靶向人表皮生长因子受体3的人源Fab抗体构建及鉴定
2015-11-29张晶刘玉杰万德有刘蕴慧王芳武逸熏付洁宋海峰
张晶 ,刘玉杰,2,万德有,刘蕴慧,王芳,武逸熏,付洁,宋海峰
1.军事医学科学院 放射与辐射医学研究所,北京 100850;2.广西医科大学,广西 南宁 530000
人表皮生长因子受体3(human epidermal growth factor receptor 3,HER3)是表皮生长因子受体(epidermal growth factor receptor,EGFR)家族的一员,该家族是目前最重要的癌症治疗靶点之一,被广泛研究并应用于多种癌症的研究及治疗中,如乳腺癌、宫颈癌、胃癌、肺癌等。早在2005 年就有了第一个完全人源化的针对该家族中EGFR 的抗体药物,而以HER2 为靶点的抗体药物赫赛汀更是2014年公认的明星药物之一。尽管C 端缺少激酶活性,但HER3 通过与HER2 和EGFR 形成异源二聚体,促发下游通路激活,在靶向HER2及EGFR的抗体类药物如赫赛汀、西妥昔单抗等抗体类药物的耐药中HER3 补偿性表达上调。因此,HER3 在机体耐药机制中发挥重要作用[1-3]。自2010 年AVEO Pharma 完成首个HER3 抗体项目起[4],已有多家公司和研究机构针对HER3 靶点进行抗体研发工作[5],且数量呈逐年增多状态。
抗体-药物偶联物(antibody conjudge drug,ADC)是目前抗体应用的趋势之一,如Seattle Genetics 公司以CD30 为靶点的Adcetris、罗氏公司以HER2 为靶点的Kadcyla。目前ADC 药物虽然以全长型抗体为主,但药物分子越大越难以透过毛细管内皮层及穿过肿瘤细胞外间隙到达实体瘤的深部。基于上述不足,小型化或适度小型化是研制ADC 药物的重要途径,如scFv、Fab 类基因工程抗体受到越来越多的关注及应用[6-8]。这类抗体因含有完整的抗体可变区而保留了与抗原特异性结合的能力,同时由于其分子小而更易于与其他药物制备成不同形式的更为安全的疫苗和药物制剂,同时也更易于通过较为经济的方式获得抗体制备成诊断试剂[9]。
本实验室前期构建了哺乳动物细胞展示型全人源scFv 抗体库,并通过流式细胞分选技术获得了靶向人HER3 胞外区蛋白的scFv 抗体[10]。在本实验中,我们为将HER3 scFv 抗体改构成Fab 形式,利用真核表达载体p3457 构建了HER3 Fab 抗体真核表达质粒,并通过293E 悬浮细胞进行表达,对其亲和力及初步的生物学活性进行鉴定,为后续相关研究奠定基础。
1 材料与方法
1.1 材料
293T、293E 细胞由本实验室保存;大肠杆菌DH5α购自北京原平皓生物技术公司;pPICZα AVH-CH-VL-CL、pDisplay-scFv、p3457 均由本实验室构建并保存。
293E 细胞培养基购自Gbico 公司;BspQⅠ购自NEB 公司;PrimeSTAR 购自TaKaRa 公司;DNA maker 购自北京博迈德生物技术有限公司;DNA 琼脂糖胶回收试剂盒购自康为世纪生物技术有限公司;PEI转染试剂购自Invitrogen 公司;HRP 标记的抗His 抗体购自西美杰生物技术公司;兔抗人ERBB3(Nterm)单克隆抗体购自ABGENT 公司;罗丹明标记的抗兔IgG(H+L)多克隆抗体购自KPL 公司;DAPI 购自碧云天生物技术研究所;CCK8检测试剂盒购自北京天恩泽基因科技有限公司;引物由上海生工生物技术有限公司合成。
1.2 HER3 Fab型抗体表达载体构建
1.2.1 VH、VL、CH1、CL基因扩增 抗人源HER3抗体模板来源于本实验室前期从真核哺乳动物细胞展示型人源scFv 抗体库筛选的pDisplay-scFv 质粒[11]。为从pDisplay-scFv 中扩增VH 及VL,设计引物PHfw/PHrv、PLfw/PLrv(表1),并在引物的5'端引入限制性酶切位点BSpQⅠ;为构建Fab 型重链和轻链,设计引物PHfw'/PHrv'、PLfw'/PLrv'(表1),分别从本实验室前期构建的VEGF 全长抗体质粒pPICZα A-VH-CH-VL-CL[11]中扩增CH1及CL基因。
VH、VL 的PCR 反应体系:pDisplay-scFv 1 μL,5×PrimeSTAR 缓冲液10 μL,dDTP 混合液4 μL,Primestar DNA 聚合酶0.5 μL,PHfw/PHrv 或PLfw/PLrv(10 μmol/L)各1 μL,加ddH2O至50 μL。
CH1、CL 的PCR 反应体系:pPICZα A-VH-CHVL-CL 1 μL,5×PrimeSTAR 缓冲液10 μL,dDTP 混合液4 μL,Primestar DNA 聚合酶0.5 μL,PHfw/PHrv 或PLfw/PLrv(10 μmol/L)各1 μL,加ddH2O 至50 μL。
PCR 反应参数:98℃ 8 min;98℃ 30 s,55℃30 s,72℃40 s,28个循环;72℃10 min。
1.2.2 HC、LC 基因扩增 分别用DNA 琼脂糖胶回收试剂盒回收目的大小正确的VH、VL、CH1、CL,详细步骤参见胶回收试剂盒说明书。分别通过重叠PCR 获 得HC 及LC。重链PCR 反应体系:CH1 1 μL,VH 1 μL,5×PrimeSTAR 缓冲液10 μL,dDTP混合液4 μL,Primestar DNA 聚合酶0.5 μL,PHfw/PHrv'(10 μmol/L)各1 μL,加ddH2O 至50 μL。轻链PCR 反应体系:CL 1 μL,VL 1 μL,5×Prime-STAR 缓冲液10 μL,dDTP 混合液4 μL,Primestar DNA 聚合酶0.5 μL,PLfw/PLrv'(10 μmol/L)各1 μL,加ddH2O至50 μL。
1.2.3 p3457-HC 及p3457-LC 质粒构建 分别用BspQⅠ酶切重链、轻链及载体p3457。酶切体系:HC/LC/p3457 2 μg,BspQⅠ1 μL,缓冲液1 μL,加ddH2O 至50 μL。50℃孵育2 h,65℃灭活20 min。酶切产物回收后经T4DNA 连接酶于16℃连接过夜。连接体系:HC、LC 各7 μL,p3457 1 μL,T4DNA 连接酶1 μL,T4缓冲液1 μL。将连接产物转化大肠杆菌感受态DH5α,对挑取的单克隆进行质粒小量提取和菌液PCR 鉴定,挑选阳性单克隆进行测序分析和表达鉴定。
1.3 p3457及p3457质粒表达鉴定
用含10%新生牛血清和双抗(工作浓度为100U/mL 的青霉素及工作浓度为0.1 mg/mL 的链霉素)的DMEM 培养基于37℃、5% CO2细胞培养箱中培养293T 细胞,接种50 000细胞(贴壁时密度约30%)于24 孔板中,培养24 h,至细胞密度达70%时,按PEI转染试剂说明书分别转染p3457-HC 及p3457-LC,转染48 h 后收细胞上清,经12.5% SDS-PAGE 分离后,电转移至硝酸纤维素膜上,经5%脱脂奶粉室温封闭1 h,再用1∶5000稀释的HRP标记的小鼠抗His单克隆抗体室温孵育1 h,经TBST 漂洗3 次,每次7 min,用化学发光法显色5 min,压片显影。

表1 PCR引物序列
1.4 HER3 Fab抗体表达条件摸索及纯化
以0.8×106/mL 的密度接种处于对数生长期的293E细胞,于37℃、5% CO2培养至细胞浓度为(1.4×106~2.0×106)/mL 时,按PEI 转染试 剂说明书将p3457-HC 及p3457-LC 分别按1∶1 的比例瞬时转染,第3 d 向细胞中添加10×CB5(储存浓度为33 g/L),并开始每天收集1 mL 细胞上清,第5 d 完全收集细胞,800 r/min 离心10 min 后收上清并用Western印迹检测细胞上清中HER3 Fab的表达水平。
按优化后的条件大量培养细胞以表达HER3 Fab 抗体,第5 d 收集细胞上清后用0.45 μm 滤膜过滤,用镍亲和柱纯化细胞上清。平衡液配方:NaCl 500 mmol/L,磷酸钠缓冲液20 mmol/L,pH7.4;洗脱液配方:NaCl 500 mmol/L,磷酸钠缓冲液20 mmol/L,咪唑500 mmol/L,pH7.4。1~10 个柱体积内咪唑浓度线性上升到100%洗脱,收集洗脱液,用PBS 置换缓冲液,将蛋白溶液分装保存于-80℃。
1.5 ELISA检测HER3 Fab与HER3的结合
用HER3 胞 外区(extracelluar membrane domain,ECD)蛋白(6 nmol/L)包被酶标板,4℃包被过夜;用1% BSA 于37℃封闭2 h;加入不同稀释率的HER3 Fab 抗体,37℃孵育1 h;PBST 洗涤6 次,加入HRP 标记的抗His 抗体(1∶10000),37℃孵育1 h;PBST 洗涤6 次,加入TMB 底物显色。以商品化的抗ERBB3(N-term)抗体为阳性对照,以PBS 替代HER3 Fab抗体为阴性对照。
1.6 ForteBio法鉴定HER3 Fab的亲和力
采用ForteBio 法测定HER3 Fab 抗体与HER3的平衡解离常数KD。将HER3胞外区蛋白生物素化后,以20 μg/mL 的浓度固定在链霉亲和素探针上,用PBS 平衡探针后,再与梯度稀释的HER3 Fab 抗体(500、250、125、62.5、31.25、15.625 nmol/L)结合,以PBS 作为空白对照,然后探针再孵育在PBS 中进行解离。
1.7 CCK8 法检测HER3 Fab 抗体对MDA-MB-453细胞增殖的影响
将处于对数生长期的MDA-MB-453 细胞消化成单个细胞,以4000/孔接种到96 孔板中,24 h 后去除上清,第1、2组分别加入50 μL 完全培养基,第3、4组分别加入50 μL含40 μg/mL HER3 Fab抗体的完全培养基;1 h后第1、3组分别加入50 μL 完全培养基,第2、4组分别加入50 μL含200 ng/mL神经调节蛋白1(NRG1)的完全培养基;每组设3 个复孔,24 h时分别加入10 μL CCK8,37℃孵育1 h后用酶标仪测定D450nm值。
2 结果
2.1 质粒p3457-HC、p3457-LC及其表达产物的鉴定
质粒构建流程如图1。VH、VL、CH1、CL PCR产物经2%琼脂糖凝胶电泳鉴定分别可见359、309、315、315 bp 条带(图2A);VH 及CH1 重叠PCR 产物HC、VL 与CL 重叠PCR 产物LC 经1.5%琼脂糖凝胶电泳鉴定分别可见674、624 bp 条带(图2B);质粒p3457-HC 及p3457-LC 的大小分别为5184 及5134 bp。挑选单克隆经菌液PCR 鉴定后分析均为阳性(图2C)。挑选菌液PCR 阳性的单克隆进行小量质粒提取并测序,编码区序列完全正确(数据略),说明克隆构建成功。Western 印迹结果显示,p3457-HC及p3457-LC 质粒分别瞬时共转染293E 细胞的表达产物在相对分子质量约25×103处可见目的条带(结果略)。在瞬时共转染293E 时,第5 d 时抗体表达量最高(图3)。
2.2 抗体结合实验及亲和力检测结果
ELISA 检测表明HER3 Fab 能与HER3 胞外区蛋白直接结合(图4)。采用ForteBio 实验测定单克隆抗体与HER3 的平衡解离常数KD为1.08×10-8mol/L。
2.3 HER3 Fab 对MDA-MB-453 细胞增殖能力的影响
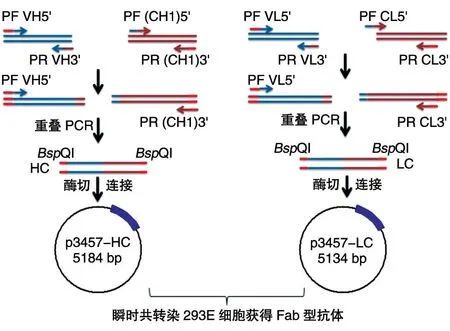
图1 p3457-HC、p3457-LC表达载体的构建
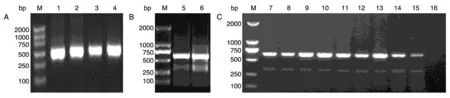
图2 HER3 Fab质粒的构建及鉴定
在NRG1 刺激的条件下将HER3 Fab 抗体与MDA-MB-453 细胞共同孵育24 h,CCK8 法检测结果表明NRG1 刺激组(第2 组)与未刺激组(第1 组)结果有显著差异(P=0.03),说明细胞经100 ng/mL NRG1刺激后HER3膜表达增加,细胞增殖明显。同时,在NRG1同等刺激条件下,加入HER3 Fab组(第3 组)与未加入HER3 Fab 组(第2 组)相比结果有差异(P=0.07),说明HER3 Fab 抗体可以显著抑制MDA-MB-453细胞增殖。结果见图5。

图3 不同表达时间点HER3 Fab的表达鉴定

图4 ELISA检测HER3 Fab与HER3的结合
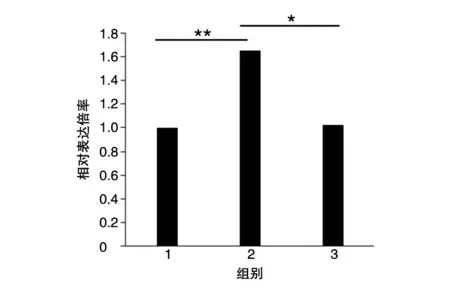
图5 CCK8检测HER3 Fab对MDA-MB-453细胞增殖的抑制性
3 讨论
目前哺乳动物细胞表达抗体的2 种形式是瞬时转染和构建稳定细胞系。一般在大量生产抗体类蛋白应用于试剂盒开发或抗体药物时,会使用稳定转染的方式,但实验研究新的抗体时一般采用瞬时转染的方式。瞬时转染表达抗体一般是将抗体的轻链和重链分别构建到2 种载体上,再通过转染试剂共转染到同一细胞中,细胞分别翻译出轻、重链,再在内质网中通过二硫键将轻、重链合成为抗体形式。
一般情况下,293E 哺乳动物悬浮细胞表达抗原蛋白时仅需要3 d,而表达抗体类蛋白时需要4~6 d。在本实验中,我们验证了蛋白在不同表达时间的表达水平,结果显示在转染后第3 d 抗体开始表达,并随着时间的延长表达水平逐渐提高但细胞活率逐渐下降,在第5和第6 d抗体的表达量水平一致。考虑到细胞活率,我们在第5 d 时收取细胞上清纯化蛋白。
HER3 属于酪氨酸激酶受体家族,经配体刺激后触发构型改变并与该家族中其他成员EGFR、HER2,特别是HER2 形成异源二聚体。NRG1 是促发HER3 下游通路激活的重要因子之一,多种与HER3 抗体研究有关的实验设计都用NRG1 刺激靶细胞。在本研究中,我们还设置了在未经NRG1 刺激的情况下加入HER3 Fab 的组别,结果显示该条件下Fab 抗体的孵育对细胞增殖无影响,但是在刺激情况下Fab显著抑制细胞增长,该现象与HER3的膜表达量呈正相关,与文献报道相符。
综上,我们构建、表达并纯化获得了平衡解离常数KD为1.08×10-8mol/L 的靶向人HER3 的Fab 型抗体,其在NRG1 刺激下对MDA-MB-453 细胞的增殖有一定的抑制能力,这为后期对HER3 高表达型癌症的研究和诊断提供了一种新的思路与方法。
[1]Schoeberl B,Faber A C,Li D,et al.An ErbB3 antibody,MM-121,is active in cancers with ligand dependent activation[J].Cancer Res,2010,70(6):2485-2494.
[2]Kugel C H 3rd,Hartsough E J,Davies M A,et al.Functionblocking ERBB3 antibody inhibits the adaptive response to RAF inhibitor[J].Cancer Res,2014,74(15):4122-4132.
[3]Ma J,Lyu H,Huang J,et al.Targeting of erbB3 receptor to overcome resistance in cancer treatment[J].Mol Cancer,2014,13:105.
[4]Meetze K,Vincent S,Tyler S,et al.Neuregulin 1 expression is a predictive biomarker for response to AV-203,an ERBB3 inhibitory antibody,in human tumor models[J].Clin Cancer Res,2015,21(5):1106-1114.
[5]Aurisicchio L,Marra E,Roscilli G et al.The promise of anti-ErbB3 monoclonals as new cancer therapeutics[J].Oncotarget,2012,3(8):744-758.
[6]Roque C,Sheung A,Rahman N,et al.Effect of polyethylene glycol conjugation on conformational and colloidal stability ofa monoclonal antibody antigen-binding fragment(Fab')[J].Mol Pharm,2015,12(2):562-575.
[7]Hristodorov D,Mladenov R,Brehm H,et al.Recombinant H22(scFv) blocks CD64 and prevents the capture of anti-TNF monoclonal antibody[J].MAbs,2014,6(5):1283-1289.
[8]Gu J,Lei Y,Huang Y,et al.Fab fragment glycosylated IgG may play a central role in placental immune evasion[J].Hum Reprod,2015,30(2):380-391.
[9]Shukra A M,Sridevi N V,Chandran D,et al.Production of recombinant antibodies using bacteriophages[J].Eur J Microbiol Immunol,2014,4(2):91-98.
[10]Zhang J,Zhang X,Liu Q,et al.Mammalian cell display for rapid screening scFv antibody therapy[J].Acta Biochim Biophys Sin,2014,46(10):859-866.
[11]陈薇,张晶,刘强,等.VEGF 全长抗体在毕赤酵母中的表达与活性鉴定[J].生物技术通讯,2014,25(4):492-496.
