋势面回归方程Table 2 Regression equation of trend surface analysis
不同林龄落叶松林聚集度指数分别为6.213(27 a)、6.868(32 a)、7.061(39 a),林下更新苗均呈聚集分布其聚集度指数分别为2.725(27 a)、3.674(32 a)、6.807(39 a),聚集度指数愈小聚集程度愈高,从图5来看,不同林龄落叶松林在空间结构上随着林龄增加呈均匀分布趋势,从图6来看,不同林龄落叶松林内更新苗分布及生长发育不同,从更新苗模拟图可以看出27年生落叶松林下更新苗主要集中在阳坡且生长较好,32a生落叶松林下更新苗聚集在阴坡且生长较好;39a生落叶松林更新苗聚集度较低趋于均匀分布,更新苗聚集度指数随着林龄增加其聚集度呈下降趋势。
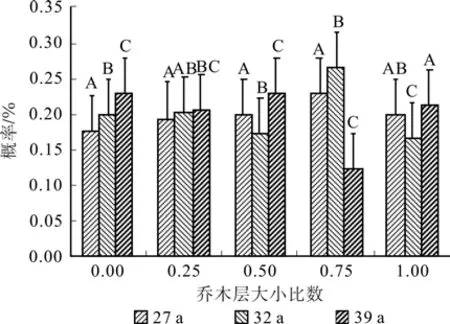
图5 落叶松可视化趋势面分布图Fig.5 Visualization of trend surface maps of Larix principis-rupprechtii
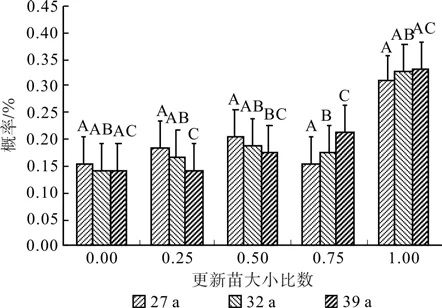
图6 更新苗可视化趋势面分布图Fig.6 Visualization of trend surface maps of seedlings
3.4 林下植被与更新
不同林龄落叶松人工林下的植被分布对幼苗天然更新具有重要作用,利用SAS统计软件对不同林龄林下的更新苗数量、枯落物厚度、草本和灌木的盖度、草本和灌木的均匀度指数进行了ANOVA分析,结果如表3所示,不同林龄林下更新苗数量差异显著(P<0.05),且与林龄呈负相关关系,随着林龄的增加不同林龄林下枯落物厚度呈增加趋势但差异不显著(P>0.05),不同林龄林下草本植物盖度及均匀度分布指数差异显著(P<0.05),并与林龄呈正相关关系,不同林龄林下灌木盖度及均匀度分布指数差异不明显(P>0.05)。因此研究表明,在研究区不同林龄落叶松人工林林下影响幼苗更新数量及更新格局的主要因素为草本植物的盖度和均匀度。
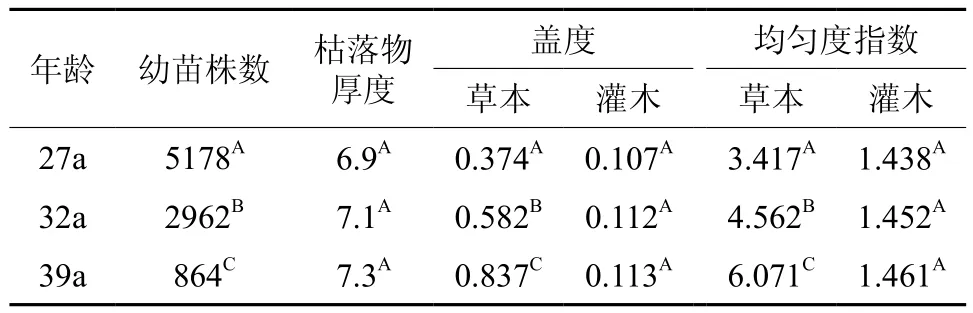
表3 林下植被与更新Table 3 Understory vegetation and seedlings
4 讨 论
森林空间结构分析只是掌握森林空间结构特征的手段,与应用联系起来才具有实际意义[29],正如Pommerening[30]指出,正确理解森林空间结构是实现森林可持续经营的关键。森林更新是森林的重建过程,包括种子的生产扩散,以及幼苗的转化与生长等多个环节,更新的每个环节都不同程度地受环境的影响[31],森林环境(光照、灌木杂草、凋落物等)常常决定更新幼苗发生的种类、数量和分布格局,对下一代森林的形成起着关键的作用[32-33]。在研究落叶松人工纯林及更新苗空间结构过程中,选用角尺度和大小比数作为不同林龄落叶松人工林对幼苗更新影响的空间格局参数,角尺度是用于描述苗木个体在水平地面上分布形式的空间格局参数[34],大小比数可以对林木(胸径 、地径、树高和冠幅等)的状态进行描述即优势、亚优势、中庸、劣势和绝对劣势[24]。周蔚等[35]认为可利用胸径大小比数来替代树高和冠幅大小比数来描述林木的大小分化程度,通过对不同林龄华北落叶松人工林结构与更新的研究发现,不同林龄落叶松人工林径阶分布于林龄呈正相关关系,其大小比数随着林龄增加林木分化呈递减趋势,平均角尺度分布在0.486~0.431的范围内,均趋于均匀分布,不同林龄落叶松林下更新苗大小比数在空间结构上随着林龄增加其分化趋势并不明显,更新苗平均角尺度均大于0.517且均呈聚集分布,且聚集程度与林龄呈负相关关系,不同林龄更新苗株数与苗龄的总体分布格局均呈倒J字型,这与乌吉斯古楞[28]在金沟岭林场研究过伐林更新幼苗空间结构分析结果一致。
通过对林分及更新苗趋势面分析发现不同林龄落叶松林在空间结构上随着林龄增加呈均匀分布趋势,这与本文研究林分大小比的结果相一致;更新苗聚集度指数随着林龄增加其聚集度呈下降趋势。林分结构与林下物种的相互作用是群落结构和动态的决定因子[36],在研究过程中不同林龄林分郁闭度在0.69~0.79的范围内,且已有研究表明在人工林下光环境的差异并不会影响种子萌发及幼苗更新[37-38],而与林下地被物类型密切相关,O’brien等[39]研究发现在其它影响因子固定的前提下,灌木覆盖每增加 1 %,辐射松(Pinus radiata)的更新概率将下降 18.5 %,在不同林龄落叶松人工林林分中灌木盖度及均匀度分布指数均无明显差异,因此灌木盖度及均匀度不是影响林下更新的主要原因;不同林龄林下草本植物的盖度及均匀度与林龄呈正相关关系且差异显著(P<0.05),这与Zimmerman[40]研究得出的草本植物的存在极大地抑制了木本植物的建立和生长的结果相一致,研究中不同林龄枯落物厚度虽无明显差异,但枯落物层会对人工林更新产生不利影响[41],因此在研究区探索枯落物及土壤对华北落叶松人工林天然更新的影响有待进一步研究,在对华北落叶松人工林进行抚育间伐的过程中应结合清理林下草本及灌木,为落叶松人工林天然更新创造良好的生存条件。
[1]Pommerening A.Approaches to quantifying forest structures[J].Forestry.2002,75(3):305-324.
[2]惠刚盈,Gadow K V, 胡艳波,等.林木分布格局类型的角尺度均值分析方法[J].生态学报,2004,24(6):1225-1229.
[3]董灵波,刘兆刚,马 妍,等.天然林林分空间结构综合指数的研究[J].北京林业大学学报,2013,35(1):16-22.
[4]Christian S, Valerie L, Patricio N,et al. Spatial patterns in an old-growth Nothofagus obliqua forest in south-central Chile[J].Forestry Ecology and Mangement,2006,231:38-46.
[5]Akhavan R, Sagheb Talebi K, Zenner E K,et al. Spatial patterns in different forest development stages of an intact old-growth Oriental beech forest in the Caspian region of Iran[J].EurJ Forest Res, 2012, 131(5): 1355-1366.
[6]Lan G Y, Hu Y H, Gao M,et al. Topography related spatial distribution of dominant tree species in a tropical seasonal rain forest in China[J]. Forestry Ecology and Mangement, 2012, 261:1507-1513.
[7]惠刚盈,胡艳波.混交林树种空间隔离程度表达方式的研究[J].林业科学研究,2001,14(1):23-27.
[8]赵 阳,余新晓,信忠保,等.北京市西山地区栓皮栎林空间结构特征研究[J].水土保持通报,2011,31(4):22-27.
[9]岳永杰,余新晓,李钢铁,等.北京松山自然保护区蒙古栎林的空间结构特征[J].应用生态学报,2009,20( 8):1811-1816.
[10]马洪婧,袁发银,刘中亮,等.北亚热带栎树混交林空间结构特征分析[J].西北林学院学报,2013,28(1):151-156.
[11]Aber J D. Foliage-height profiles and succes sion in northern hardwood forest[J]. Ecology, 1979, 60:18-231.
[12]Brown M J, Parker G C. Canopy light transmittance in a chronosequence of mixed species deciduous forest[J]. Canadian Journal of Forest Research, 1994, 24(8): 1694-1703.
[13]Denslow J S, Guzman G S. Variation in stand structure, light and seedling abundance across a tropical moist forest chronos equence, Panama[J]. Journal of Vegetation Science, 2000, 11:201-212 .
[14]Clark D B, Clark D A, Rich P M,et al. Landscape-scale analysi s of forest structure and unders torylight environments in a neotropical lowland rain forest[J].Canadian Journal of Forest Research, 1996, 26(5):747-757.
[15]Nicotra A B,Chazdon R L,Lriatre V B.Spatial heterogeneity of light andwoody seedling regeneration in tropical wet forest[J].Ecology,1999,80:1908 -1926.
[16]David R L,Lawrence C B. An analysis of structure of tree seedling populations on a Lahar[J]. Landscape Ecology, 1998,13: 307-3221.
[17]金永焕,李敦求,姜好相,等.长白山区次生林恢复过程中天然更新的动态[J].南京林业大学学报: 自然科学版,2005,29(5): 65-68.
[18]袁正科,田育新.不同功能防护林类型的判别技术研究[J].林业科学 ,1998,11(3) :1-5.
[19]Palmer M W. Fractal geometry: a tool for describing spatial patterns of plant com munities[J].Vegetatio,1988,75:91-102.
[20]辛晓平,王宗礼,李向林.宜昌百里荒草山草坡群落物种分布的空间趋势分析[J].生态学报,2003,23(8):1519-1525.
[21]毛 磊,王冬梅,杨晓晖,等.樟子松幼树在不同林分结构中的空间分布及其更新分析[J].北京林业大学学报,2008, 30(6):71-77.
[22]周建云,李荣,何景峰,等.近自然经营对辽东栎林优势乔木更新的影响[J].林业科学,2013,49(8):15-20.
[23]惠刚盈,Klaus von Gadow,Matthias Albert.一个新的林分空间结构参数-大小比数[J].林业科学研究,1999,12(1): 1-6.
[24]惠刚盈.角尺度——一个描述林木个体分布格局的结构参数 [J]. 林业科学 ,1999,35(1): 37-42.
[25]耿协鹏,杨传勇,胡 鹏.基于地图代数距离变换的空间实体分布的聚集度分析[J].测绘科学,2006,31(2):86-87.
[26]Legendra P. Quantitative methods and biogeographic analysis .In: Garbary D J,South R R,eds. Evolutionary biogeography of marinealgae of the North Atlantic. 1989,9-34.
[27]刘志龙,虞木奎,马 跃,等.不同种源麻栎种子和苗木性状地理变异趋势面分析[J].生态学报,2011,31(22):6796-6804.
[28]乌吉斯古楞,王俊峰,郑小贤,等.金沟岭林场过伐林更新幼苗空间结构分析[J].中南林业科技大学学报,2009,29(4):21-25.
[29]汤孟平.森林空间结构研究现状与发展趋势[J].林业科学,2010, 46(1):117-122.
[30]Pommerening A. Evaluating structural indices by reversing forest structural analysis. Forestry Ecology and Mangement, 2006,224(3): 3266-277.
[31]汤景明,翟明普.影响天然林树种更新因素的研究进展[J].福建林学院学报,2005,25( 5):379-383.
[32]韩有志,王政权.天然次生林中水曲柳种子的扩散格局[J].植物生态学报,2002,26( 1):51-57.
[33]韩有志,程志枫,常 洁,等.水曲柳人工林下天然更新幼苗的空间格局[J].山西农业大学学报,2000,20( 4):335-338.
[34]黄丽霞,袁位高,江 波,等.不同经营方式下杨梅林分布空间结构比较[J].浙江林学院学报,2009,26(2):209-214.
[35]周 蔚,杨 华,亢新刚,等.长白山云冷杉原始林径阶多样性及空间结构分析[J].江西农业大学学报,2012,34(2):311-316.
[36]Taylor A H, Huang J Y, Zhou S Q. Canopy tree development and undergrowth bamboo dynamics in old_growth Abies Betula forests in southwestern China:a 12-year sudy[J].Forest Ecology and Management, 2004,200:347- 360.
[37]Hunziker U, Brang P. Microsite patterns of conifer seedling establishment and growth in a mixed stand in the Southern Alps[J]. Forest Ecology and Management, 2005,210:67-79.
[38]尹华军,程新颖,赖 挺,等.川西亚高山65年人工云杉林种子雨、种子库和幼苗定居研究[J].植物生态学报,2011,35(1):35-44.
[39]O’brien M J,O. haraK L, Erbilgin N,et al. Overstory and shrub effects on natural regeneration processes in native Pinus radiata stands[J]. Forest Ecology and Management, 2007,240:178-185 .
[40]Zimmerman JK, Pascarella J B, Aide T M. Barriers to forest regeneration in an abandoned pasture in Puerto Rico[J].Restoration Ecology, 2000,8:350-360 .
[41]胡 蓉,林 波,刘 庆.林窗与凋落物对人工云杉林早期更新的影响[J].林业科学,2011,47(6):23-29.
Stand Structure and Natural Regeneration ofLarix principis-rupprechtiiPlantation in North Mountain of Hebei
WANG Dong-zhi1, ZHANG Dong-yan2, ZHANG Zhi-dong1, MU Hong-xiang1, ZHANG Yu-zhen1
(1.College of Forestry, Agricultural University of Hebei, Baoding 071000, Hebei, China;2.College of Business, Agricultural University of Hebei, Baoding 071000, Hebei, China)
Stand structure largely determines the stability of forest and the survival of seedlings and their distribution pattern, therefore it is important to research stand structure and regeneration pattern. The stand structure and seedling distribution pattern of differentLarix principis-rupprechtiiplantations were studied using the methods of uniform angel index, neighborhood and trend surface analysis. The results showed that the tree individuals were randomly distributed in a 27-year-oldLarix principis-rupprechtiiplantation, while uniform distribution was found in 32-year-old and 39-year-oldLarix principis-rupprechtiiplantations respectively. Seedlings individuals were clumply distributed in all plantations with different stand years and the degree of aggregation was negatively correlated with stand age.The individual’s differentiation showed a decreasing trend with increasing stand age. The variation of spatial structure decreased with increasing stand age and tended to be relatively stable in old-growth plantations. The individual’s differentiation of seedling was not obvious in spatial structure with increasing stand age. The grass cover is the major determinent of the number and distribution of seedlings.The management practices, including thinning, grass and litter removal, should be conduct to promote natural regeneration inLarix principis-rupprechtiiPlantation.
plantation; parameter of spatial structure; trend surface analysis; natural regeneration
S758.5+3;S754.1
A
1673-923X(2015)10-0033-07
10.14067/j.cnki.1673-923x.2015.10.006
2014-02-10
科技部十二五科技支撑计划(2012BAD22B0304 );国家自然科学基金(31370636);河北省林业厅项目(1308451)
王冬至,博士研究生
张玉珍,副教授;E-mail:hxr1962@163.com
王冬至,张冬燕,张志东,等. 冀北山区不同林龄落叶松人工林林分结构及更新[J].中南林业科技大学学报,2015,35(10): 33-39.
[本文编校:吴 彬]
