枯草芽孢杆菌表达系统及其启动子研究进展
2015-10-26余小霞田健刘晓青伍宁丰
余小霞 田健 刘晓青 伍宁丰
(中国农业科学院生物技术研究所,北京 100081)
枯草芽孢杆菌表达系统及其启动子研究进展
余小霞 田健 刘晓青 伍宁丰
(中国农业科学院生物技术研究所,北京 100081)
枯草芽孢杆菌作为一种革兰氏阳性细菌,由于其具有非致病性、分泌蛋白能力强的特性和良好的发酵基础及生产技术,是目前原核表达系统中表达和分泌外源蛋白的理想宿主,成为原核表达系统中的一种重要的模式菌株。而实现外源蛋白的高效表达的关键因素之一是使用强并可控制的启动子。目前,枯草芽孢杆菌中常用的启动子为组成型、诱导物诱导型、时期特异性及自诱导型。详细介绍枯草芽孢杆菌表达系统以及其常用启动子的优缺点,并对克隆新的启动子的方法做了总结,旨为完善枯草表达系统和工业生产外源蛋白奠定基础。
枯草芽孢杆菌;表达系统;组成型启动子;诱导型启动子
枯草芽孢杆菌(Bacillus subtilis)是一种重要的原核表达宿主,具有很高的应用价值,其培养简单快速,具有较强的分泌蛋白质的能力、非致病性及良好的发酵基础和生产技术,是目前生产各种工业用酶,如淀粉酶、蛋白酶和脂肪酶等的理想表达宿主,是微生物研究领域中的一种重要模式菌株。但是目前,枯草芽孢杆菌表达系统仍然存在不完善的地方,大部分外源蛋白在其体内的表达仍旧不高,这主要是因为:(1)枯草芽孢杆菌在其对数生长末期表达和分泌大量的蛋白酶,影响外源蛋白的稳定性和产量;(2)质粒的分化和结构不稳定性,有时外源蛋白的组成型表达会影响质粒的稳定性;(3)毒性蛋白在其体内不易表达;(4)有些外源蛋白的表达分泌到培养基中会影响宿主菌的生长;(5)分子遗传操作与大肠杆菌相比较困难。这些因素限制了枯草杆菌表达系统的发展。枯草芽孢杆菌168菌株的基因组全序列测序工作已于1997年完成[1],但到目前为止,其基因组中尚有近50%[2]的基因的功能仍未阐明。因此目前针对枯草芽孢杆菌中基因及相关元件功能,以及其对内部及外界环境响应的研究仍是针对该菌基础研究的重点。
外源蛋白在枯草芽孢杆菌中的高效表达是实现其在工业应用上的重要途径,而使用强并可控制的启动子是外源蛋白高效表达的关键因素之一,决定着工业生产酶的大规模生产。
本文简单介绍了枯草芽孢杆菌表达系统的特点和表达目的蛋白时使用的载体系统,重点介绍枯了枯草芽孢杆菌表达系统中的重要组成部分——启动子,并对启动子的分类和克隆进行了详细的叙述,最后总结了枯草芽孢杆菌表达系统的现状及展望。
1 枯草芽孢杆菌表达系统的概述
早在100多年前,人们就开始对枯草芽孢杆菌进行研究,但早期的工作主要针对其分离鉴定、形态观察及功能鉴定等进行研究。1958年,Spizizen[3]发现枯草芽孢杆菌可以在自然条件下形成感受态,摄入外源DNA,从而建立了枯草芽孢杆菌化学感受态制备方法。此后,随着DNA重组技术的发明,枯草芽孢杆菌表达系统成为研究热点。
枯草芽孢杆菌是一种存在着发育过程的原核生物,该菌在细胞的培养过程中具有典型的生长规律,如生长延滞期、对数期、稳定期、衰亡期及芽孢发育期。芽孢杆菌通常是采用更换不同的σ因子的方式来实现细胞中不同基因的特异性表达。枯草芽孢杆菌中的σ因子有多种[4],其中σA、σB、σC、σD、σH、σI、σX、σW与枯草芽孢杆菌营养生长相关,且枯草芽孢杆菌中的σA相当于大肠杆菌σ70,是营养生长最主要的σ因子,占σ因子总量的90%-95%,负责转录持家基因和孢子形成早期基因;而σE、σF、σG、σK与孢子形成有关。枯草芽孢杆菌表达系统的另一特点是其分泌系统,由于枯草芽孢杆菌是革兰氏阳性细菌,仅有一层细胞膜。因此,蛋白质的分泌无需转位至周质空间(革兰氏阴性细菌,如大肠杆菌需经过此步骤)而直接通过细胞质转移到胞外。通过生物信息学的预测,枯草芽孢杆菌中共有294种蛋白质被预测为分泌型蛋白质[5]。根据分泌型蛋白有无典型的信号肽,可以将其分泌系统分为依赖信号肽的分泌途径和不依赖信号肽的分泌途径。由于对不依赖信号肽的分泌途径的研究很少,且与依赖信号肽的分泌途径不同,因此又将其称为非典型分泌途径[6]。
枯草芽孢杆菌中至少存在4种依赖信号肽的分泌途径[7]:Sec分泌途径(Sec-SRP cooperation pathway)、Tat分泌途径(Twin-arginine translocation pathway)、ABC转运子途径(ATP-binding cassette transporters)、假菌丝蛋白输出途径(Pseudopilin export pathway)。Sec分泌途径是枯草芽孢杆菌中的主要分泌途径,经此系统分泌的蛋白质含有Sec型信号肽[8];Tat分泌途径一般只分泌已形成正确构象的胞内蛋白质,经此系统分泌的蛋白质含有Tat型信号肽[8];ABC转运子途径主要用于细菌素等小分子的运输[9];假菌丝蛋白输出途径与枯草芽孢杆菌细胞感受态的形成有关[10]。虽然枯草芽孢杆菌有强的胞外蛋白分泌能力,但较遗憾的是外源蛋白在其体内的表达量不高,主要原因有以下两种:(1)枯草芽孢杆菌表达系统不成熟;(2)枯草芽孢杆菌野生型菌株本身会产生大量的胞外蛋白酶,容易造成目的蛋白的降解。
枯草芽孢杆菌应用的最为广泛的宿主菌是枯草168菌株,目前已经知道该菌株可产生8种胞外蛋白酶(其中不包括一些未发现的蛋白酶),利用基因突变的方法失活枯草168菌株基因组上中性蛋白酶、碱性蛋白酶、金属蛋白酶、胞内蛋白酶、芽孢杆菌肽酶F以及中性蛋白酶B六种蛋白酶基因后,将该菌株命名为WB600,其蛋白酶活性仍保留野生型菌株的0.32%[11]。WB700是在WB600的基础上将丝蛋白酶基因vpr在内的7个蛋白酶基因全失活的突变株,其蛋白酶活力是野生菌株的0.1%。随后,来自加拿大卡加利大学的Wong等[12]在WB700的基础上,继续将枯草杆菌的胞壁蛋白酶基因cwp失活,发展了在野生菌株的基础上缺失了8个蛋白酶基因的胞外蛋白酶活性更低的WB800菌株。因此,现在用于外源表达目的蛋白常用的枯草杆菌菌株大部分使用的是枯草168系列的突变株。
2 枯草芽孢杆菌表达载体系统
克隆载体和表达载体是基因重组技术的重要工具,在分子遗传操作的所有领域中处于十分重要的地位。根据类型的不同可将其分为3类[13]:独立自主复制的质粒载体、整合型载体及噬菌体载体。下面主要介绍在枯草芽孢杆菌中常用的独立自主复制质粒载体和整合型载体。
2.1 独立自主复制的质粒载体
枯草芽孢杆菌168系列菌株目前已全部完成测序,成为研究枯草芽孢杆菌的标准菌株,大部分的枯草芽孢杆菌不含有内源性质粒,如今常用的质粒大部分来自于葡萄球菌和链球菌[14,15]。这些质粒载体可在枯草芽孢杆菌中自主复制,根据其复制方式可分为滚环复制型载体(Rolling circle-type replication vectors)和θ复制型载体,大部分来自于革兰氏阳性细菌的小的质粒载体(<12 kb)是通过滚环复制机制复制的,而一些大的质粒载体是通过θ复制机制复制的。如最开始鉴定的来自于金黄色葡萄球菌的4个质粒 pUB110,pC194,pE194和pT181都是通过滚环机制复制的。pUB110带有卡那霉素的抗性基因,拷贝数较低,每个细胞中有30-50个拷贝[16];pC194带有氯霉素的抗性基因,拷贝数约为15[17];pE194较特殊,是个温度敏感性的质粒载体,32℃正常复制,随着温度增加,拷贝数降低,45℃时停止复制,这为人们纯化含有此类质粒的细胞提供了一个思路。另一方面,pE194可以在细菌基因组上多个位点发生特异性整合,整合子可以通过抗性(红霉素)和培养温度(50℃)进行筛选[18,19]。pT181带有四环素的抗性基因,每个细胞中约有20个拷贝[18]。
这些质粒一般可以在枯草芽孢杆菌中自主复制,表达其抗性基因,但在传代培养中仍存在许多问题,其中最大的问题是质粒的分离不稳定性和结构不稳定性[20]。质粒分离不稳定性表现在细胞在无筛选压力条件质粒易丢失;结构不稳定包括质粒DNA的重排、筛减和增加。质粒载体的不稳定性严重影响了其在枯草芽孢杆菌的研究应用,如许多基因不能直接构建在枯草芽孢杆菌质粒载体,转入宿主菌进行表达。然而,大肠杆菌分子遗传操作成熟,不会产生质粒不稳定性现象,所以构建大肠杆菌-枯草芽孢杆菌的穿梭质粒载体是十分必要的,这样就可以将外源基因在大肠杆菌中构建完成后再转化到枯草芽孢杆菌中表达。目前常用的穿梭质粒载体有 pEB10[21]、pEB20[22]、pEB60[22]、pUB18[23]、pUB19[24]和pWB980[24]等,详细信息,见表1。

表1 常用的大肠杆菌-枯草芽孢杆菌穿梭质粒载体
2.2 整合型质粒载体
芽孢杆菌中高效表达外源蛋白的最大障碍是构建的重组表达质粒在宿主菌中通常不稳定,解决这一问题的有效途径是使用整合型载体。枯草整合型质粒载体一般含有大肠杆菌质粒的复制起点(通常为pBR322或其衍生质粒)、抗性筛选标记基因(常用于枯草的标记基因有卡那霉素、氯霉素、红霉素等)及一段或两段整合基因(即与宿主细胞染色体序列同源的基因)。它的特点是限制性复制,即整合型质粒只在大肠杆菌中有复制功能,转入革兰氏阳性细菌如枯草芽孢杆菌中则无此功能,因此只有整合进宿主基因组中才能正常的表达目的基因。整合型质粒载体根据其整合方式不同可分为单交换整合和双交换整合。
2.2.1 单交换整合 通过一段位于外源基因某侧的同源序列(与枯草芽孢杆菌基因组同源的序列),与枯草芽孢杆菌基因组的目标序列发生同源重组,从而将外源基因整合到染色体上,单交换需要的同源臂较短,整合效率高且稳定,不易发生逆整合(图1)。

图1 单交换整合型载体
2.2.2 双交换整合 通过位于外源基因两侧的同源序列,与枯草芽孢杆菌基因组的目标序列发生同源重组。为了提高双交换的整合效率,其需要的同源臂长度至少要400-500 bp[25],才能发生有效的重组;其次还需要考虑发生重组的线性双链DNA的稳定性。因为线性双链DNA导入枯草芽孢杆菌体内容易被AddAB解旋酶降解,但若在线性双链DNA末端加上Chi位点(χBs,5'-AGCGG-3')可以解决此问题[26,27],因此在构建重组质粒载体时可以在克隆同源臂时引入Chi位点以提高重组效率。目前常用的双交换整合型载体是通过amyE基因位点进行整合[28],插入的外源基因破坏了amyE基因,宿主菌不能表达淀粉酶,从而可以通过功能平板和酶活测定来鉴定阳性重组子。另外,一些其他的整合型载体还可通过thrC位点进行整合,插入的外源基因破坏thrC基因,从而导致苏氨酸营养缺陷型[29],以此来筛选重组子(图2)。常见的整合型载体,见表2。
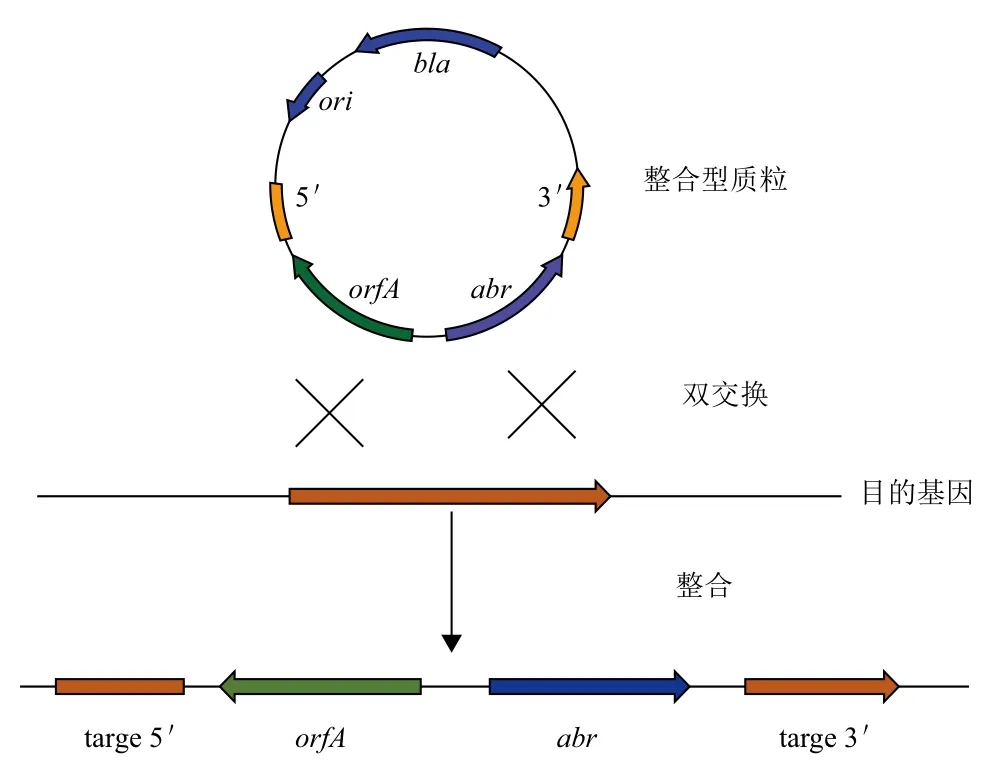
图2 双交换整合型载体
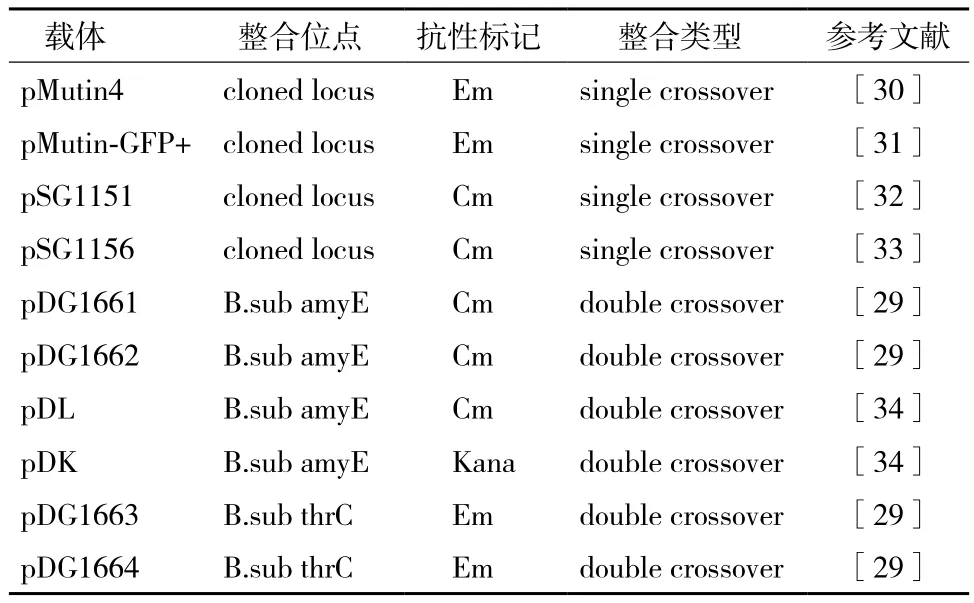
表2 常见的芽孢杆菌整合型载体
3 枯草表达系统的启动子
启动子是位于结构基因5'端上游的DNA序列,能活化RNA聚合酶,使之与模板DNA准确的结合并具有转录起始的特异性。启动子是实现外源基因高效表达的关键。
3.1 启动子的分类
依照控制转录水平的高低,启动子可以分为强启动子和弱启动子;从启动子诱导机制上分类,启动子可以分为组成型启动子、诱导型启动子、时期特异性启动子及自诱导启动子,下面着重介绍枯草芽孢杆菌中已发表的组成型启动子和诱导型启动子[2],并对时期特异性启动子和自诱导启动子做简单的介绍。
3.1.1 组成型启动子 组成型启动子就是不需要任何诱导物,可以持续性表达目的蛋白的启动子。这类启动子启动强度通常要求较强,且成本低,一些组成型启动子常通过构建胞内或分泌表达载体表达目的蛋白。常用的枯草杆菌组成型启动子有P43启动子和噬菌体启动子,其中P43启动子是枯草杆芽孢菌胞苷脱氨酶(cdd)基因的启动子,广泛用于研究,该启动子属于重叠启动子,分别由σA和σB识别,前者负责转录持家基因和孢子形成早期基因,后者一般负责与胁迫相关的基因[35]。研究发现P43启动子为强启动子,启动强度强于诱导型启动子PsacB和PamyE,普遍作为表达载体的调控元件[36]。Yang等[37]通过启动子诱捕系统(Promoter trap system)从地衣芽孢杆菌基因组中筛选到一个强启动子Pshuttle-09,其强度是P43的8倍,在分析Pshuttle-09序列的基础上,Yang等又构建了一个人工双启动子Plaps,此启动子的强度是P43的13倍,是目前应用在枯草芽孢杆菌中表达的最强的组成型启动子。不过组成型启动子也存在一些问题,如组成型表达载体不稳定,表达量不易控制,不适合表达对宿主菌有害的或有毒性蛋白,另外重复使用同一种启动子表达多个外源基因可能会引起基因沉默或共抑制现象等。
3.1.2 诱导型启动子 与组成型启动子相比,诱导型启动子可根据需要在特定的诱导条件,快速诱导基因转录的“开”与“关”。诱导型启动子可以分为两类:一类与环境应答有关,如渗透压、pH值、氧气匮乏、低温、热休克等,在这些环境下,启动子启动相应基因转录,以适应环境变化;另一类是化学诱导启动子,即受化学诱导物诱导而提高相应基因的表达量。下面主要介绍枯草芽孢杆菌中常用的几个化学物诱导启动子。
枯草芽孢杆菌中最常见的是受IPTG诱导的启动子,IPTG能够与大肠杆菌的转录阻遏蛋白LacI结合,使得LacI从操纵位点上脱离,从而诱导下游基因的表达。最早应用的是启动子Pspac,由枯草芽孢杆菌噬菌体SPO1和大肠杆菌lac操纵子融合而成[38]。随后Phan等[39]将热激蛋白GroES和GroEL编码序列的强启动子与lac操纵子融合,构成受IPTG诱导的启动子Pgrac。Phan等[40]进一步通过优化启动子Pgrac的调控元件,构建另一个更强的启动子Pgrac100。虽然受IPTG诱导的启动子强并且可控,但它存在以下3个缺点:(1)IPTG价格昂贵,且有毒性,不适宜进行大规模的外源蛋白的发酵生产;(2)Pspac启动强度仍然不够,外源蛋白的表达量不是很高;(3)Pspac不属于严紧的可控启动子,从而导致在未加入诱导物IPTG的情况下,仍然有少量的重组蛋白表达。因而这些由IPTG诱导的启动子不适用于工业上大规模蛋白酶的生产。
枯草芽孢杆菌中第2个启动子系统是以木糖为诱导物的启动子Pxyl。Pxyl由xylR编码的阻遏蛋白严格控制表达,通过添加0.1%-2%的木糖,能够诱导该启动子的表达,木糖启动子系统对代谢中的大部分代谢产物不敏感,不过却受到葡萄糖的抑制[41]。
枯草芽孢杆菌中第3个启动子系统是以蔗糖为诱导物的sacB启动子,sacB基因是枯草芽孢杆菌基因组上受蔗糖诱导调控的基因,编码外分泌的蔗糖果聚糖酶(Levansucrase)。其启动子sacR转录起始是持续性的,不受蔗糖诱导的控制,但在无蔗糖的情况下,其表达强度比有蔗糖诱导时低100倍[42]。
这3个启动子诱导系统在枯草芽孢杆菌中应用广泛,尤其是前两个系统,常被用做外源蛋白高效表达的元件,但它们共同存在一个缺点就是所用的诱导物IPTG和木糖价钱昂贵,在工业生产中易增加成本,因此限制了在工业生产中的应用。而蔗糖诱导启动子的启动强度相对较弱,无法在工业生产中高效表达目标蛋白。
除了上述介绍的3种诱导启动子外,枯草芽孢杆菌中还有受淀粉诱导的持续性表达的淀粉酶基因启动子Pamy[36],以及受磷酸[43]、柠檬酸盐[44]、四环素[45]、枯草菌素[46]、细胞壁抗生素[47]和甘氨酸[48]等诱导的启动子。此外,为了降低诱导物的成本以更利于工业化应用,Heravi等[49]开发了以甘露醇为诱导物的表达系统。Yang等[14,50]利用麦芽糖为诱导物的启动子Pglv构建了表达系统。Reza等[51]通过施加环境压力和葡萄糖匮乏构建了PohrB表达系统。
3.1.3 时期特异性启动子 转录组学分析显示基因表达是有时期依赖性的[52]。一些基因特异性的在对数生长期表达,而另外一些基因却在菌体进入稳定期后才被激活表达。应用这些时期特异性表达基因的调控元件来表达外源基因,可以避免添加外源的诱导物。目前有两个此类启动子被用于外源基因的表达。一个是启动子PrpsF,特异性在对数期表达,利用此启动子已成功地将来源于C. perfringens的β-类毒素的基因进行了高效表达[53]。另一个使用的启动子是在对数末期表达的PaprE。PaprE是σA依赖型的启动子,且它的活性被高度控制。使用lacZ作为报告基因,将其与启动子共同构建在枯草杆菌整合型载体上,整合进枯草杆菌基因组进行外源蛋白的表达,结果发现β-半乳糖苷酶的表达量约占总蛋白量的10%[54]。
3.1.4 自诱导启动子 自诱导启动子在工业生产中具有优势。Lee等[55]运用苏云金芽胞杆菌伴孢晶体蛋白基因cry3Aa的启动子Pcry3Aa构建了枯草芽孢杆菌稳定期自诱导表达系统,并且通过对此启动子进行优化,优化后外源蛋白lacZ的表达量较优化前提高了5倍。优化后的Pcry3Aa启动子可能会成为枯草芽孢杆菌表达系统的一个强的备选启动子。另外,Wenzel等[56]利用启动子PmanP构建了适合高细胞密度发酵的自诱导系统。这种新型的枯草芽孢杆菌自诱导系统在工业生产上将不仅可以提高外源蛋白的产量,还可以降低生产成本。
综上所述,枯草芽孢杆菌表达系统常用的启动子,见表3。

表3 枯草芽孢杆菌表达系统常用的启动子
3.2 启动子克隆的方法
启动子的研究一方面是为了得到有利于实际应用的高强度启动子;另一方面也有助于进一步了解基因组和遗传特性。目前克隆启动子的方法多种多样,大致可分为两种:一是用启动子诱捕系统(即用到启动子探针质粒载体)筛选启动子;另一种根据基因组信息结合计算机软件预测直接用PCR扩增已知的基因启动子,然后再进行后期生物实验验证,这种方法精确性较差。因此,对于筛选到的启动子还需要进行进一步的优化。
3.2.1 利用启动子诱捕系统筛选启动子 利用启动子诱捕系统筛选启动子是一种简单有效且更加准确的方法[37,57]。启动子诱捕系统中最为核心的部分是构建启动子探针质粒载体,这种载体通常由不含启动子成分的报告基因和一个筛选标记基因组成,只有当内源基因编码区插入到载体上,序列中产生融合转录本时报告基因才可能表达。在探针载体构建时,报告基因的选择至关重要。作为报告基因,它的全序列必须是已知的、已克隆研究过,另外报告基因的表达产物在宿主菌中不存在背景干扰,并且其表达产物的检测应简单、快捷、灵敏度高、重复性好。目前常用的报告基因有以下几种:(1)β-半乳糖苷酶基因:细菌β-半乳糖苷酶基因,尤其是来自于嗜热芽孢杆菌的耐热性的β-半乳糖苷酶基因被广泛应用,它能水解底物X-gal,利用蓝白斑筛选可鉴定插入启动子的阳性克隆。Yang等[37]利用来自于嗜热芽孢杆菌的bgaB基因为报告基因构建启动子捕获载体pShuttleF,从地衣芽孢杆菌的基因组中筛选出比P43表达强度强8倍的PShuttle-09启动子;(2)氯霉素转乙酰酶报告基因(CAT) :细菌CAT酶可以将乙酰基从乙酰辅酶A转移到氯霉素的3-羟基位上,而使宿主菌失去抗性,再通过检测放射性标记的底物实现量化。以CAT为报告基因检测简单,重现性好且灵敏度高。Harwood等[58]以氯霉素抗性基因为报告基因构建启动子捕获载体,在枯草芽孢杆菌中克隆了一些启动子片段。(3)绿色荧光蛋白报告基因(gfp) :绿色荧光蛋白基因最初是从维多利亚发光水母(Aequorea victoria)中分离得到,在508 nm处自行发射绿色荧光,无需辅助因子和底物。GFP相对分子质量小、对活细胞无毒害、荧光稳定且检测方法简单直观(通过荧光显微镜或流式细胞仪检测),加热、变性剂及一般的蛋白酶均不能使GFP失活。GFP上述的优点使其成为报告基因的后起之秀。目前已经有多种GFP突变体,如蓝色荧光蛋白(CFP)、黄色荧光蛋白(YFP)及加强型的荧光蛋白(EGFP)等。Gat等[57]以gfp和egfp基因为报告基因,通过启动子诱捕载体从炭疽芽孢杆菌(Bacillus anthracis)中筛选出用于重组蛋白rPA表达的启动子。此外,还有β-葡萄糖苷酸酶(β-glucuronidase)基因、酸性磷酸脂酶基因及荧光素酶基因等,在启动子筛选中也应用广泛。
3.2.2 结合生物信息学和PCR技术克隆启动子 应用启动子诱捕载体克隆启动子无需知道基因的具体序列,可随机筛选启动子,这样不用设计引物,可得到大量的启动子片段。但这种方法需要构建启动子文库,筛选工作量大。随着生物信息学和PCR技术的发展,利用计算机软件预测加上PCR扩增和后期的实验验证,从基因组中分离已知序列的启动子逐渐成为热门。
Wang等[59]在苏云金芽胞杆菌的RNA测序结果的基础上,运用生物信息学预测未知基因的转录起始位点,并将转录起始位点上游500 bp的核苷酸进行分析,寻找与之匹配的σ因子和转录因子的识别位点,从而判断是否存在潜在的可开发的启动子。使用PCR技术将选定的启动子克隆并构建到表达载体上,后期进一步通过实验验证这些选定的启动子强度。在此基础上,他们筛选出20个不同时期表达的启动子,为苏云金芽胞杆菌的分子生物学的基础研究及应用研究奠定了基石。
除了上述的普通PCR扩增启动子外,还有常用反向PCR、衔接头PCR及TAIL-PCR等方法扩增启动子序列。
4 展望
目前,已有许多的原核基因和真核基因在枯草芽孢杆菌中表达。已克隆的原核基因很多,如属于分子遗传学范畴的一些与质粒构建有关的抗性基因;与DNA结构有关的DNA重组、诱变、修复等相关基因;与基因表达及其调控有关的Sigma因子、Trna、正负调控基因等;与芽孢形成萌发的spoOA、spoIIG基因等;以及与苏云金芽胞杆菌伴胞晶体蛋白相关的cry、cyt基因等;甚至有些与感受态、蛋白分泌有关的Com、SecDF基因等。此外,还有一些工业生产用酶也成功在枯草芽孢杆菌中表达,如碱性蛋白酶、中性蛋白酶A和B、α-淀粉酶、β-淀粉酶、支链淀粉酶、木聚糖酶、脂肪酶、β-半乳糖苷酶和真枯草多肽酶F等。
真核基因在原核生物中表达总是不理想,其主要原因一是原核生物中缺乏蛋白翻译后修饰如糖基化等问题;二是枯草杆菌的胞外蛋白酶易降解真核基因产物。因此,其缺失蛋白酶基因的枯草芽孢杆菌成为高表达外源蛋白的理想菌株。
基因表达体系是一个复杂的过程,与大肠杆菌表达系统比较,枯草表达系统还不是很成熟。近年来,科研工作者在枯草芽孢杆菌分泌表达外源蛋白方面取得很大的进展,已建立一个有效的外源蛋白枯草表达系统。相比于传统的大肠杆菌系统,枯草表达系统要远远落后于它,但在蛋白表达纯化和分泌活性蛋白方面则要明显优于大肠杆菌。实践证明,枯草表达系统可以表达多种可溶性的并具有生物活性的蛋白质。在进一步发展和完善枯草表达系统过程中,势必要不断完善和扩大枯草芽孢杆菌载体系统和宿主系统,而在载体系统的改造过程中,启动子调控元件起着重中之重的作用。因此,从启动子出发,筛选新的强启动子元件,构建成熟高效的枯草表达系统,无论是在理论研究还是实际生产中都有十分重要的意义。
[1]Harwood CR, Wipat A. Sequencing and functional analysis of thegenome of Bacillus subtilis strain 168 [J]. FEBS Lett, 1996, 389(1):84-87.
[2]张晓舟. 枯草杆菌新型表达系统和遗传操作体系的建立及应用[D]. 南京:南京农业大学, 2006.
[3]Spizizen J. Transformation of biochemically deficient strains of Bacillus subtilis by deoxyribonucleate [J]. Proceedings of the National Academy of Sciences of the United States of America, 1958,44(10):1072.
[4]Haldenwang WG. The sigma factors of Bacillus subtilis [J]. Microbiological Reviews, 1995, 59(1):1-30.
[5]Tjalsma H, Bolhuis A, Jongbloed JD, et al. Signal peptide-dependent protein transport in Bacillus subtilis:a genome-based survey of the secretome [J]. Microbiology and Molecular Biology Reviews, 2000,64(3):515-547.
[6]Bendtsen JD, Kiemer L, Fausbøll A, et al. Non-classical protein secretion in bacteria [J]. BMC Microbiology, 2005, 5(1):58.
[7]Tjalsma H, Antelmann H, Jongbloed JD, et al. Proteomics of protein secretion by Bacillus subtilis:separating the “secrets” of the secretome [J]. Microbiology and Molecular Biology Reviews, 2004,68(2):207-233.
[8]Fu LL, Xu ZR, Li WF, et al. Protein secretion pathways in Bacillus subtilis:Implication for optimization of heterologous protein secretion [J]. Biotechnology Advances, 2007, 25(1):1-12.
[9]Banerjee S, Hansen JN. Structure and expression of a gene encoding the precursor of subtilin, a small protein antibiotic [J]. Journal of Biological Chemistry, 1988, 263(19):9508-9514.
[10]Chung Y, Breidt F, Dubnau D. Cell surface localization and processing of the ComG proteins, required for DNA binding during transformation of Bacillus subtilis [J]. Molecular Microbiology,1998, 29(3):905-913.
[11]Wu XC, Lee W, Tran L, et al. Engineering a Bacillus subtilis expression-secretion system with a strain deficient in six extracellular proteases [J]. Journal of Bacteriology, 1991, 173(16):4952-4958.
[12] Murashima K, Chen CL, Kosugi A, et al. Heterologous production of Clostridium cellulovorans engB, using protease-deficient Bacillus subtilis, and preparation of active recombinant cellulosomes[J]. Journal of Bacteriology, 2002, 184(1):76-81.
[13] Plasmids SB, Harwood CR, Cutting SM(eds)//Molecular biological methods for Bacillus [J]. New York:John Wiley and Sons, 1990:75-174.
[14] Yang MM, Zhang WW, Zhang XF, et al. Construction and characterization of a novel maltose inducible expression vector in Bacillus subtilis [J]. Biotechnology Letters, 2006, 28(21):1713-1718.
[15] Kunst F, Ogasawara N, Moszer I, et al. The complete genome sequence of the gram-positive bacterium Bacillus subtilis [J]. Nature, 1997, 390(6657):249-256.
[16] Lacey R, Chopra I. Genetic studies of a multi-resistant strain of Staphylococcus aureus [J]. Journal of Medical Microbiology, 1974,7(2):285-297.
[17]Iordanescu S, Surdeanu M, Della-Latta P, et al. Incompatibility and molecular relationships between small staphylococcal plasmids carrying the same resistance marker [J]. Plasmid, 1978, 1(4):468-479.
[18]Iordănescu S. Three distinct plasmids originating in the same Staphylococcus aureus strain [J]. Archives Roumaines de Pathologie Expérimentales et de Microbiologie, 1976, 35(1-2):111.
[19]Hofemeister J, Israeli-Reches M, Dubnau D. Integration of plasmid pE194 at multiple sites on the Bacillus subtilis chromosome [J]. Molecular and General Genetics MGG, 1983, 189(1):58-68.
[20]Bron S, Meijer W, Holsappel S, et al. Plasmid instability and molecular cloning in Bacillus subtilis [J]. Research in Microbiology, 1991, 142(7):875-883.
[21]Bron S, Luxen E, Swart P. Instability of recombinant pUB110 plasmids in Bacillus subtilis:plasmid-encoded stability function and effects of DNA inserts [J]. Plasmid, 1988, 19(3):231-241.
[22]Nagarajan V, Albertson H, Chen M, et al. Modular expression and secretion vectors for Bacillus subtilis [J]. Gene, 1992, 114(1):121-126.
[23]Wang LF, Wong SL, Lee SG, et al. Expression and secretion of human atrial natriuretic α-factor in Bacillus subtilis using the subtilisin signal peptide [J]. Gene, 1988, 69(1):39-47.
[24]Wu SC, Wong SL. Development of improved pUB110-based vectors for expression and secretion studies in Bacillus subtilis [J]. Journal of Biotechnology, 1999, 72(3):185-195.
[25]Melnikov A, Youngman PJ. Random mutagenesis by recombinational capture of PCR products in Bacillus subtilis and Acinetobactercalcoaceticus [J]. Nucleic Acids Research, 1999, 27(4):1056-1062.
[26]Chédin F, Noirot P, Biaudet V, et al. A five-nucleotide sequence protects DNA from exonucleolytic degradation by AddAB, the RecBCD analogue of Bacillus subtilis [J]. Molecular Microbiology,1998, 29(6):1369-1377.
[27]Chedin F, Ehrlich SD, Kowalczykowski SC. The Bacillus subtilis AddAB helicase/nuclease is regulated by its cognate Chi sequence in vitro [J]. Journal of Molecular Biology, 2000, 298(1):7-20.
[28]Hidenori S, Henner DJ. Construction of a single-copy integration vector and its use in analysis of regulation of the trp operon of Bacillus subtilis [J]. Gene, 1986, 43(1):85-94.
[29]Guérout-Fleury AM, Frandsen N, Stragier P. Plasmids for ectopic integration in Bacillus subtilis [J]. Gene, 1996, 180(1):57-61.
[30]Vagner V, Dervyn E, Ehrlich SD. A vector for systematic gene inactivation in Bacillus subtilis [J]. Microbiology, 1998, 144(11):3097-3104.
[31]Kaltwasser M, Wiegert T, Schumann W. Construction and application of epitope-and green fluorescent protein-tagging integration vectors for Bacillus subtilis [J]. Applied and Environmental Microbiology, 2002, 68(5):2624-2628.
[32]Feucht A, Lewis PJ. Improved plasmid vectors for the production of multiple fluorescent protein fusions in Bacillus subtilis [J]. Gene,2001, 264(2):289-297.
[33]Lewis PJ, Marston AL. GFP vectors for controlled expression and dual labelling of protein fusions in Bacillus subtilis [J]. Gene,1999, 227(1):101-109.
[34]Yuan G, Wong SL. Regulation of groE expression in Bacillus subtilis:the involvement of the sigma A-like promoter and the roles of the inverted repeat sequence(CIRCE) [J]. Journal of Bacteriology, 1995, 177(19):5427-5433.
[35]Zhang XZ, Cui ZL, Hong Q, et al. High-level expression and secretion of methyl parathion hydrolase in Bacillus subtilis WB800[J]. Applied and Environmental Microbiology, 2005, 71(7):4101-4103.
[36]Ye RQ, Kim JH, Kim BG, et al. High-level secretory production of intact, biologically active staphylokinase from Bacillus subtilis [J]. Biotechnology and Bioengineering, 1999, 62(1):87-96.
[37]Yang MM, Zhang W, Ji SY, et al. Generation of an artificial double promoter for protein expression in Bacillus subtilis through a promoter trap system [J]. PLoS One, 2013, 8(2):9.
[38]Yansura DG, Henner DJ. Use of the Escherichia coli lac repressor and operator to control gene expression in Bacillus subtilis [J]. Proceedings of the National Academy of Sciences, 1984, 81(2):439-443.
[39]Phan TTP, Nguyen HD, Schumann W. Novel plasmid-based expression vectors for intra-and extracellular production of recombinant proteins in Bacillus subtilis [J]. Protein Expression and Purification, 2006, 46(2):189-195.
[40]Phan TTP, Nguyen HD, Schumann W. Development of a strong intracellular expression system for Bacillus subtilis by optimizing promoter elements [J]. Journal of Biotechnology, 2012, 157(1):167-172.
[41]Kim L, Mogk A, Schumann W. A xylose-inducible Bacillus subtilis integration vector and its application [J]. Gene, 1996, 181(1):71-76.
[42]Zukowski MM, Miller L. Hyperproduction of an intracellular heterologous protein in a sacU sup hsup mutant of Bacillus subtilis[J]. Gene, 1986, 46(2):247-255.
[43]Lee JK, Edwards CW, Hulett FM. Bacillus licheniformis APase I gene promoter:a strong well-regulated promoter in B. subtilis [J]. Journal of General Microbiology, 1991, 137(5):1127-1133.
[44]Yamamoto H, Murata M, Sekiguchi J. The CitST two-component system regulates the expression of the Mg-citrate transporter in Bacillus subtilis [J]. Molecular Microbiology, 2000, 37(4):898-912.
[45]Geissendörfer M, Hillen W. Regulated expression of heterologous genes in Bacillus subtilis using the Tn10 encodedtet regulatory elements [J]. Applied Microbiology and Biotechnology, 1990, 33(6):657-663.
[46]Bongers RS, Veening JW, Van Wieringen M, et al. Development and characterization of a subtilin-regulated expression system in Bacillus subtilis:strict control of gene expression by addition of subtilin [J]. Applied and Environmental Microbiology, 2005, 71(12):8818-8824.
[47]Toymentseva AA, Schrecke K, Sharipova MR, et al. The LIKE system, a novel protein expression toolbox for Bacillus subtilis based on the liaI promoter [J]. Microbial Cell Factories, 2012, 11(1):143.
[48]Phan TTP, Schumann W. Development of a glycine-inducible exp-ression system for Bacillus subtilis [J]. Journal of Biotechnology,2007, 128(3):486-499.
[49]Heravi KM, Wenzel M, Altenbuchner J. Regulation of mtl operon promoter of Bacillus subtilis:requirements of its use in expression vectors [J]. Microbial Cell Factories, 2011, 10:83.
[50]Yang MM, Zhang WW, Chen YL, et al. Development of a Bacillus subtilis expression system using the improved Pglv promoter [J]. Microbial Cell Factories, 2010, 9(1):55.
[51]Panahi R, Vasheghani-Farahani E, Shojaosadati SA, et al. Induction of Bacillus subtilis expression system using environmental stresses and glucose starvation [J]. Annals of Microbiology, 2014, 64(2):879-882.
[52]Blom EJ, Ridder AN, Lulko AT, et al. Time-resolved transcriptomics and bioinformatic analyses reveal intrinsic stress responses during batch culture of Bacillus subtilis [J]. PLoS One, 2011, 6(11):e27160.
[53]Nijland R, Lindner C, Van Hartskamp M, et al. Heterologous production and secretion of Clostridium perfringens β-toxoid in closely related Gram-positive hosts [J]. Journal of Biotechnology,2007, 127(3):361-372.
[54]Jan J, Valle F, Bolivar F, et al. Construction of protein overproducer strains in Bacillus subtilis by an integrative approach [J]. Applied Microbiology and Biotechnology, 2001, 55(1):69-75.
[55]Lee SJ, Pan JG, Park SH, et al. Development of a stationary phasespecific autoinducible expression system in Bacillus subtilis [J]. Journal of Biotechnology, 2010, 149(1):16-20.
[56]Wenzel M, Müller A, Siemann-Herzberg M, et al. Self-inducible Bacillus subtilis expression system for reliable and inexpensive protein production by high-cell-density fermentation [J]. Applied and Environmental Microbiology, 2011, 77(18):6419-6425.
[57]Gat O, Inbar I, Aloni-Grinstein R, et al. Use of a promoter trap system in Bacillus anthracis and Bacillus subtilis for the development of recombinant protective antigen-based vaccines [J]. Infection and Immunity, 2003, 71(2):801-813.
[58]Harwood CR, Williams DM, Lovett PS. Nucleotide sequence of a Bacillus pumilus gene specifying chloramphenicol acetyltransferase[J]. Gene, 1983, 24(2):163-169.
[59]Wang J, Ai X, Mei H, et al. High-throughput identification of promoters and screening of highly active promoter-5’-UTR DNA region with different characteristics from Bacillus thuringiensis [J]. PLoS One, 2013, 8(5):e62960.
(责任编辑 狄艳红)
Research Progress of Bacillus subtilis Expression System and Its Promoter Regulatory Elements
Yu Xiaoxia Tian Jian Liu Xiaoqing Wu Ningfeng
(Institute of Biotechnology, Chinese Academy of Agricultural Sciences, Beijing 100081)
As a Gram-positive bacteria, Bacillus subtilis is an attractive host for the production of heterologous secretory proteins for several reasons:it is non-pathogenic and the capable of secreting functional extracellular proteins directly to the culture medium, a great deal of vital information concerning large scale fermentation and production technology. One of the key factors for achieving high-level expression of heterologous proteins is the use of a strong and control promoter. In present, the promoters of Bacillus subtilis can be classified into three categories:constitutive promoters, inducer-specific promoters and autoinducible promoters. This paper described the advantages and disadvantages of Bacillus subtilis expression system and the classification of promoters. At the same time, we summarized the methods of the amplification of new promoters, which provided a foundation for improving Bacillus subtilis expression systems and the industrial production of heterologous proteins .
Bacillus subtilis;expression system;constitutive promoters;inducible promoters
10.13560/j.cnki.biotech.bull.1985.2015.02.005
2014-07-10
国家“863计划”项目(2014AA021303)
余小霞,硕士研究生,研究方向:微生物分子生物学与基因工程;E-mail:yuxiaoxia198921@sina.com
伍宁丰,研究员,研究方向:微生物分子生物学与基因工程;E-mail:wuningfeng@caas.cn
