不同预处理对PEG胁迫下水稻幼苗抗氧化系统的影响
2015-09-10陈美静刘倩雯张宝龙等
陈美静 刘倩雯 张宝龙等
摘要: 以模式植物水稻为材料,分别用蒸馏水、10%PEG、100 mmol/L NaCl、1 mmol/L SA、0.1 mmol/L ABA水溶液对水稻种子进行预处理,并以15%PEG模拟水分胁迫,分别测定PEG胁迫0、2、4、6 d时的抗氧化酶[超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)、过氧化氢酶(catalase,CAT)]活性及丙二醛(MDA)、游离脯氨酸含量。结果显示,SOD、POD活性均随着PEG胁迫时间的延长呈上升趋势,CAT活性则呈先上升后下降的趋势;ABA、SA预处理均能在一定时间内显著提高PEG胁迫水稻SOD、POD、CAT活性,ABA预处理降低MDA含量,但对脯氨酸含量无显著影响。研究结果表明,ABA预处理可作为简单、有效的策略缓解水分对水稻的胁迫作用。
关键词: 水稻;PEG胁迫;预处理;抗氧化酶;过氧化产物
中图分类号: S511.01 文献标志码: A
文章编号:1002-1302(2015)08-0076-03
目前世界上的水稻种植面积达到了1.3亿hm2,其中亚洲占92%。我国是水稻种植大国,水稻是我国重要的粮食作物之一。水稻对水分的要求极高,而我国处于温带大陆性气候区,水分缺失是重要的限制因子 [1-2]。面对严峻的水资源短缺问题,研究水稻耐水分胁迫的响应机理、提高水稻的抗旱能力迫在眉睫 [3]。 植物受到PEG等非生物胁迫时会产生活性氧自由基(ROS),对细胞膜系统、蛋白质、脂类、核酸等生物大分子具有强烈的破坏作用 [4-5]。预处理是目前常用的提高植物抗逆性的有效方法之一 [6],Seo等曾研究NaCl胁迫下茉莉酸对水稻赤霉素、脱落酸的影响 [7];Cayuela等曾研究在NaCl胁迫下,用NaCl预处理番茄幼苗来提高番茄果实的产量 [8]。因此,预处理作为提高植物抗逆性的有效方法,应引起足够的重视。 本研究以水稻为试验材料,旨在从抗氧化酶、过氧化产物方面来研究当水稻面临PEG模拟胁迫时,预处理是否能够提高其抗旱能力。
1 材料与方法
1.1 材料准备
以模式植物水稻(Oryza sativa L.)为试验材料,品种为辽星一号,由沈阳农业大学稻作研究室提供。
精选水稻种子,分6组进行预处理。(1)对照组:无预处理;(2)蒸馏水组:用蒸馏水浸种;(3)PEG组:用10%PEG溶液浸种;(4)NaCl组:用100 mmol/L NaCl 溶液浸种;(5)SA组:用 1 mmol/L SA 溶液浸种;(6)ABA组:用0.1 mmol/L ABA 溶液浸种。用400 mL溶液(完全浸没种子)于室温下浸种18 h,然后用蒸馏水冲洗3~5次,铺开放在滤纸上阴干、备用。
将6组种子分别于28 ℃恒温箱(黑暗)中浸种24 h,催芽24~36 h,用Hoagland 完全营养液于人工气候箱(昼/夜温度为28 ℃/25 ℃、光照/黑暗时间为16 h/8 h、相对湿度80%、光照度3 000 lx)中培养,每组播种3盆,每天补充营养液,7 d后将6组水稻幼苗分别用含15%PEG的完全营养液模拟水分胁迫 6 d。并在胁迫前(0 d)和胁迫后2、4、6 d进行各项生理指标的测定。
1.2 生理指标的测定方法
超氧化物歧化酶(superoxide dismutase,SOD)活性采用氮蓝四唑(NBT)光化还原法 [9]测定;过氧化物酶(peroxidase,POD)活性采用愈创木酚法 [10]测定;过氧化氢酶(catalase,CAT)活性采用紫外-可见分光光度法 [11]测定;丙二醛(MDA)、游离脯氨酸含量采用张志良等的方法 [12]测定。
1.3 数据的处理
试验数据采用SPSS 20.0、Excel软件相结合进行方差分析(ANOVA)、LSD检测,比较其差异显著性,所有数据指标均重复3次。
2 结果与分析
2.1 不同预处理对PEG胁迫下水稻幼苗抗氧化酶活性的影响
SOD、POD、CAT是清除植物体内活性氧的抗氧化酶,在水分胁迫条件下,抗氧化酶可以降低体内活性氧的积累,从而降低机体受伤害的程度 [13]。
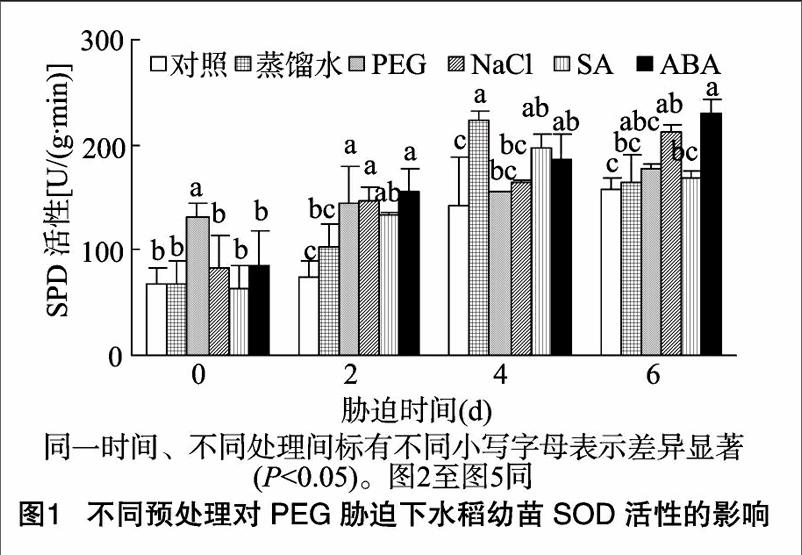
由图1可见,在未进行PEG胁迫时,PEG预处理组SOD活性显著高于对照组及其他预处理组;随着胁迫时间的延长,除蒸馏水、SA预处理组呈先上升后下降趋势外,其他各处理组SOD活性总体呈上升趋势,而自始至终PEG预处理组SOD上升幅度并不明显。胁迫2 d时,除蒸馏水处理组外,各预处理组的SOD活性均显著高于对照组;胁迫4 d时,蒸馏水、SA、ABA处理组的SOD活性显著高于对照组;当胁迫时间达到6 d时,NaCl、ABA预处理组的SOD活性显著高于对照组。这些结果表明,在15%PEG胁迫条件下,预处理激活了水稻的ROS防御机制,使其通过提高SOD活性来保护水稻自身免受ROS的损伤。总体来看,PEG、NaCl、ABA预处理效果较好。
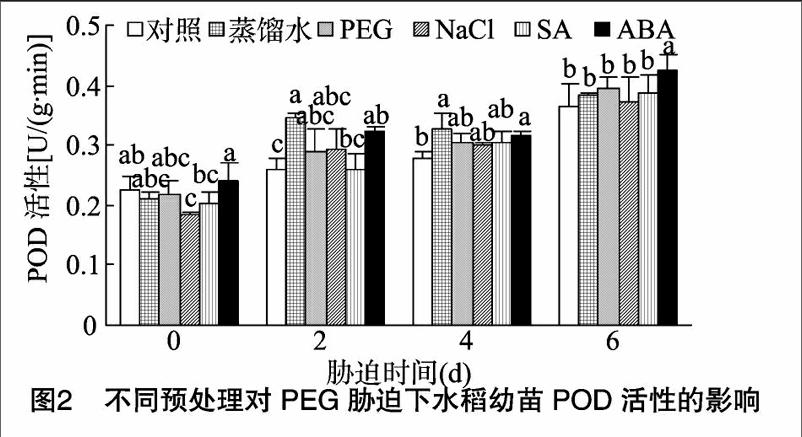
由图2可知,在无胁迫处理时,NaCl预处理POD活性低于对照组,其余各处理组POD活性与对照组间不存在显著差异;随着胁迫时间的延长,各处理组POD活性总体呈上升趋势。PEG胁迫2~4 d时,蒸馏水、ABA处理组POD活性显著高于对照组;当胁迫时间达到6 d时,仅ABA处理组的POD活性显著高于对照组。表明在PEG胁迫下,预处理激活了水稻的ROS防御机制,经过预处理的水稻幼苗抗逆能力更强,特别是蒸馏水、ABA预处理的效果更为明显。
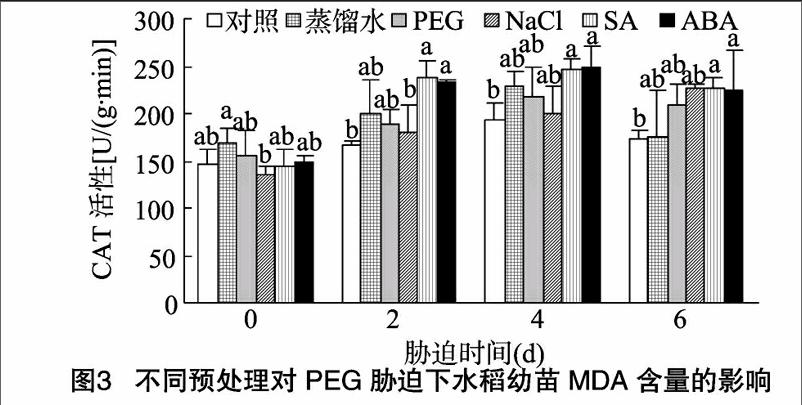
由图3可知,在未受胁迫时,各处理组与对照间CAT活性不存在显著差异;随着胁迫时间的延长,除NaCl处理组CAT活性呈上升趋势外,其他各处理组呈现先上升后下降的趋势。罗英等研究表明,随着PEG胁迫时间的延长,SA预处理的凤仙花CAT活性呈先上升后下降的趋势 [14],本试验结果与之相似。在PEG胁迫2、4、6 d时,各处理组CAT活性变化趋势相似,SA、ABA处理组显著高于对照组,表明在15%PEG胁迫下,预处理激活水稻的ROS防御机制,通过提高CAT活性清除体内过多的过氧化氢来降低自身受伤害的程度,特别是SA、ABA预处理的效果较为明显。
2.2 不同预处理对PEG胁迫下水稻幼苗MDA含量的影响
植物在受PEG胁迫下会产生许多ROS,过多的ROS会使膜脂过氧化,MDA便是膜脂过氧化的产物,会损伤膜系统。由图4可知,在未进行PEG胁迫时,各处理组间MDA含量很低[CM(25]且不存在显著差异;胁迫2 d时,各处理组MDA含量与对照间不存在显著差异;胁迫4 d时,对照组MDA含量急剧升 高,蒸馏水、PEG、ABA预处理组MDA含量显著低于对照组;当胁迫时间达到6 d时,PEG、ABA预处理组的MDA含量显著低于对照组。结果表明,预处理特别是PEG、ABA处理可明显降低MDA含量,从而降低水稻膜脂过氧化的损伤程度。
2.3 不同预处理对PEG胁迫下水稻幼苗游离脯氨酸含量的影响
以PEG模拟水分胁迫会使植物在大量失水时产生渗透胁迫,脯氨酸作为一种有效的渗透调节物质,在植物体内的积累可以调节植物细胞的渗透势 [15]。由图5可知,随着胁迫时间的延长,脯氨酸含量呈上升趋势;在未受胁迫时,NaCl、SA处理的脯氨酸含量较高;随着胁迫时间的延长,对照组脯氨酸含量上升明显,蒸馏水处理组脯氨酸含量上升幅度最小,且含量最低;胁迫处理2 d时,蒸馏水、PEG处理组脯氨酸含量显著低于对照,NaCl、SA、ABA处理组脯氨酸含量与对照无显著差异;胁迫处理4 d时,蒸馏水、PEG、NaCl处理组脯氨酸含量显著低于对照。对照组的脯氨酸含量呈明显上升趋势,表明随着水稻受PEG胁迫程度加深,需要通过增加脯氨酸含量来维持自身水分平衡;胁迫2、4 d,预处理组特别是蒸馏水、PEG处理组的脯氨酸含量明显低于对照组,说明预处理明显提高了水稻的抗性。
3 结论
研究表明,提高植物体内抗氧化酶活性是提高植物抗逆性的有效途径之一 [16-17]。本研究表明,随着PEG胁迫时间的延长,SOD、POD活性呈上升趋势,经过预处理水稻幼苗SOD、POD活性明显高于对照组,表明预处理是一种抗旱锻炼,可提高水稻的抗旱能力;CAT活性多数呈现先上升后下降的趋势。刘晓辉等对西瓜幼苗的研究表明,在外施激素条件下,随着低温胁迫程度的增加,CAT活性呈先上升后下降的趋势 [18]。这可能由于随着PEG胁迫时间的延长,水稻体内积累的ROS过多,CAT来不及清除过多的过氧化氢,导致胁迫后期CAT活性下降 [19]。虽然在胁迫后期,CAT活性有所下降,但是经过SA、ABA预处理的水稻幼苗CAT活性均高于对照组,同样表明了预处理可以提高水稻的抗旱能力。整体来看,SA、ABA预处理对提高15%PEG胁迫下水稻幼苗的抗氧化酶活性效果更为明显,更能提高水稻的抗逆性。
植物在受到PEG等非生物胁迫时,会形成ROS,如果过量的ROS不能及时被清除,植物将会受到严重的氧化伤害 [20]。MDA是植物细胞膜脂过氧化作用的最终产物,对细胞膜具有毒害作用,是最常用的膜脂过氧化指标 [21]。Srivastava 等对芥菜的研究表明,在PEG胁迫下蒸馏水处理的MDA含量要低于对照,而ABA预处理却高于对照 [22]。本试验研究表明,随着PEG胁迫时间的延长,MDA含量呈明显的上升趋势,胁迫4、6 d,对照组MDA含量明显高于其他各预处理组,其中以ABA、PEG处理组的MDA含量最低,表明预处理,特别是PEG、ABA处理明显提高了水稻的抗性。
脯氨酸作为植物体内一种重要的渗透调节物质,在受到PEG胁迫时会大量积累,从而提高其渗透调节能力,对植物起到缓冲保护作用,可提高植物的抗逆性,因此许多人把脯氨酸的积累能力看作是植物抗旱性选择的基础 [23-24]。本试验结果表明,随着PEG胁迫时间的延长,脯氨酸含量呈明显的上升趋势。表明15%PEG引起渗透胁迫,植物体内脯氨酸含量增加以提高渗透调节能力。各预处理组脯氨酸含量明显低于对照组,表明随着胁迫时间的延长,经过预处理的水稻幼苗特别是蒸馏水、PEG预处理组并未感受到强烈的渗透胁迫,不需要大量增加脯氨酸来调节渗透势差,从而明显提高了水稻的抗逆性。
参考文献:
[1] 马廷臣,余蓉蓉,陈荣军,等. PEG-6000模拟干旱对水稻幼苗期根系的影响[J]. 中国生态农业学报,2010,18(6):1206-1211.
[2]戴高兴,彭克勤,萧浪涛,等. 聚乙二醇模拟干旱对耐低钾水稻幼苗丙二醛、脯氨酸含量和超氧化物歧化酶活性的影响[J]. 中国水稻科学,2006,20(5):557-559.
[3]丁 雷,李英瑞,李 勇,等. 梯度干旱胁迫对水稻叶片光合和水分状况的影响[J]. 中国水稻科学,2014,28(1):65-70.
[4]Kanazawa S,Sano S,Koshiba T,et al. Changes in antioxidative enzymes in cucumber cotyledons during natural senescence:comparison with those during dark-induced senescence[J]. Physiologia Plantarum,2000,109:211-216.
[5]Becana M,Dalton D A,Moran J F,et al. Reactive oxygen species and antioxidants in legume nodules[J]. Physiologia Plantarum,2000,109(4):372-381.
[6]Casenave E C,Toselli M E. Hydropriming as a pre-treatment for cotton germination under thermal and water stress conditions[J]. Seed Science and Technology,2007,35(1):88-98.
[7]Seo H S,Kim S K,Jang S W,et al. Effect of jasmonic acid on endogenous gibberellins and abscisic acid in rice under NaCl stress[J]. Biologia Plantarum,2005,49(3):447-450.
[8]Cayuela E,Esta M T,Parra M,et al. NaCl pre-treatment at the seedling stage enhances fruit yield of tomato plants irrigated with salt water[J]. Plant and Soil,2001,230(2):231-238.
[9]Hichem H,Mounir D,Naceur E A. Differential responses of two maize (Zea mays L.) varieties to salt stress:changes on polyphenols composition of foliage and oxidative damages[J]. Industrial Crops and Products,2009,30(1):144-151.
[10]Zhang J X,Kirkham M B. Drought-stress-induced changes in activities of superoxide dismutase,catalase,and peroxidase in wheat species[J]. Plant & Cell Physiology,1994,35(5):785-791.
[11]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:195.
[12]张志良,瞿伟菁,李小方. 植物生理学实验指导[M]. 北京:高等教育出版社,1996.
[13]孙映波,尤 毅,朱根发,等. 干旱胁迫对文心兰抗氧化酶活性和渗透调节物质含量的影响[J]. 生态环境学报,2011,20(11):1675-1680.
[14]罗 英,杨仁强,肖 莲,等. 水杨酸预处理对水分胁迫下凤仙花幼苗抗氧化能力的影响[J]. 江苏农业科学,2010(6):243-245.
[15]周晋军,刘 蓬,尹静静,等. 脯氨酸代谢途径在调控水稻phyB突变体干旱胁迫耐性中的作用[J]. 山东农业科学,2014,46(3):1-4.
[16]杜士云,王德正,吴 爽,等. 三类雄性不育水稻花药和叶片中抗氧化酶活性变化[J]. 植物生理学报,2012,48(12):1179-1186.
[17]Alvarez M E. Salicylic acid in the machinery of hypersensitive cell death and disease resistance[J]. Plant Molecular Biology,2000,44(3):429-442.
[18]刘晓辉,张 显,郑俊鶱,等. 激素预处理对低温胁迫下西瓜幼苗活性氧含量和抗氧化酶活性的影响[J]. 西北植物学报,2014,34(4):746-752.
[19]Dhindsa R S,Matowel W. Drought tolerance in two mosses:correlated with enzymatic defense against lipid peroxidation[J]. J Exp Bot,1981,32(1):79-91.
[20]Bartels D,Sunkar R. Drought and salt tolerance in plants[J]. Crit Rev Plant Sci,2005,24(1):23-58.
[21]赵天宏,孙加伟,赵艺欣,等. CO2和O3浓度升高及其复合作用对玉米(Zea mays L.)活性氧代谢及抗氧化酶活性的影响[J]. 生态学报,2008,28(8):3644-3653.
[22] Srivastava A K,Lokhande V H,Patade V Y,et al. Comparative evaluation of hydro-,chemo-,and hormonal-priming methods for imparting salt and PEG stress tolerance in Indian mustard (Brassica juncea L.)[J]. Acta Physiologiae Plantarum,2010,32(6):1135-1144.
[23] Bohnert H J,Jensen R G. Strategies for engineering water-stress tolerance in plants[J]. Trends in Biotechnology,1996,14(3):89-97.
[24]tajner D,Mimica-Duki c[DD(-1*2][HT6]'[DD)] N,Gasi c[DD(-1*2][HT6]'[DD)] O. Adaptability to drought in sugar beet cultivars[J]. Biologia Plantarum,1995,37(1):107-112.
