Eff ects of total saponins of Panax notoginseng on immature neuroblasts in the adult olfactory bulb following global cerebral ischemia/reperfusion
2015-12-15XuHeFengjunDengJinwenGeXiaoxinYanAihuaPanZhiyuanLi
Xu He, Feng-jun Deng, Jin-wen Ge, Xiao-xin Yan Ai-hua Pan, Zhi-yuan Li
1 Department of Anatomy and Neurobiology, School of Basic Medical Sciences, Central South University, Changsha, Hunan Province, China
2 Department of Anatomy, Yiyang Medical College, Yiyang, Hunan Province, China
3 Department of Pharmacy, Yiyang Medical College, Yiyang, Hunan Province, China
4 Department of Integrated Traditional and Western Medicine, Hunan University of Traditional Chinese Medicine, Changsha, Hunan Province, China
Eff ects of total saponins of Panax notoginseng on immature neuroblasts in the adult olfactory bulb following global cerebral ischemia/reperfusion
Xu He1,2,#, Feng-jun Deng3,#, Jin-wen Ge4, Xiao-xin Yan1, Ai-hua Pan1,*, Zhi-yuan Li1,*
1 Department of Anatomy and Neurobiology, School of Basic Medical Sciences, Central South University, Changsha, Hunan Province, China
2 Department of Anatomy, Yiyang Medical College, Yiyang, Hunan Province, China
3 Department of Pharmacy, Yiyang Medical College, Yiyang, Hunan Province, China
4 Department of Integrated Traditional and Western Medicine, Hunan University of Traditional Chinese Medicine, Changsha, Hunan Province, China
The main active components extracted from Panax notoginseng are total saponins. They have been shown to inhibit platelet aggregation, increase cerebral blood fl ow, improve neurological behavior, decrease infarct volume and promote proliferation and diff erentiation of neural stem cells in the hippocampus and lateral ventricles. However, there is a lack of studies on whether total saponins of Panax notoginseng have potential benefi ts on immature neuroblasts in the olfactory bulb following ischemia and reperfusion. This study established a rat model of global cerebral ischemia and reperfusion using four-vessel occlusion. Rats were administered total saponins of Panax notoginseng at 75 mg/kg intraperitoneally 30 minutes after ischemia then once a day, for either 7 or 14 days. Total saponins of Panax notoginseng enhanced the number of doublecortin (DCX)+neural progenitor cells and increased co-localization of DCX with neuronal nuclei and phosphorylated cAMP response element-binding/DCX+neural progenitor cells in the olfactory bulb at 7 and 14 days post ischemia. These fi ndings indicate that following global brain ischemia/reperfusion, total saponins of Panax notoginseng promote diff erentiation of DCX+cells expressing immature neuroblasts in the olfactory bulb and the underlying mechanism is related to the activation of the signaling pathway of cyclic adenosine monophosphate response element binding protein.
nerve regeneration; total saponins of Panax notoginseng; cerebral ischemia/reperfusion; immature neurons; neurogenesis; doublecortin; olfactory bulb; neural regeneration
Funding: This study was supported by the Hunan Provincial Innovation Foundation for Postgraduate in China, No. CX2014B099 (to XH); the Science Foundation of Hunan Provincial Education Department of China, No. 11C1264 (to FJD), 13C958(to XH).
He X, Deng FJ, Ge JW, Yan XX, Pan AH, Li ZY (2015) Eff ects of total saponins of Panax notoginseng on immature neuroblasts in the adult olfactory bulb following global cerebral ischemia/reperfusion. Neural Regen Res 10(9):1450-1456.
Introduction
The subventricular zone of the lateral ventricle is a neurogenic niche because it can maintain neurogenesis throughout life (Luskin, 1993; Alvarez-Buylla and Garcı́a-Verdugo, 2002; Ponti et al., 2013). Precursors from the subventricular zone migrate toward the olfactory bulb following the rostral migratory stream pathway where they are functionally integrated into circularity networks (Winner et al., 2002; Ma et al., 2009; Nissant et al., 2009; Macklis, 2012). Doublecortin (DCX) is an endogenous microtubule-associated protein with an immature phenotype and is a good marker of neuroblasts (Francis et al., 1999; Brown et al., 2003; Couillard-Despres et al., 2005; Saaltink et al., 2012). Transient cerebral ischemia or brain trauma can stimulate endogenous neurogenesis in certain areas of the brain including the olfactory bulb, and this contributes to neuronal recovery to varying degrees (Zhang et al., 2001; Iwai et al., 2003; Choi et al., 2010; Pan et al., 2013). However, the capacity of endogenous self-repair is limited, so it is imperative to fi nd either candidate drugs or other strategies to induce and amplify endogenous adult precursors following global cerebral ischemia.
Panax notoginseng is a traditional Chinese medicine widely used in China to remove blood stasis. Total saponins of Panax notoginseng (TSPN) are the bioactive constituents of Panax notoginseng (Ng, 2006), and are responsible for inhibiting apoptosis and caspase activation (Li et al., 2009), enhancing endogenous brain-derived neurotrophic factor expression, reducing cytokines and blood-brain barrier permeability, and stimulating neural stem cell proliferation and diff erentiation in the hippocampus (Si et al., 2011). Although the olfactory bulb is an area that displays neurogenesis and is an important part of the rostral migratory stream, there are no reports on
whether TSPN could enhance neurogenesis in the olfactory bulb following global cerebral ischemia/reperfusion (I/R). This study evaluated the eff ects of TSPN on immature neuroblasts in the olfactory bulb of a rat model of transient global cerebral I/R induced by four-vessel occlusion.
Materials and Methods
Establishment of transient global cerebral I/R models
A total of 80 specifi c-pathogen-free male Sprague-Dawley rats weighing 250–290 g and aged 8 weeks were obtained from the Animal Center of Xiangya School of Medicine, Central South University, China (license No. SCXK (Xiang) 2009-0012). All animal procedures conformed to the National Institutes of Health Guide for the care and use of laboratory animals, and all procedures were approved by the Ethics Committee of Central South University, China. All efforts were made to minimize animal discomfort and reduce the number of animals used.
Four-vessel occlusion was used to induce cerebral ischemia (Pulsinelli and Brierley, 1979). Briefl y, all rats were anesthetized with 1% sodium pentobarbital diluted in distilled water (50 mg/kg, intraperitoneally) in a sterile room. The rats were positioned in Kopf stereotaxic apparatus (David KOPF Instruments, Tujunga, CA, USA) and the vertebral arteries were irreversibly occluded by electrocoagulation. A non-absorbable suture was looped around the carotid arteries. On the following day, rats were re-anesthetized and carotid arteries were disconnected carefully to avoid damage to the vagus nerve, and then occluded with micro-arterial clamps for 30 minutes. Rats lost their righting refl ex during I/R and this was achieved by de-clamping the arteries. During the surgery, rectal temperature was monitored and maintained at 37 ± 0.5°C with a rectal probe and a heat lamp was used to ensure the body temperature at 37°C. Eight rats died, so the remaining 72 animals were equally and randomly divided into the TSPN group and vehicle group.
TSPN administration
The rats in the TSPN group were administered TSPN (reagent No. M004538, high-performance liquid chromatography ≥ 98%, Chengdu Maikaxi Chemical Co., Ltd., Chengdu, China) intraperitoneally 30 minutes after brain ischemia. The dose of TSPN was 75 mg/kg suspended in 0.9% sodium salt 10 g/L, once per day for 1, 7 and 14 days after reperfusion. Rats in the vehicle group were treated with an equal volume of sodium salt, one injection per day until the rats were sacrifi ced at either 1, 7 or 14 days post ischemia.
Tissue preparation
Rats in both groups were intraperitoneally anesthetized with a high dose of sodium pentobarbital (400 mg/kg) and then perfused transcardially with saline followed by 4% paraformaldehyde. The brains were post-fi xed at 4°C overnight, and were treated in gradual concentrations of sucrose (15% and 30%) until they sank. Coronal sections were prepared across the olfactory bulb (Paxinos and Watson, 2005) in a cryostat (Thermo Shandon Limited, UK). Twelve sets of 30-μm sections were cut and collected for western blot assay. For double immunofl uorescence, 12 sets of 8-μm sections were also collected by thaw-mounting on positively charged microslides.
Immunohistochemistry and immunofl uorescent labeling Immunohistochemistry was carried out using the avidin-biotin complex method. Sections were treated with 3% H2O2for 30 minutes at 24 ± 1°C, and pre-incubated for 1 hour, followed by incubation with goat anti-DCX (1:2,000; Santa Cruz Biotechnology, Dallas, TX, USA) overnight at 4°C. Sections were further reacted with rabbit anti-goat IgGs (Boster, Wuhan, Hubei Province, China) at 1:400 for 2 hours at 24 ± 1°C and subsequently with avidin-biotin complex reagents for an additional 2 hours. Immunoreactivity was visualized using 0.003% hydrogen peroxide and 0.05% 3,3′-diaminobenzidine.
Immunofl uorescent labeling was carried out as follows: sections were immersed in PBS containing 5% donkey serum and 0.1% Triton X-100 for 1 hour, treated with goat anti-DCX (1:2,000), mouse anti-neuronal nuclei (NeuN) (1:4,000; Merck Millipore, Darmstadt, Germany), rabbit anti-phosphorylated cyclic adenosine monophosphate response element binding protein (p-CREB) (1:2,000; Cell Signaling, Boston, MA, USA) overnight at 4°C, followed by a 2-hour reaction with Alexa-Fluor 488 and Alexa-Fluor 594-donkey anti-goat, anti-mouse, anti-rabbit IgGs (1:200, Invitrogen, Carlsbad, CA, USA). Sections were counterstained with bisbenzimide (1:50,000; Hoechst 33342).
Immunostained sections were examined and imaged on a microscope equipped with a digital camera and imaging system (BX40, Olympus, Tokyo, Japan). The sections of the olfactory bulb used for comparison were on the same horizontal plane. Because the DCX+cells in the olfactory bulb were in a cluster, the mean optical density of images was captured using an identical photographic setting and compared using NIH Image J (NIH, MD, USA). Images at 40× magnification were taken for morphological analyses of positive cells. Co-localization of cells in the olfactory bulb was calculated in a set of sections. In each section, fi ve fi elds were captured with a 20× objective lens. The total DCX+cells and co-localized cells were added together for each section, brain, and group. The formula for co-localization rate was: co-localized DCX+cells/total DCX+cells × 100%.
Western blot assay
The olfactory bulb was snap-frozen in liquid nitrogen. Frozen tissues were homogenized in a cocktail buff er, including phosphatase inhibitors and protease inhibitors (Roche Applied Science, Mannheim, Germany). The samples were then centrifuged at 1,200 × g for 15 minutes at 4°C. Bicinchoninic acid assay was used to measure the protein concentrations from supernatants. A total of 20 μg protein was loaded in each lane and separated by 12% Bis-Tris sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Subsequently, the blotted proteins were transferred to nitrocellulose membrane. The membrane was blocked using 10% nonfat milk for 2 hours at 24 ± 1°C, then incubated overnight with goat anti-DCX
(1:1,000, Santa Cruz Biotechnology) and mouse anti-GAPDH (1:10,000; Boster). The membrane was washed and incubated with horseradish peroxidase-conjugate rabbit anti-goat or anti-mouse (1:5,000; Merck Millipore, Merck KGaA, Germany) for 2 hours at 24 ± 1°C, followed by exposure to photographic fi lms for 1 to 5 minutes. Western blot band intensity (integrated optical density) was measured using NIH Image J. GAPDH was used as an internal reference.

Figure 1 Eff ects of TSPN on DCX immunoreactivity in the olfactory bulb of adult rats following global cerebral ischemia/reperfusion.

Figure 2 Representative western blot assay (A) and quantifi cation of DCX protein levels (B) in the olfactory bulb after global cerebral ischemia/reperfusion.
Statistical analysis
All data are expressed as the mean ± SD, and were analyzed using Prism GraphPad5.0 (GraphPad Software Inc., La Jolla, CA, USA). Comparisons were conducted using one-way analysis of variance followed by Bonferroni post hoc tests. A value of P ≤ 0.05 was considered statistically signifi cant.
Results
TSPN up-regulated the number of DCX+cells in the olfactory bulb following global cerebral I/R

Figure 3 Double immunofl uorescence staining for DCX and NeuN (A) and quantitative data (B) in the olfactory bulb of adult rats at 14 days post ischemia.
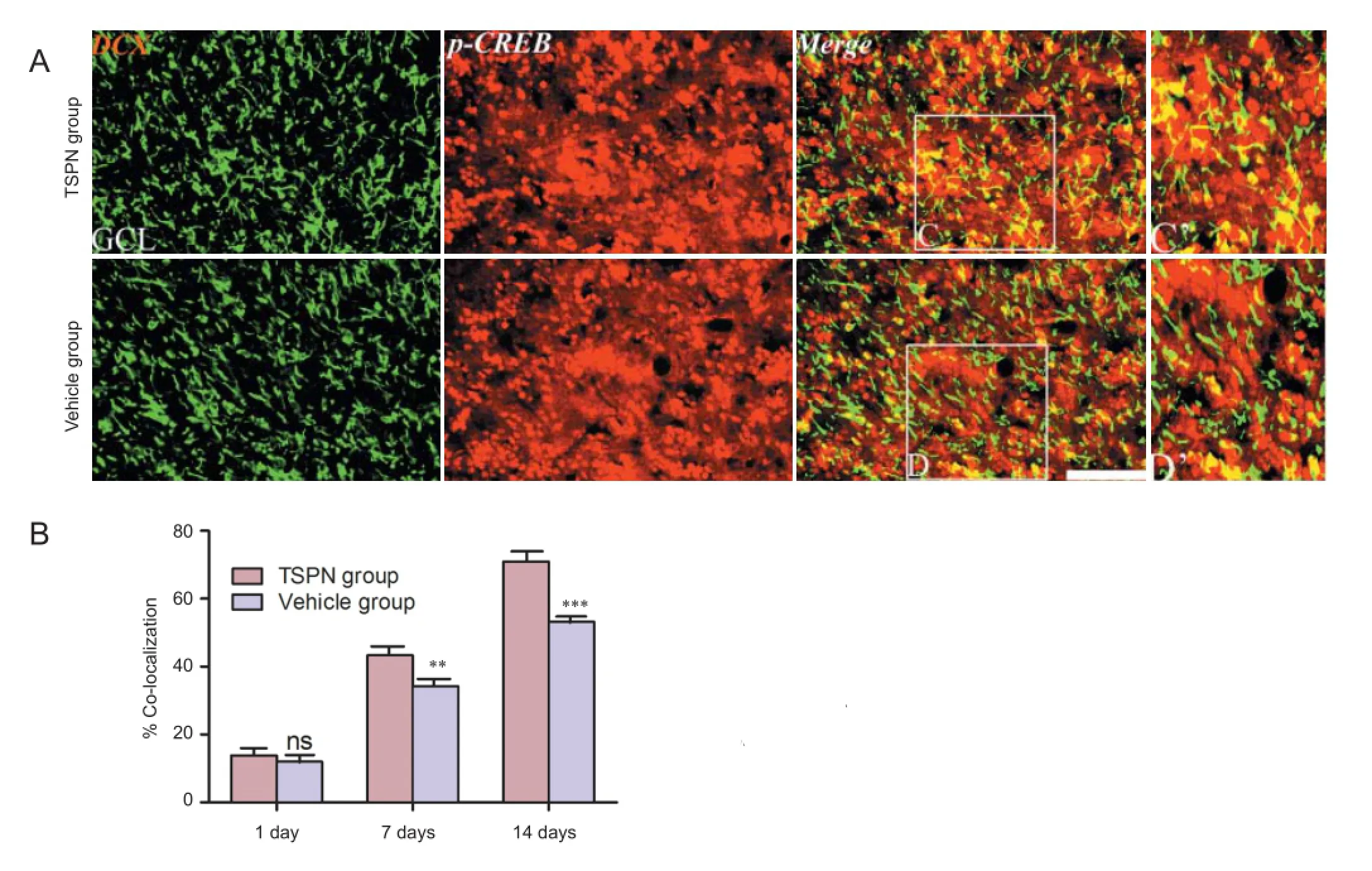
Figure 4 Double immunofl uorescence staining for DCX and p-CREB (A) and quantitative data (B) in the olfactory bulb of adult rats at 14 days post ischemia.
DCX has been used as a marker for immature neurons, and DCX+cells populate in the piriform cortex and olfactory bulb of rabbits, cats, rats, guinea pigs and non-human primates
(Xiong et al., 2008; Cai et al., 2009; Zhang et al., 2009; Klempin et al., 2011; He et al., 2014; Zeng et al., 2014). In the present study, global cerebral I/R aff ected DCX labeling, with varied shapes and sizes of cells detectable mainly around the granule cell layer, internal plexiform layer, and mitral cell layer. Dendrite-like processes chained in a cluster in the granule cell layer (Figure 1A3, B3). There was no signifi cant diff erence in the mean optical density of DCX in the granule cell layer in vehicle or TSPN groups 1 day post ischemia (P> 0.05). In comparison with the vehicle group, TSPN signifi cantly increased the mean optical density of DCX+cells in the granule cell layer at 7 and 14 days post ischemia (day 7: P < 0.05; day 14: P < 0.001; Figure 1C). Furthermore, the mean optical density of DCX+cells was signifi cantly higher in the TSPN group than in the vehicle group in the internal plexiform layer and mitral cell layer (P < 0.0001, F = 16.8, degrees of freedom (df) = 5,24; P < 0.0001, F = 10.7, df = 5,24). However, there was no signifi cant diff erence in the internal plexiform layer and mitral cell layer between the TSPN and vehicle groups 1 day post ischemia (P > 0.05). At 7 and 14 days after ischemia, the mean optical density of DCX+cells in the internal plexiform layer was signifi cantly higher in the TSPN group than in the vehicle group (day 7: P< 0.05; day 14: P < 0.01). The mean optical density of DCX+cells in the mitral cell layer was signifi cantly higher in the TSPN group than in the vehicle group at 14 days post ischemia (P < 0.05; Figure 1D and E).
TSPN elevated DCX protein levels in the olfactory bulb following global cerebral I/R
DCX protein levels in the olfactory bulb formation were determined by western blot assay. Immunoblotted DCX signal increased in the TSPN group compared with the vehicle group at 7 and 14 days post ischemia (Figure 2A). The mean optical density of DCX protein level at 7 and 14 days post ischemia was signifi cantly higher in the TSPN group than in the vehicle group (day 7: P < 0.001; day 14: P < 0.001), but not at 1 day post ischemia (P > 0.05; Figure 2B).
TSPN enhanced co-localization of DCX with NeuN in the olfactory bulb following global cerebral I/R
There were a higher number of DCX+cells in the granule cell layer than in any other layer between the vehicle and TSPN groups (Figure 1). We therefore focused on the granule cell layer. DCX+cells expressing NeuN were detected in the olfactory bulb at all time points post ischemia (Figure 3A), and the ratios of DCX+cells co-expressing NeuN to the total DCX-labeled cells were higher in the TSPN group than in the vehicle group at both 7 and 14 days post ischemia (day 7: P < 0.01; day 14: P < 0.001), but not at 1 day (P > 0.05; Figure 3B).
TSPN induced immature neuroblasts in the olfactory bulb through phosphorylation of cyclic adenosine
monophosphate response element binding protein (CREB) following global cerebral I/R
To further understand the underlying mechanism by which TSPN regulated immature neurons in the olfactory bulb, we tested the activation of p-CREB in the olfactory bulb following global cerebral ischemia using immunohistochemistry. A number of p-CREB/DCX double-labeled cells were found in both groups at various time points (Figure 4A). Moreover, the percentage of p-CREB/DCX double-labeled cells signifi -cantly increased in the TSPN group compared with the vehicle group at 7 days (P < 0.01) and at 14 days (P < 0.001), but not at 1 day post ischemia (P > 0.05; Figure 4B).
Discussion
We examined TSPN eff ects on immature neuroblasts in an animal model of global cerebral I/R. TSPN evidently enhanced DCX-expressing immature neurons in the olfactory bulb. Moreover, TSPN noticeably increased DCX/NeuN co-expression and p-CREB/DCX+cells, which suggests that TSPN can promote neurochemical diff erentiation of immature neural cells in the olfactory bulb following global ischemia by up-regulating the p-CREB signaling pathway.
Neurogenesis is a process whereby precursor cells divide into immature neurons and then diff erentiate into neurons with a mature phenotype (Lazarini et al., 2014). Immature neurons expressing DCX are of great signifi cance to brain plasticity and development (Nacher et al., 2001; Bloch et al., 2011) and are involved in cytoskeletal changes (Kutsuna et al., 2014) and structural plasticity, and are thought to occur at synaptic and neuritic levels under physiological conditions (Luo, 2002; He et al., 2014). Neurogenesis can be accelerated after brain ischemia in the adult rat, monkey, human cerebrum including the olfactory bulb (Jin et al., 2001b; Tonchev et al., 2003; Gould, 2007; Kutsuna et al., 2014). In our study, the increased numerical density of immature DCX-expressing neurons indicates an increased population of newborn immature neurons and dendritic processes. To further confi rm the immunohistochemistry result, western blots were carried out to detect DCX protein levels. Densitometry was consistent with the immunofl uorescence, suggesting that TSPN increased the population of immature neurons in the olfactory bulb following global cerebral I/R.
It is well known that when newly generated precursor cells in the adult brain begin developing and maturing biochemically and morphologically (Brown et al., 2003), newly generated neurons in the olfactory bulb are crucial for olfactory function (Gheusi et al., 2000; Lledo and Saghatelyan, 2005; Kermen et al., 2010). NeuN is a mature neuronal marker (Mullen et al., 1992; Sarnat et al., 1998) and is both apparent and increased during hippocampal neurogenesis. Therefore, the extent of DCX/NeuN co-expression may implicate the neurochemical maturity of immature olfactory DCX-expressing neurons. Results from this study showed a signifi cant increase in NeuN co-expression among the DCX+neurons in the olfactory bulb of global cerebral ischemia rats following intraperitoneally injecting TSPN at 7 and 14 days. These fi ndings suggest that TSPN could enhance and drive the neurochemical diff erentiation of immature neurons in the olfactory bulb toward the phenotype of maturation.
CREB is an important nuclear transcription factor, and
experiments in vivo and vitro demonstrated that CREB signaling pathways play a vital role in the neurogenesis and survival of immature neuroblasts in the olfactory bulb (Giachino et al., 2005), subventricular zone (Gampe et al., 2011; Herold et al., 2011) and hippocampus (Jagasia et al., 2009; Kim et al., 2010). Activated CREB is involved in many biological functions such as enhancement of neuronal regeneration, synapse formation and spatial learning (Walton et al., 1996; O’Connell et al., 2000; Rajan et al., 2015). In transient ischemic adult rodents, high levels of p-CREB existed (Walton et al., 1996; Tanaka et al., 2000; Jin et al., 2001a) in immature DCX-expressing neurons in the dentate gyrus (Nakagawa et al., 2002). Cell counting showed that there were a large number of p-CREB/DCX+neural progenitor cells in the olfactory bulb of the two groups at diff erent time points, which suggested engagement of the CREB signaling pathway in adult neurogenesis and recruitment of immature DCX-expressing neurons for brain repair following global cerebral ischemia. Furthermore, TSPN apparently increased the number of the DCX+cells expressing p-CREB in the olfactory bulb of rats surviving 7 and 14 days, implying that TSPN could modulate the newborn immature neuroblasts by up-regulating the p-CREB signaling pathway.
In summary, TSPN could enhance regeneration and maturation of DCX+cells expressing immature neuroblasts in the olfactory bulb of adult rats through activating the CREB signaling pathway following global cerebral I/R.
Author contributions: ZYL and AHP designed the study. XH, FJD, and JWG implemented the experiments. XH wrote and revised the paper. XXY analyzed the data and supervised the whole research. All authors approved the fi nal version of the paper.
Confl icts of interest: None declared.
Alvarez-Buylla A, Garcı́a-Verdugo JM (2002) Neurogenesis in adult subventricular zone. J Neurosci 22:629-634.
Bloch J, Kaeser M, Sadeghi Y, Rouiller EM, Redmond DE, Brunet JF (2011) Doublecortin-positive cells in the adult primate cerebral cortex and possible role in brain plasticity and development. J Comp Neurol 519:775-789.
Brown JP, Couillard-Després S, Cooper-Kuhn CM, Winkler J, Aigner L, Kuhn HG (2003) Transient expression of doublecortin during adult neurogenesis. J Comp Neurol 467:1-10.
Cai Y, Xiong K, Chu Y, Luo DW, Luo XG, Yuan XY, Struble RG, Clough RW, Spencer DD, Williamson A, Kordower JH, Patrylo PR, Yan XX (2009) Doublecortin expression in adult cat and primate cerebral cortex relates to immature neurons that develop into GABAergic subgroups. Exp Neurol 216:342-356.
Choi JH, Yoo KY, Park OK, Lee CH, Kim SK, Hwang IK, Lee YL, Shin HC, Won MH (2010) Relation among neuronal death, cell proliferation and neuronal diff erentiation in the gerbil main olfactory bulb after transient cerebral ischemia. Cell Mol Neurobiol 30:929-938.
Couillard-Despres S, Winner B, Schaubeck S, Aigner R, Vroemen M, Weidner N, Bogdahn U, Winkler J, Kuhn HG, Aigner L (2005) Doublecortin expression levels in adult brain refl ect neurogenesis. Eur J Neurosci 21:1-14.
Francis F, Koulakoff A, Boucher D, Chafey P, Schaar B, Vinet MC, Friocourt G, McDonnell N, Reiner O, Kahn A, McConnell SK, Berwald-Netter Y, Denoulet P, Chelly J (1999) Doublecortin is a developmentally regulated, microtubule-associated protein expressed in migrating and diff erentiating neurons. Neuron 23:247-256.
Gampe K, Brill MS, Momma S, Götz M, Zimmermann H (2011) EGF induces CREB and ERK activation at the wall of the mouse lateral ventricles. Brain Res 1376:31-41.
Gheusi G, Cremer H, McLean H, Chazal G, Vincent JD, Lledo PM (2000) Importance of newly generated neurons in the adult olfactory bulb for odor discrimination. Proc Natl Acad Sci U S A 97:1823-1828.
Giachino C, De Marchis S, Giampietro C, Parlato R, Perroteau I, Schütz G, Fasolo A, Peretto P (2005) cAMP response element-binding protein regulates diff erentiation and survival of newborn neurons in the olfactory bulb. J Neurosci 25:10105-10118.
Gould E (2007) How widespread is adult neurogenesis in mammals? Nat Rev Neurosci 8:481-488.
He X, Zhang XM, Wu J, Fu J, Mou L, Lu DH, Cai Y, Luo XG, Pan A, Yan XX (2014) Olfactory experience modulates immature neuron development in postnatal and adult guinea pig piriform cortex. Neuroscience 259:101-112.
Herold S, Jagasia R, Merz K, Wassmer K, Lie DC (2011) CREB signalling regulates early survival, neuronal gene expression and morphological development in adult subventricular zone neurogenesis. Mol Cell Neurosci 46:79-88.
Iwai M, Sato K, Kamada H, Omori N, Nagano, Shoji M, Abe K (2003) Temporal profi le of stem cell division, migration, and diff erentiation from subventricular zone to olfactory bulb after transient forebrain ischemia in gerbils. J Cereb Blood Flow Metab 23:331-341.
Jagasia R, Steib K, Englberger E, Herold S, Faus-Kessler T, Saxe M, Gage FH, Song H, Lie DC (2009) GABA-CREB signalling regulates maturation and survival of newly generated neurons in the adult hippocampus. J Neurosci 29:7966-7977.
Jin K, Mao X, Simon R, Greenberg D (2001a) Cyclic AMP response element binding protein (CREB) and CREB binding protein (CBP) in global cerebral ischemia. J Mol Neurosci 16:49-56.
Jin K, Minami M, Lan JQ, Mao XO, Batteur S, Simon RP, Greenberg DA (2001b) Neurogenesis in dentate subgranular zone and rostral subventricular zone after focal cerebral ischemia in the rat. Proc Natl Acad Sci U S A 98:4710-4715.
Kermen F, Sultan S, Sacquet J, Mandairon N, Didier A (2010) Consolidation of an olfactory memory trace in the olfactory bulb is required for learning-induced survival of adult-born neurons and long-term memory. PLoS One 5:e12118.
Kim JS, Yang M, Cho J, Kim SH, Kim JC, Shin T, Moon C (2010) Promotion of cAMP responsive element-binding protein activity ameliorates radiation-induced suppression of hippocampal neurogenesis in adult mice. Toxicol Res 26:177-183.
Klempin F, Kronenberg G, Cheung G, Kettenmann H, Kempermann G (2011) Properties of doublecortin-(DCX)-expressing cells in the piriform cortex compared to the neurogenic dentate gyrus of adult mice. PLoS One 6:e25760.
Kutsuna N, Yamashita A, Eriguchi T, Oshima H, Suma T, Sakatani K, Yamamoto T, Yoshino A, Katayama Y (2014) Acute stress exposure preceding transient global brain ischemia exacerbates the decrease in cortical remodeling potential in the rat retrosplenial cortex. Neurosci Res 78:65-71.
Lazarini F, Gabellec MM, Moigneu C, de Chaumont F, Olivo-Marin JC, Lledo PM (2014) Adult neurogenesis restores dopaminergic neuronal loss in the olfactory bulb. J Neurosci 34:14430-14442.
Li H, Deng CQ, Chen BY, Zhang SP, Liang Y, Luo XG (2009) Total saponins of Panax Notoginseng modulate the expression of caspases and attenuate apoptosis in rats following focal cerebral ischemia-reperfusion. J Ethnopharmacol 121:412-418.
Lledo P-M, Saghatelyan A (2005) Integrating new neurons into the adult olfactory bulb: joining the network, life-death decisions, and the eff ects of sensory experience. Trends Neurosci 28:248-254.
Luo L (2002) Actin cytoskeleton regulation in neuronal morphogenesis and structural plasticity. Annu Rev Cell Dev Biol 18:601-635.
Luskin MB (1993) Restricted proliferation and migration of postnatally generated neurons derived from the forebrain subventricular zone. Neuron 11:173-189.
Ma DK, Kim WR, Ming Gl, Song H (2009) Activity-dependent extrinsic regulation of adult olfactory bulb and hippocampal neurogenesis. Ann N Y Acad Sci 1170:664-673.
Macklis Jeff rey D (2012) Human adult olfactory bulb neurogenesis? Novelty is the best policy. Neuron 74:595-596.
Mullen RJ, Buck CR, Smith AM (1992) NeuN, a neuronal specifi c nuclear protein in vertebrates. Development 116:201-211.
Nacher J, Crespo C, McEwen BS (2001) Doublecortin expression in the adult rat telencephalon. Eur J Neurosci 14:629-644.
Nakagawa S, Kim JE, Lee R, Chen J, Fujioka T, Malberg J, Tsuji S, Duman RS (2002) Localization of phosphorylated cAMP response element-binding protein in immature neurons of adult hippocampus. J Neurosci 22:9868-9876.
Ng TB (2006) Pharmacological activity of sanchi ginseng (Panax notoginseng). J Pharm Pharmacol 58:1007-1019.
Nissant A, Bardy C, Katagiri H, Murray K, Lledo PM (2009) Adult neurogenesis promotes synaptic plasticity in the olfactory bulb. Nat Neurosci 12:728-730.
O’Connell C, Gallagher HC, O’Malley A, Bourke M, Regan CM (2000) CREB phosphorylation coincides with transient synapse formation in the rat hippocampal dentate gyrus following avoidance learning. Neural Plast 7:279-289.
Pan A, Li M, Gao JY, Xue ZQ, Li Z, Yuan XY, Luo DW, Luo XG, Yan XX (2013) Experimental epidural hematoma causes cerebral infarction and activates neocortical glial and neuronal genesis in adult guinea pigs. J Neurosci Res 91:249-261.
Paxinos G, Watson C (2005) The Rat Brain in Stereotaxic Coordinates. London: Academic Press.
Ponti G, Obernier K, Guinto C, Jose L, Bonfanti L, Alvarez-Buylla A (2013) Cell cycle and lineage progression of neural progenitors in the ventricular-subventricular zones of adult mice. Proc Natl Acad Sci U S A 110:E1045-1054.
Pulsinelli WA, Brierley JB (1979) A new model of bilateral hemispheric ischemia in the unanesthetized rat. Stroke 10:267-272.
Rajan K, Thangaleela S, Balasundaram C (2015) Spatial learning associated with stimulus response in goldfi sh Carassius auratus: relationship to activation of CREB signalling. Fish Physiol Biochem 41:685-694.
Saaltink D-J, Håvik B, Verissimo CS, Lucassen PJ, Vreugdenhil E (2012) Doublecortin and doublecortin-like are expressed in overlapping and non-overlapping neuronal cell population: implications for neurogenesis. J Comp Neurol 520:2805-2823.
Sarnat HB, Nochlin D, Born DE (1998) Neuronal nuclear antigen (NeuN): a marker of neuronal maturation in the early human fetal nervous system1. Brain Dev 20:88-94.
Si YC, Zhang JP, Xie CE, Zhang LJ, Jiang XN (2011) Effects of Panax notoginseng saponins on proliferation and diff erentiation of rat hippocampal neural stem cells. Am J Chin Med 39:999-1013.
Tanaka K, Nogawa S, Nagata E, Ito D, Suzuki S, Dembo T, Kosakai A, Fukuuchi Y (2000) Persistent CREB phosphorylation with protection of hippocampal CA1 pyramidal neurons following temporary occlusion of the middle cerebral artery in the rat. Exp Neurol 161:462-471.
Tonchev AB, Yamashima T, Zhao L, Okano H (2003) Diff erential proliferative response in the postischemic hippocampus, temporal cortex, and olfactory bulb of young adult macaque monkeys. Glia 42:209-224.
Walton M, Sirimanne E, Williams C, Gluckman P, Dragunow M (1996) The role of the cyclic AMP-responsive element binding protein (CREB) in hypoxic-ischemic brain damage and repair. Brain Res Mol Brain Res 43:21-29.
Winner B, Cooper-Kuhn CM, Aigner R, Winkler J, Kuhn HG (2002) Long-term survival and cell death of newly generated neurons in the adult rat olfactory bulb. Eur J Neurosci 16:1681-1689.
Xiong K, Luo DW, Patrylo PR, Luo XG, Struble RG, Clough RW, Yan XX (2008) Doublecortin-expressing cells are present in layer II across the adult guinea pig cerebral cortex: partial colocalization with mature interneuron markers. Exp Neurol 211:271-282.
Zeng JJ, He XH, Yan XX, Luo XG, Pan AH (2014) Unilateral olfactory functional deprivation model in the left peripheral nostrils by electric cautery injury: olfactory bulb neurogenesis and transformation. Zhongguo Zuzhi Gongcheng Yanjiu 18:231-238.
Zhang RL, Zhang ZG, Zhang L, Chopp M (2001) Proliferation and diff erentiation of progenitor cells in the cortex and the subventricular zone in the adult rat after focal cerebral ischemia. Neuroscience 105:33-41.
Zhang XM, Cai Y, Chu Y, Chen EY, Feng JC, Luo XG, Xiong K, Struble RG, Clough RW, Patrylo PR, Kordower JH, Yan XX (2009) Doublecortin-expressing cells persist in the associative cerebral cortex and amygdala in aged nonhuman primates. Front Neuroanat 3:17.
Copyedited by Paul P, Park M, Yu J, Qiu Y, Li CH, Song LP, Zhao M
*Correspondence to:
Ai-hua Pan, Ph.D. or Zhi-yuan Li, Ph.D., panaihua@csu.edu.cn or
li_zhiyuan@gibh.ac.cn.
# These authors contributed equally to this work.
orcid:
0000-0003-2068-8028 (Ai-hua Pan)
10.4103/1673-5374.165514
http://www.nrronline.org/
Accepted: 2015-08-11
杂志排行

中国神经再生研究(英文版)的其它文章
- PTEN inhibition and axon regeneration and neural repair
- Neural correlates of the Heidelberg Music Therapy: indicators for the regeneration of auditory cortex in tinnitus patients?
- The choline pathway as a strategy to promote central nervous system (CNS) remyelination
- Enhancing endogenous stem cells in the newborn via delayed umbilical cord clamping
- Elastic modulus aff ects the growth and diff erentiation of neural stem cells
- Non-steroidal anti-infl ammatory drugs (NSAIDs) and neuroprotection in the elderly: a view from the mitochondria