荒漠竹节虫的形态及生物学特性研究
2015-12-03陈新燕马东建范兆田
陈新燕,史 雯,马东建,范兆田,吴 卫*
(1.新疆大学生命科学与技术学院,乌鲁木齐 830046;2.新疆大学理化测试中心,乌鲁木齐 830046)
竹节虫目昆虫因其超高的拟态而广为人知,目前有记录的3 科500 属3000种,均为植食性物种,多分布于炎热潮湿的热带、亚热带地区,鲜少在地处温带内陆干旱区的中国新疆鲜有发现,目前发现该种为唯一一种在中国新疆的干旱、半干旱荒漠中分布的竹节虫目昆虫;其分布区域昼夜温差大,夏季极其干旱、炎热而冬季则寒冷而漫长,因此获名荒漠竹节虫(黄仁鑫等,2005)。国外对该种昆虫仅有少量报道(БЕЙ-БИЕНКО,1964;Hennemann et al.,2008),在生物学特性方面尚无文献记载。本文作者在2006年4月至2013年10月对荒漠昆虫的野外考察期间,观察并采集了一部分在新疆干旱荒漠环境中生存的荒漠竹节虫,并分别于2006年5月至2008年12月,2013年4月至12月在室内进行人工饲养,对其形态学和生物学特性有了进一步的了解,现记述如下。
1 材料与方法
1.1 研究对象
荒漠竹节虫隶属于 Phasmatodea 目Diapheromeridae 科Sceptrophasma 属(Brock & Seow-Choen,2000),实验材料为2006年6月至2013年10月期间,在乌鲁木齐市郊及准噶尔盆地荒漠南缘采集的所有成体、卵和幼体。
1.2 调查地点
1.2.1 雅玛里克山
地理位置为 N43° 45' 44″-43° 47' 25″,E87°33' 16″-87° 34'56″,位于乌鲁木齐市西侧,约呈南北走向,平均海拔800 m,最高点青年峰海拔1391 m,山体地表面积约40 km2,长度约16 km。地质结构为油页岩、片麻岩构成;植被以荒漠植被为主,植物群落构成较简单,春秋季较湿润,夏季干旱,冬季漫长而寒冷。1983-1996年统计表明,年均温7.5℃,极端最高温37.7℃,极端最低温度-19.1℃,年降水量为266.9 mm,年蒸发量2731 mm,是总降水量的10.2 倍;平均相对湿度58%,属中温带荒漠气候。由于近年来的荒山绿化和封山育林,使荒山植被除了荒漠外,还有荒漠草原、真草原、草甸、沼泽的出现(叶卫英,2005)。
1.2.2 准噶尔盆地南缘
本地区代表着欧亚大陆腹地、温带内陆荒漠区。地带性植被为典型的荒漠植被,如梭梭Haloxylon ammodendron、沙拐枣 Calligonum mongolicum、绢蒿Seriphidium korovinii 等;气候类型为典型的大陆性干旱气候,年均温6.6℃,最高气温42.6℃,最低气温-41.6℃,年降水量164 mm,年蒸发量2000 mm 左右。本研究选择102 团荒漠(E87°81'09″N44°36'49″)和阜康荒漠生态试验站(E87°87'92″ N44°37'71″)2个区域进行调查。生境较为相似,但后者区域内植被保护较好,且有人工植被及试验地分布,是研究的理想选地。
1.3 研究方法
1.3.1 野外观察
分别在上述调查地点对荒漠竹节虫成虫及幼虫的生态、行为、食性等进行观察,于卵期在其栖息地的寄主植物上寻找自然产出的卵粒,标记并定期观察,以确定其孵化时间。
1.3.2 室内饲养
除了野外观察,还在室内进行人工饲养,方法如下:人工移植不同栖息地的寄主植物绢蒿至室内,与当地采集到的4 龄或5 龄若虫一同罩在自制的纱笼中,历经蜕皮、羽化、产卵、孵化等阶段;五龄若虫单独饲养,以观察其繁殖特性,用Olympus μ-mini D、尼康D50 和Olympus E-410等数码相机进行户外及室内影像记录;用尼康SMZ-800 解剖显微镜和Lnoe-1403VT 扫描电子显微镜进行成体及卵的显微结构观察;使用HTC-1电子温湿度计测量温、湿度数据,标本保存于新疆大学生命科学与技术学院。
2 结果与分析
2.1 成虫形态特征
身体细长、纤弱,树枝状,灰土色,表面光裸,完全无翅(图1 A)。
头部呈卵圆形,触角粗短,明显分为16 节,其上有稀疏的感觉毛,长度不超过前足腿节的1/3;柄节背腹方向扁平,长而宽大,背面有一纵向脊;梗节短,宽为长的1.2 倍;鞭节14 节,略扁平,第一鞭小节的长度是宽度的2.6 倍,第三和第五节鞭节的长和宽几乎相等,末节端部圆,长是宽的2.2 倍;其余鞭节宽大于长。复眼椭圆而突出,复眼间具成对纵行的结节,无单眼。唇基前缘平直,额略凹陷。颞边缘隆起,长度是复眼纵向直径的5 倍;后头平,向后变窄(图1 B)。胸部细长较扁,无翅,前胸为前端略窄的梯形,比头部稍窄,中胸是胸部最长的一节,长为宽的6 倍,中胸、后胸背板中央有一纵棱,与两侧边的脊平行,前胸、中胸和后胸比例为1.0∶6.2∶4.6。足细长,前足股节基部弯曲,稍扁。中、后足胫节下方中棱达胫节端部。腿节∶胫节∶跗节长度比例为:前足:4.1∶4.3∶1.0;中足:4.0∶4.3∶1.0;后足:4.4∶5.0∶1.0。
腹部扁平,明显分为10 节,第1 节较短,第2-7 节逐渐延长,第4-5 节最宽。雌体腹部第7节腹板末端具尖突,腹瓣光滑,尖矛形,不超过臀节基部;臀节有时凹陷。肛上板有3 条纵棱,两侧的不甚明显,第8、9 节的背板中棱较平坦;尾须不分节(图1:C,D,E)。
体长:6.2-7.6 mm(雌体)。
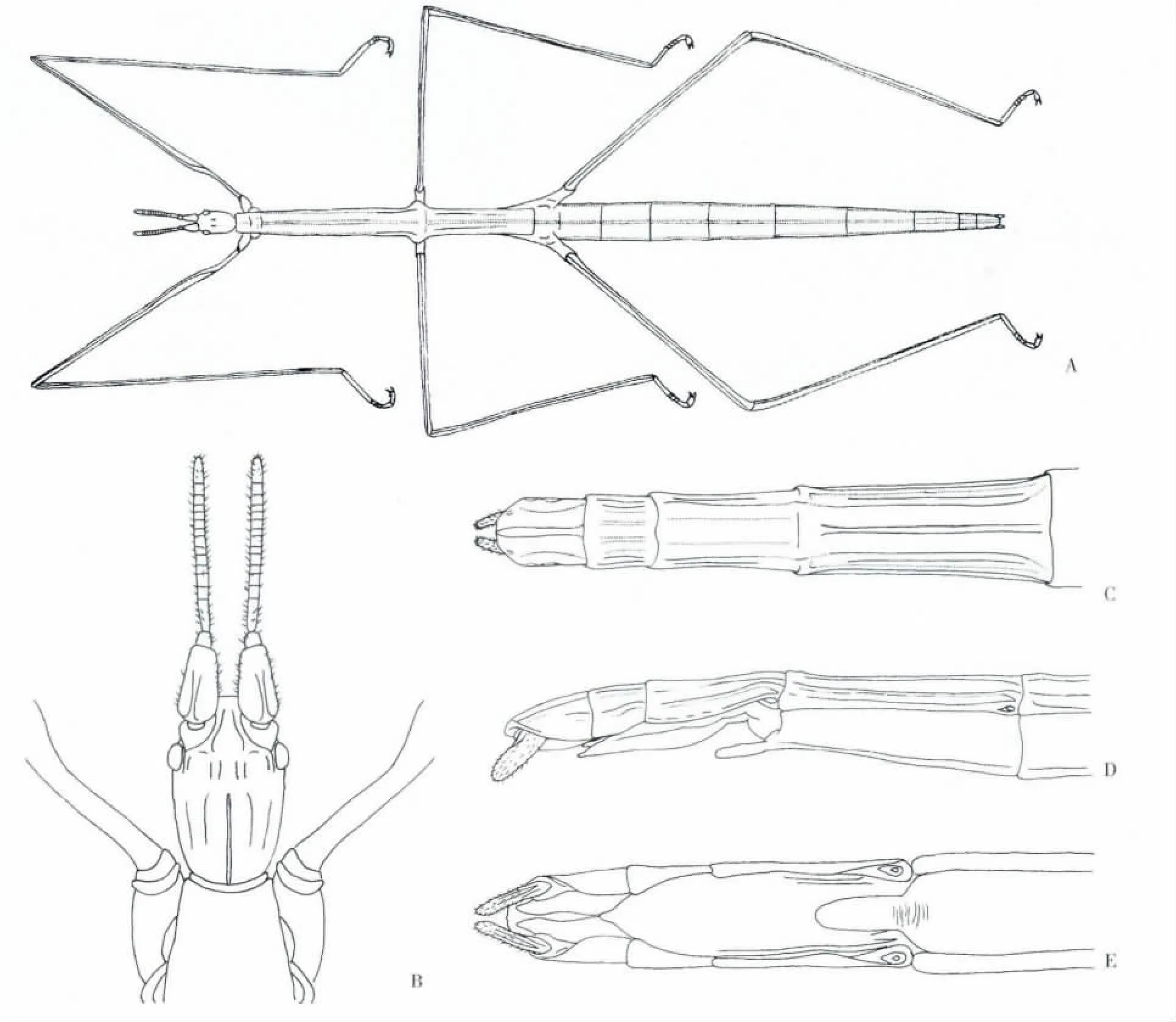
图1 A,荒漠竹节虫成虫)♀;B,头部背面观(♀);C,腹部第7-10 体节背面观(♀);D,腹部第7-10 体节侧面观(♀);E,腹部第8-10 体节腹面观(♀)Fig.1 A,Sceptrophasma bituberculatum ♀;B,Dorsal view of head(♀);C,Dorsal view of VII-X abdominal segments(♀);D,Lateral view of VII-X abdominal segments(♀);E,Ventral view of VIII-X abdominal segments(♀)
2.2 分布及栖息地环境
据记载本种分布于哈萨克斯坦,土库曼斯坦和中国新疆(БЕЙ-БИЕНКО,1964;蔡保灵,1987;Hennemann et al.,2008)。1980年至今的野外调查期间,仅在新疆伊犁河谷、天山低山山麓、准噶尔盆地荒漠仅发现了几次。它们多生活于有枯枝的干旱、半干旱的荒漠植丛中(图2),以其中一种绢蒿属Serphidium 植物为寄主(野外采集时,荒漠竹节虫的卵、若虫和成虫在其上均有发现)。据采集者称,白茨属Nitraria 植物亦可能是其寄主植物,初次室内喂养时,禾本科植物芨芨草Achnatherum splendens 也可被其取食。
2.3 产卵及卵的外形结构
2006年6月初第一次观察到了荒漠竹节虫的卵:长椭圆形,具灰、褐色图案,类似植物的种子,卵长4.9-5.5 mm,宽1.15-1.20mm,高1.0-1.2mm;卵盖一端为前极,略向背面弯曲,U 形卵孔杯位于卵背面后1/3 处,外壳坚硬而脆,表面具网状的纹饰,背面中线自卵孔杯之后一直延伸到卵囊末端。卵通常是单个排出,卵盖朝上,被分散地黏附在寄主植物的枝条上,具有很好的隐蔽性。自然条件下,曾最多在一株寄主植物上发现3 枚卵。室内饲养统计表明,每只雌虫产卵数为30-50 枚(图3,4,5)。

图2 野外的荒漠竹节虫雌虫Fig.2 A female Sceptrophasma bituberculatum in the field
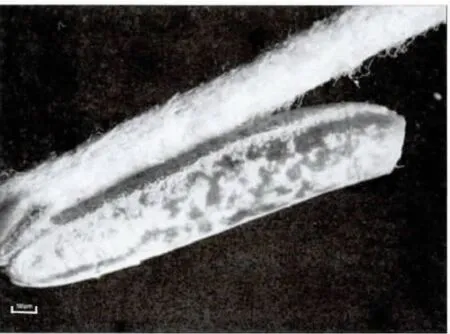
图3 荒漠竹节虫卵侧面观Fig.3 Lateral view of egg of Sceptrophasma bituberculatum
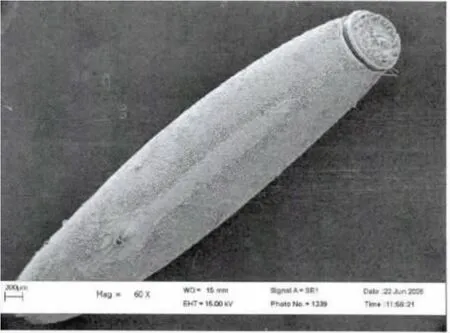
图4 荒漠竹节虫卵的亚显微结构Fig.4 Dorsal view of egg of Sceptrophasma bituberculatum by SEM
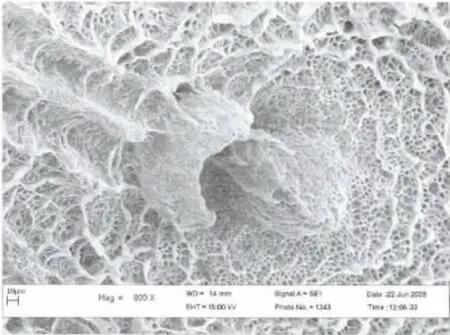
图5 卵孔杯表面亚显微结构Fig.5 Micropyle and sponge like surface of the egg by SEM
2.4 生活史
多年野外观察和室内饲养的结果表明:新疆分布的荒漠竹节虫一年一代,越冬个体常常出现在每年四月中旬(气温8-10℃持续数天后)至9月下旬(气温常保持在10-18℃),成虫常于5月中旬出现,在野外可持续存活至6月下旬。荒漠竹节虫具有逼真的拟态,成虫不仅在野外很难找寻,室内笼养时也常常会被它们迷惑,四龄若虫之前阶段的幼体更加难以被发现,在野外仅在极其偶然的情况下采集到8 头四龄以前的若虫,因此对若虫的研究主要来源于室内的饲养和观察。产卵最早见于5月18日,野外采集的成虫在人工饲养和食源充分的情况下,成虫的寿命和产卵期明显增长,可持续到7月中旬,卵约经47 d 开始孵化,孵化最早见于7月3日,孵化高峰期在7月下旬至8月中旬,室内卵的孵化可持续到11月中旬。一龄若虫细小,难以被发现,野外观察的数据是通过直接观察和对产在已标记寄主植物上卵的定期观察推论出来的:如卵壳完整,说明尚未孵化,如卵盖张开或脱落,内部变空,说明若虫已孵化。新生的若虫体长11-14 mm,身体修长柔软,淡黄绿色,拟态和生活行为方面与成虫相似。人工饲养条件下孵化的若虫龄期一般为17-22 d。野外生存的荒漠竹节虫在经过了三次蜕皮以后便开始准备越冬(9月后捕捉到的若虫均为4龄),而室内饲养的个体由于室温较高,虽不冬眠,在4 龄后亦不再蜕皮,直至年底死亡。

表1 荒漠竹节虫年生活史观察(自然条件和人工饲养条件下)Table 1 Observation of generational life history of Sceptrophasma bituberculatum both in the field and indoors in Urumqi
2.5 孵化率
观察期间,荒漠竹节虫在人工饲养条件下共产卵550 粒,291 粒孵化,孵化率为52.91%。考虑未孵化的卵存在滞育期,但经过观察,未孵化的卵直到来年的7、8月依旧没有丝毫孵化的迹象。对一些当年和前一年没有孵化的卵在显微镜下解剖观察,发现虽然有些卵即将完成胚胎发育,并且可以明显看出幼体结构,却没有丝毫生命迹象,并且散发着腐烂的味道(图6)。另外一些则根本没有发育,卵内物质呈干涸凝固状,约占原体积的1/2。
2.6 蜕皮

图6 几乎完全发育的死亡胚胎Fig.6 An almost fully developed dead embryo from unhatched egg
竹节虫目的昆虫为半变态发育,荒漠竹节虫需经过五次蜕皮羽化为成虫。人工纱笼中饲养的荒漠竹节虫每个龄期持续17-22 d,1-5 龄虫体体长分别为:10-14mm(图7),19-21mm,25-28 mm,32-39 mm 和46-54 mm。每次蜕皮前虫体明显膨胀,蜕皮时若虫紧紧攀附在寄主植物的枝干上,头部朝下弯向胸部,蜕皮从头部开始,然后是胸部,腹部,自然状态下蜕皮常常发生在早晨,历时约30 min,条件不适时可能发生在白天,蜕皮时间也会相应延长,如果受到惊扰可能还会终止并导致死亡。人工饲养初期,在塑料罐中饲养的若虫因没有提供枝条可攀附,曾观察到它们腹面向上静止躺在罐底,但第二天却发现它们再次“复活”,且身体明显变长并完成了一次蜕皮。若虫一般不取食虫蜕。

图7 荒漠竹节虫刚孵化的若虫Fig.7 A newly hatched nymph of Sceptrophasma bituberculatum
2.7 再生
荒漠竹节虫有附肢再生的现象。饲养观察期间,一头刚孵化的若虫在第二天便失去了右侧前足,经过17 d,发生第一次蜕皮以后,断损的足再生,包括腿节、胫节和跗节(仅有4个跗分节),腿节短而直,胫节和跗节向背方盘曲一圈余,爪呈芽状,缺角质尖端(图8);但该若虫5 d后死亡,没能继续观察到再生附肢的生长状况。而观察中,在五龄若虫阶段,附肢有断损,随着最后一次蜕皮,断损处并没有新的肢芽长出。

图8 二龄若虫再生的前足Fig.8 Regenerated fore leg of a second instar nymph
2.8 防御
荒漠竹节虫的自我防御行为较为温和,荒漠竹节虫在日间一直保持前足外伸的姿势,呈其寄主植物干枯的枝干状,与寄主植株融为一体,很难被发现,一旦有风吹草动,它们还会在枝干上来回摇曳模仿树枝的摆动;黄昏时分荒漠竹节虫比较活跃,有较强的取食行为,食量较大;受到外界惊扰时,便会立即逃向植丛深处,同时腹部向背方卷曲(模仿蝎的行为,其生境中常有蝎的分布)作为最常见的防御行为。
此外,舍弃附肢的自残行为应该也是它们逃脱敌害的一种防御方式,在户外采集时常可见到断足的虫体。经过观察,低龄若虫缺失的附肢有再生现象。没有假死和放射化学物质的防御行为。
2.9 越冬
与其他的竹节虫目的昆虫不同,荒漠竹节虫产卵后没有滞育现象,很快开始个体发育,多以四龄若虫度过寒冷的冬天。在野外采集中,2006年4月16日所采集的标本为四龄若虫,体长32 mm,当时室外温度达到8-10℃,2007年9月27日和2009年10月3日采集的标本依旧是四龄若虫(9月27日采集标本长度39 mm),当时室外温度为10-18℃,没有观察到具体的越冬行为和场所。
2.10 雌雄比例及繁殖方式
在对荒漠竹节虫的观察研究中,共采集标本54 头,均为雌体,未见若虫经室内单独饲养羽化,产出的卵可孵化为若虫,可确定孤雌生殖为其繁殖方式之一。
2.11 现状
过去20年的野外调查表明,目前在新疆北部的荒漠竹节虫呈点状分散分布。尽管在某些地区很难找到1 头虫体,但在合适的生境分布区中依旧存在着一定的种群,只要发现1 头,便会在它周围发现其他虫体,例如,2006年5月1日在一处蒿属植物丛生的山谷中,发现了12 头5 龄若虫。但是由于该物种无翅且活动性差,环境的变化和人类活动的影响极易使其种群数量受到威胁,随后2年在上述地点采集的个体就越来越少;同样的情形也发生在准噶尔盆地南缘的荒漠上。
随着近几年城市建设的剧烈膨胀及人工绿化力度的加大,荒漠竹节虫的自然生境受到了严重的破坏和威胁,分布地也越来越少,仅2006年至2013年间就有3个分布点消失。
3 结论与讨论
鉴于竹节虫目的昆虫多分布在热带、亚热带地区,同时有Whiting et al.(2003)的系统研究结果,我们推测分布在中亚干旱荒漠的荒漠竹节虫是一个古老的物种,它们所分布的区域曾经是类似于热带炎热潮湿的环境,随着气候的改变,荒漠竹节虫也随环境的变化发生了一定的适应性改变。
除了和寄主植物枝干逼真的拟态以外,荒漠竹节虫还具有许多荒漠昆虫相同的适应性特征,如体表较为坚硬、体色灰色或土黄褐色。同样,荒漠竹节虫所产的卵形态似植物的种子,卵壳的结构可以很好地在干旱的环境中保留水分,同时又透气保证了胚胎发育所需氧气(吴卫和范兆田,2010)。
据Bedford(1976,1978)研究,生活在热带的竹节虫目昆虫种类常不发生滞育,但卵的发育时间差异极大——即使是同一物种的同一只雌体在24 h 内产的卵也是如此,干燥的环境还会使Carausius morosus 卵的发育延长;生活在温带地区的种类因成体受光周期影响较大,卵的发育通常在冬季会发生滞育或发育延迟,如Bacillus rossius,长光照周期时产的卵不经历滞育,短光照时所产的卵需经历滞育。本文中荒漠竹节虫分布在温带内陆干旱区,产卵于长日照期(5月下旬至7月上旬),卵的发育没有滞育期,产卵后一个半月左右便开始孵化,与文献记载的其它需经历数月至2 a滞育的种类(陈树椿,2008)有很大差异,这种孵化方式应该也是它在进化中对干旱荒漠生境的一种适应,如同荒漠中的短命植物,如此可以最大程度地利用卵内的水分,防止因水分散失而导致的发育失败;对未孵化卵的解剖表明,其中一部分卵的卵质干涸,这可能是其孵化失败的主要原因,而另一部分卵发育失败则是因为受到了微生物的感染。人工饲养条件下温湿度比较恒定,实验记录卵的孵化时间甚至可持续至12月初,说明温度和湿度对胚胎发育有影响。荒漠竹节虫的孵化是在一年中最炎热、干旱的季节,寄主植物几近干枯,食源状况不利于其若虫的生长,但在生存策略上讲,比起会因滞育而导致卵内水分散失最终不能孵化,应属权宜之计了。
本文观察到荒漠竹节虫以四龄若虫越冬。新疆冬季寒冷,尤其是荒漠,常见昆虫通常会钻入沙土中或石块下方越冬,但竹节虫体型细长,难以做到,结合其生境,推测它们多隐蔽在寄主植物的枝丛中或者沙石土缝中来度过漫长的冬季,荒漠竹节虫越冬的机制仍需进一步观察研究。
孤雌生殖是昆虫中较为常见的生殖方式,也是昆虫适应恶劣环境的一种表现(王成业,2011)。文献记载荒漠竹节虫有雄体(Бей-Биенко,1964;陈树椿和何允恒,2008;Hennemann et al.,2008)。而在我们历年来野外采集的54 头标本中(四龄若虫9 头,五龄成虫45头)未见雄虫,将五龄虫体单独室内饲养至产卵表明孤雌生殖是其繁殖方式之一;具体在何时出现雄体、并且发生两性生殖尚需进一步研究。
因荒漠竹节虫拟态性很强、野外难以发现,本研究结果是建立在室内外共同观察的结果之上。观察表明:温度、湿度和食物对荒漠竹节虫的生存影响很大。其中,室外温差表现为日夜温差(一般大于10℃)、季节温差(极差可达55-80℃);夏季日间荒漠湿度常常低于10%,而室内则始终为30%左右;荒漠竹节虫对食物的要求也较高,例如,对从阜康荒漠和雅马里克山采集回来的竹节虫同时投喂两个采集地的蒿,它们都明显只取食自己采集地的。因此,室内饲养的低龄幼虫未能进入下一龄期就死亡的数量较多;从野外采集带回室内饲养的四龄若虫因室内温度等原因不能进入越冬状态,但也不会继续发生蜕皮最终死亡;但同时室内饲养条件下,成虫寿命较野外延长很多,产卵时间也相应延长;卵的孵化可随之持续。
荒漠竹节虫在新疆是体长最长的昆虫,由于数量很少,在林、牧业上虽没有十分重要的经济意义,但仍然是一种珍稀的值得收藏的昆虫,尤其在昆虫对荒漠环境的适应和进化研究上是一种很好的研究材料。在此我们仅对荒漠竹节虫的一般形态特征和生物学特征进行了研究,今后将对它在荒漠环境中的适应性及遗传多样性方面进一步研究,为对该物种的多样性保护提供参考。
References)
Bedford GO.Biology and ecology of the Phasmatodea[J].Annual Review of Entomology,1978,23(1):125-149.
Bedford GO.Description and development of the eggs of two stick insects(Phasmatodea:Phasmatidae)from New Britain[J].Australian Journal of Entomology Society,1976,15(4):389-393.
Cai BL.A List of Chinese phasmids with the key to subfamilies[C].In:Contributions From Tianjin Natrual Museum,1987,4:26-30.[蔡保灵.中国竹节虫名目及亚科检索表[C].见:天津自然博物馆论文集,1987,4:26-30]
Chen PC,Chen SC.Identification,biology and control of the important species of phasmatodea in China[J].Journal of Beijing Forestry University,1997,19(4):70-75.[陈培昶,陈树椿.中国重要竹节虫的鉴别、生物学及其防治[J].北京林业大学学报,1997,19(4):70-75]
Chen SC,He YH.Phasmatodea of China[M].Beijing:China Forestry Publishing House,2008,1-476.[陈树椿,何允恒.中国虫脩目昆虫[M].北京:中国林业出版社,2008,1-476]
Hennemann FH,Conle OV,Zhang WW.Catalogue of the stick and leaf-insects(Phasmatodea)of China,with a faunistic analysis,review of recent ecological and biological studies and bibliography(Insecta:Orthoptera:Phasmatodea)[J].Zootaxa,2008,1735:1-77.
Huang RX,Wu W,Mao XF,et al.The Fauna of the Desert Insects of Xinjiang and its Formation and Evolution[M].Urumqi:Xinjiang Science and Technology Publishing House,2005,1-232.[黄人鑫,吴卫,毛新芳,等.新疆荒漠昆虫区系及其形成与演变[M].乌鲁木齐:新疆科技出版社,2005,1-232]
Whiting MF,Bradler S,Maxwell T.Loss and recovery of wings in stick insects[J].Nature,2003,421(6920):264-267.
Schwander T,Crespi BJ.Multiple direct transitions from sexual reproduction to apomictic parthenogenesis in Timema stick insects[J].Evolution,2009,63(1):84-103.
Wang CY.Genetic mechanism and evolutionary significance of the origin of parthenogenetic insects[J].Zoological Research,2011,32(6):689-695.[王成业.昆虫孤雌生殖起源的遗传机制和进化意义[J].动物学研究,2011,32(6):689-695]
Бей-Биенко ГЯ,1964.Определитель Насекомых Европейской Части Ссср[M].В Пяти Томах Ι.Издательство《Наука》.Москва-Ленинград:203-204.
Ye WY,Yin LK,Qian Yi et al.Study on the vegetation of Yama-like Mountain and plant diversity characteristic[J].Academic Journal of Xinjiang Agriculture University,2005,28(1):44-48.[叶卫英,尹林克,钱翌,等.乌鲁木齐市周边荒山植物群落类型及其物种多样性[J].新疆农业大学学报,2005,28(1):44-48]
