辽东湾网采浮游植物粒级结构的胁迫响应
2015-08-30宋广军王年斌赵海勃辽宁省海洋水产科学研究院辽宁大连116023辽宁省海洋环境监测总站辽宁大连116023
宋 伦,宋广军,王年斌,赵海勃,田 金,杨 爽,杜 静(1.辽宁省海洋水产科学研究院,辽宁 大连 116023;2.辽宁省海洋环境监测总站,辽宁 大连 116023)
辽东湾网采浮游植物粒级结构的胁迫响应
宋 伦1,2*,宋广军1,2,王年斌1,2,赵海勃1,2,田 金1,2,杨 爽1,2,杜 静1,2(1.辽宁省海洋水产科学研究院,辽宁 大连 116023;2.辽宁省海洋环境监测总站,辽宁 大连 116023)
基于2012~2014年春、夏、秋、冬季辽东湾网采浮游植物及环境因子的调查资料,对辽东湾南、北2个区域的浮游植物粒级结构分布规律及其环境胁迫响应进行了分析,并评估了浮游植物群落结构的稳定性.结果表明,辽东湾海域浮游植物春、夏、秋季丰度主要集中在小粒径范围,冬季主要集中在中粒径范围;生物量春、夏、秋季主要集中在中粒径范围,冬季主要集中在大粒径范围,南部区域浮游植物粒径整体上大于北部区域.寡营养水域以及较低的水温环境更适于大粒径浮游植物生存.春、秋、冬季浮游植物群落结构比较稳定,但夏季偏离平衡点.南部区域群落结构整体上比北部区域稳定,网采中、大粒径组的W统计量可以作为浮游植物群落结构稳定性的评估指标.
浮游植物;粒级结构;ABC曲线;生态对策;生态稳定性;辽东湾
浮游植物作为海洋初级生产力的主要贡献者,参与物质循环和能量流动,在海洋生态系统中发挥着重要的生态作用.浮游植物粒级差异及演变规律是许多生物和非生物环境因素共同作用的结果[1],浮游植物对营养盐吸收、污染物胁迫等存在一定程度的粒径效应[2-4],比较不同条件下浮游植物的粒级分布对研究生态系统的结构和功能以及探讨不同粒径的进化机制都有非常重要的意义.目前针对浮游植物粒级结构的研究大多数都是采用叶绿素 a分级测定的方法来粗略表示不同粒径生物量结构,但无法了解不同物种的生物量分布.显微测微计法虽然耗时费力,但可进行种间定量描述和深入的作用机理研究.国内学者孙军等[5]首次计算了 87种中国近海常见浮游植物的细胞体积、鲜重、碳含量、氮含量,为后来学者研究长江口[6]、南黄海[7]、大亚湾[1]浮游植物粒级结构及生物量谱奠定了基础.
辽东湾为渤海半封闭式海湾,多年来超负荷承载陆源污染,富营养化加剧,赤潮现象频繁发生[8-9].近些年,辽东湾浮游植物的研究多集中在种群结构及生态分布规律方面[10-15],在粒级结构对环境胁迫响应及稳定性评估方面鲜有研究.宋伦等[16]对辽东湾河口区和港口区浑浊带海域的浮游生物粒径结构进行了研究,发现浮游植物群落对无机氮含量高的浑浊带海域响应可能是通过小粒径生物进行反馈调节的.
本研究选取辽东湾南、北2个区域,采用显微测微计法对浮游植物粒级结构进行了对比分析,研究了群落粒径组成及季节变化规律,并首次利用ABC曲线法探讨了2个区域群落的稳定性状况及对环境胁迫变化响应规律,揭示了富营养海域浮游植物的粒径分布、生物量更替原因及其环境响应机制,可为辽东湾海域赤潮监管和生态修复效果评价提供技术支持.
1 材料与方法
1.1调查区域与样品采集
2012~2014年5月(春)、8月(夏)、10月(秋)、12月(冬,2012年未调查)分别在辽东湾海域进行了网采浮游植物及环境因子调查,于南(S区)、北(N区)2个区域分别设置10个调查站位,其中N区邻近河口,受近岸陆源环境影响较大,S区则相对影响较小(图1).
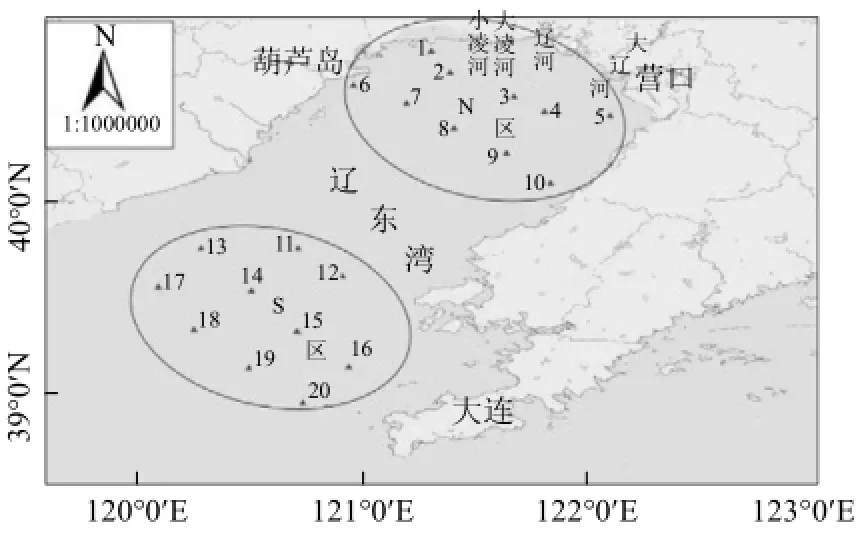
图1 辽东湾采样站位示意Fig.4 Sampling sites in the Liaodong Bay
浮游植物采用《海洋调查规范》[17]推荐的浅水Ⅲ型浮游生物网从底至表层垂直拖网;同期进行海水的水温、盐度、叶绿素a(Chl a.)、无机氮(DIN)[铵盐(NH4+)、硝酸盐(NO3-)、亚硝酸盐(NO2-)之和]、活性磷酸盐(DIP)、石油类(Oil)、pH值、溶解氧(DO)、COD、重金属汞(Hg)、铅(Pb)、铜(Cu)、锌(Zn)、镉(Cd)、砷(As)等指标的监测.调查和监测分析方法均参照《海洋调查规范》(GB/T 12763-2007)和《海洋监测规范》(GB17378-2007)[18]执行.
1.2生物粒级分析与数据处理
实验室内对浮游植物样品进行镜检、计数,并测量其几何参数(对于单个游离细胞,只统计长轴大于 5μm的个体,对于链状细胞,测量总长度后换算平均值,从粒径及营养级角度考虑,本文没有统计夜光藻),对于难以准确鉴定到种的个体,按未定种归类到属记录,根据测定的形态参数、借助数学几何运算公式和体长-含碳量的经验公式分别对浮游植物个体体积和个体含碳量进行估算[5,19-20].浮游植物湿重生物量为细胞体积×1.1,浮游植物粒径大小统一换算成等效球径(ESD)来衡量.
采用相对重要性指数(IRI)确定群落中的优势种:

式中:W为某种类的生物量百分比;N为某种类的丰度百分比;F为调查某种类出现的频率[21].
采用 Shannon-Wiener指数(H′)[16]统计物种多样性水平:
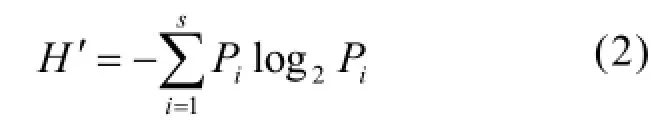
式中:Pi为第i种生物的丰度(或生物量)与样本总丰度(或生物量)的比值;S为种类数.
利用营养指数法(E)并结合CN/CP(无机氮/无机磷)对研究海域的营养状况进行分类[22]:
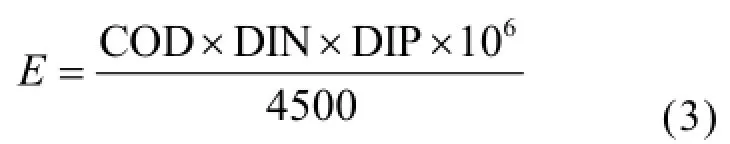
式中:E为富营养化判断值(E),COD、DIN、DIP分别为COD(mg/L)、无机氮(mg/L)、无机磷(mg/L)的测定值.若E≥1,即为富营养化.
环境指数(测定值/标准值)中标准值参照《海水水质标准》[23](GB3097-1997)中的第二类海水水质标准计算.
通过ABC曲线法对群落的稳定性和胁迫变化状况进行分析,用 W统计量(W-statistic)作为ABC曲线方法的一个统计量,其公式为:
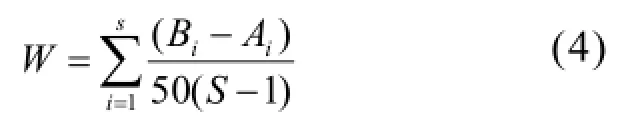
式中:Bi和Ai为ABC曲线中种类序号对应的生物量和丰度的累积百分比,S为出现物种数.当生物量优势度曲线在丰度优势度曲线之上时,W为正,反之W为负[24].
对上述公式计算、数据分析、方差检验、曲线绘制均通过 Microsoft Excel 2007和 SPSS 19.0、PRIMER 5.0软件完成.
2 结果
2.1研究区水质环境特征
调查期间 N 区水温(5.19±0.39)~(25.76± 0.21) ℃,盐度(24.87±1.01)~(27.87±0.41);S区水温(9.99±0.20)~(25.01±0.35)℃、盐度(28.93±0.31)~(30.88±0.15)(图2).N区DIN在春、秋、冬季全部超过第二类海水水质标准,S区水质均在第一类海水水质标准范围内(图3).
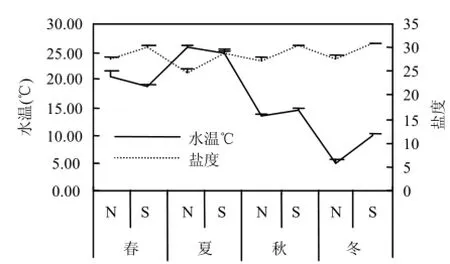
图2 研究区温、盐季节变化Fig.4 Seasonal changes in temperature and salinity in the study area
t检验(成对双样本均值分析)结果表明,N、S区部分水环境指标在调查期间差异较为显著(表 1),综合考虑差值和相关系数的大小,DIN、COD、Cd、Hg、As含量N区较S区高,而S区盐度全年高于N区,冬季水温也高于N区.
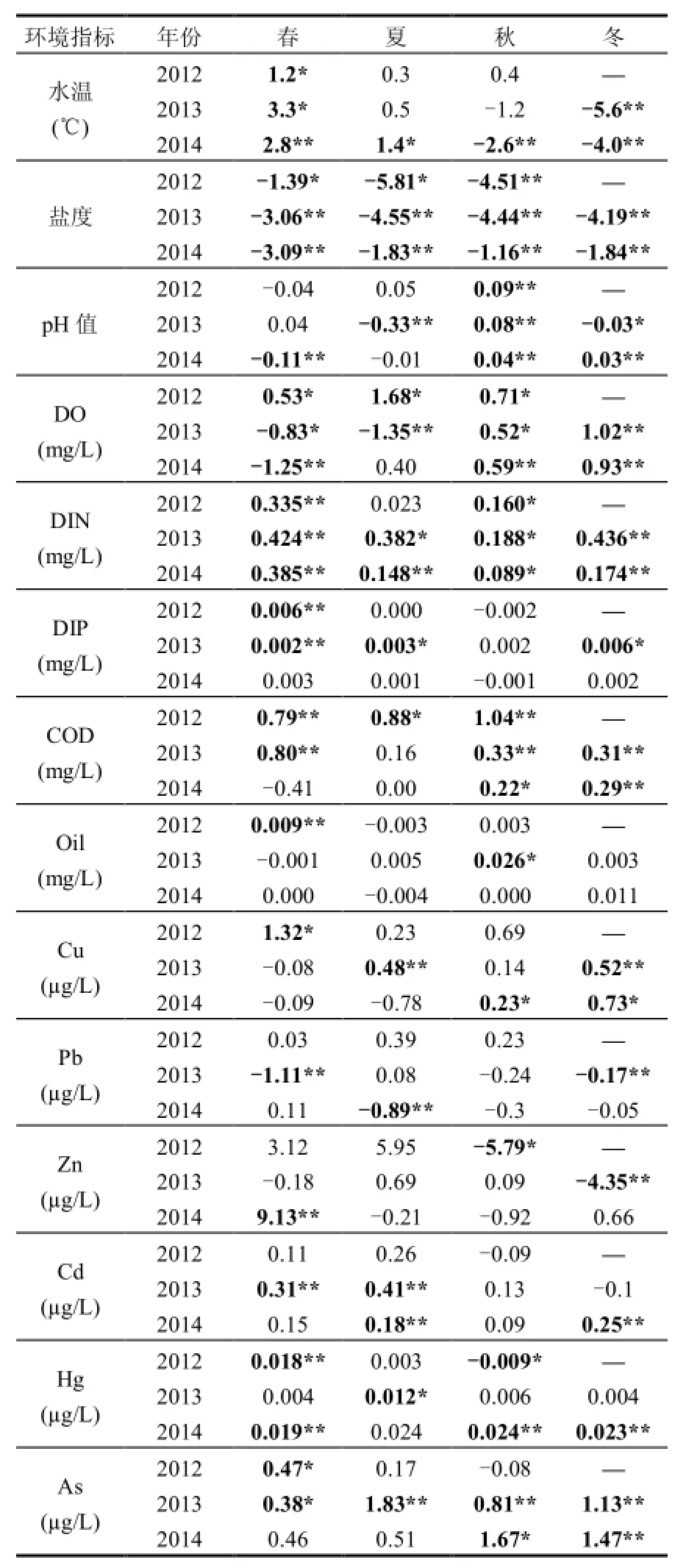
表1 研究区环境因子差值(N-S)Table 1 N-S difference of environmental factors in research area
海水营养状况分析表明,研究N区属于富营养化水平较高海域,S区基本属于低营养水平(表2).这也符合本海域的自然属性,N区北侧邻近辽河、大辽河及大、小凌河,陆域氮源输入丰富.
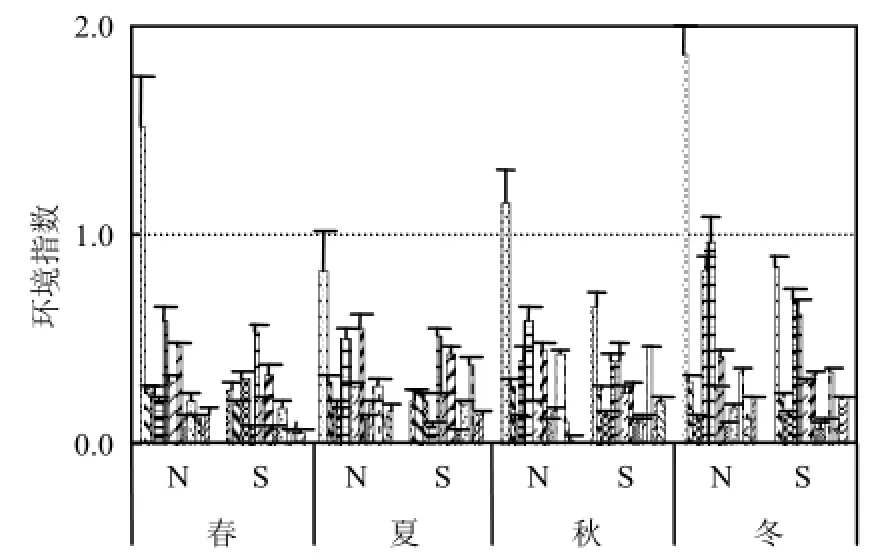
图3 研究区环境因子季节变化Fig.4 Seasonal change of environmental factors in the study area
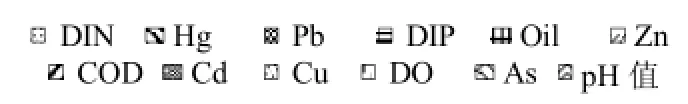
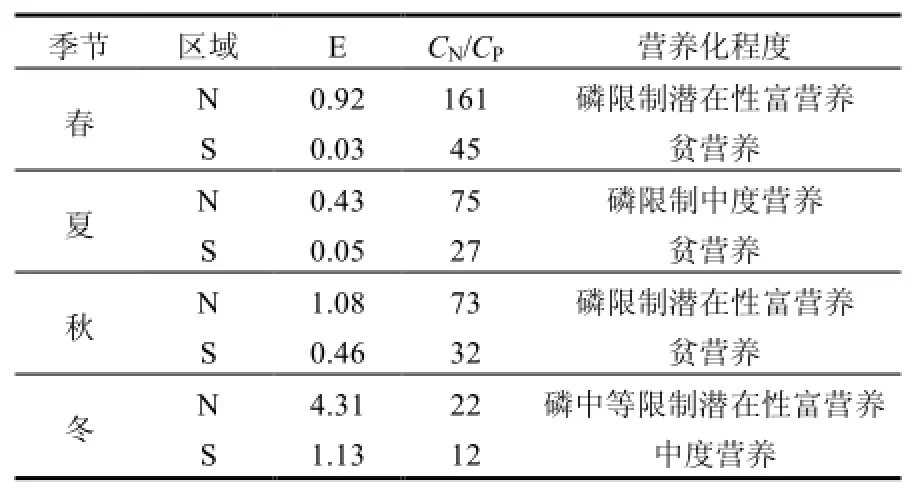
表2 研究区海水营养水平Table 1 Eutrophication level of marine in the study area
2.2优势种及特有种组成
研究区(N、S) ESD<20 μm小粒径组优势种主要由中肋骨条藻(Skeletonema costatum)、具槽帕拉藻(Paralia sulcata)、角毛藻(Chaetoceros sp.)组成,20 μm<ESD<100 μm中粒径组优势种主要由格式圆筛藻(Coscinodiscus granii)、威氏圆筛藻(Coscinodiscus wailesii)组成,ESD>100 μm大粒径组优势种主要由星脐圆筛藻(Coscinodiscus asteromphalus)、虹彩圆筛藻(Coscinodiscus oculus-iridis)组成,N、S区优势种差异不明显(表3).调查还发现,长耳齿状藻(Odontella aurita)、细弱圆筛藻在 N区出现频率较高,而冰河拟星杆藻(Asterionellopsis glacialis)、新月菱形藻(Nitzschia closterium)、 离 心 列 海 链 藻 (Thalassiosira excentrica)、丹麦角毛藻(Chaetoceros danicus)、印度翼根管藻(Rhizosolenia alata f.indica)、蜂窝三角藻(Triceratium favus)、透明辐杆藻(Bacteriastrum hyalinum)、六幅辐裥藻(Actinoptychus senarius)、优美旭氏藻矮小变形(Schroderella delicatula)、掌状冠盖藻(Stephanopyxis palmeriana)、豪猪棘冠藻(Corethron hystrix)、巨圆筛藻(Coscinodiscus gigas)、 海 洋 原 多 甲 藻 (Protoperidinium oceanicum)、长角角藻(Ceratium macroceros)在S区出现频率较高.
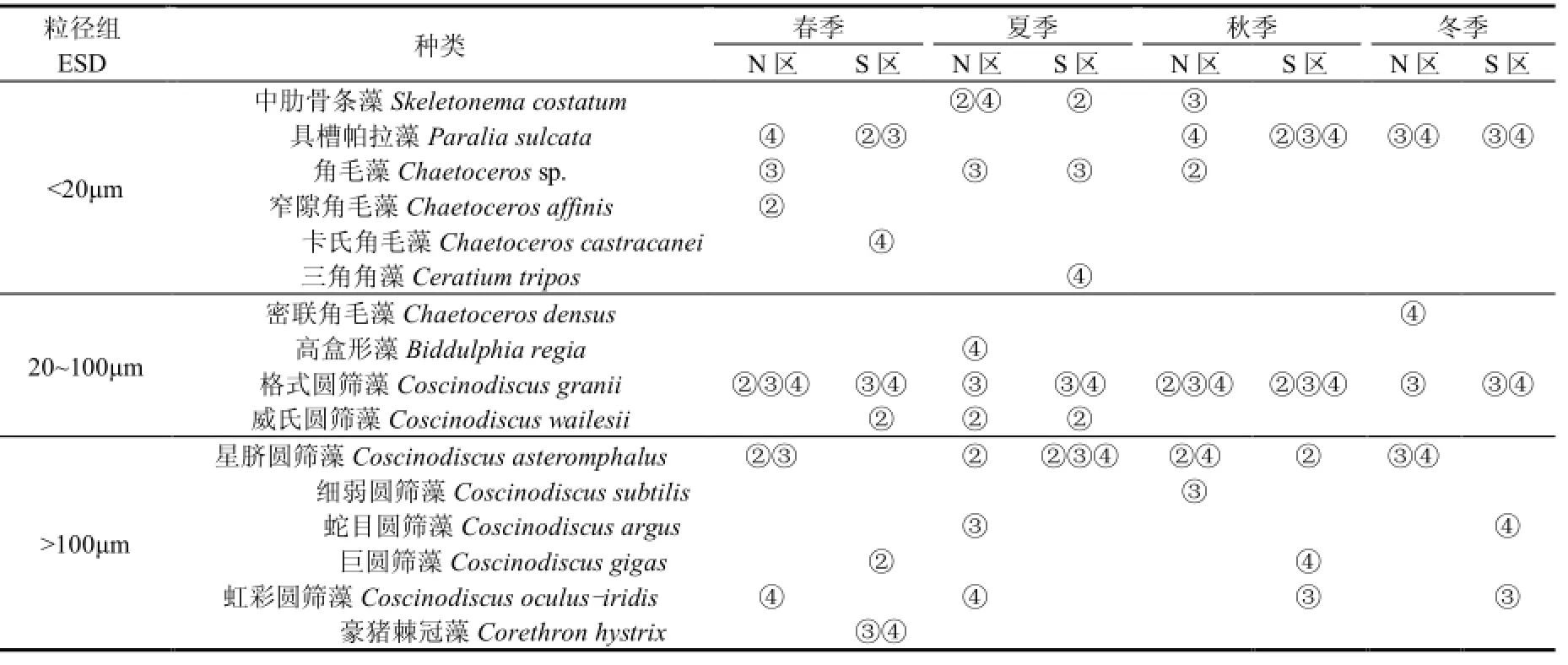
表3 研究区浮游植物各粒径组第一优势种Table 1 Phytoplankton size of the first dominant species in the study area
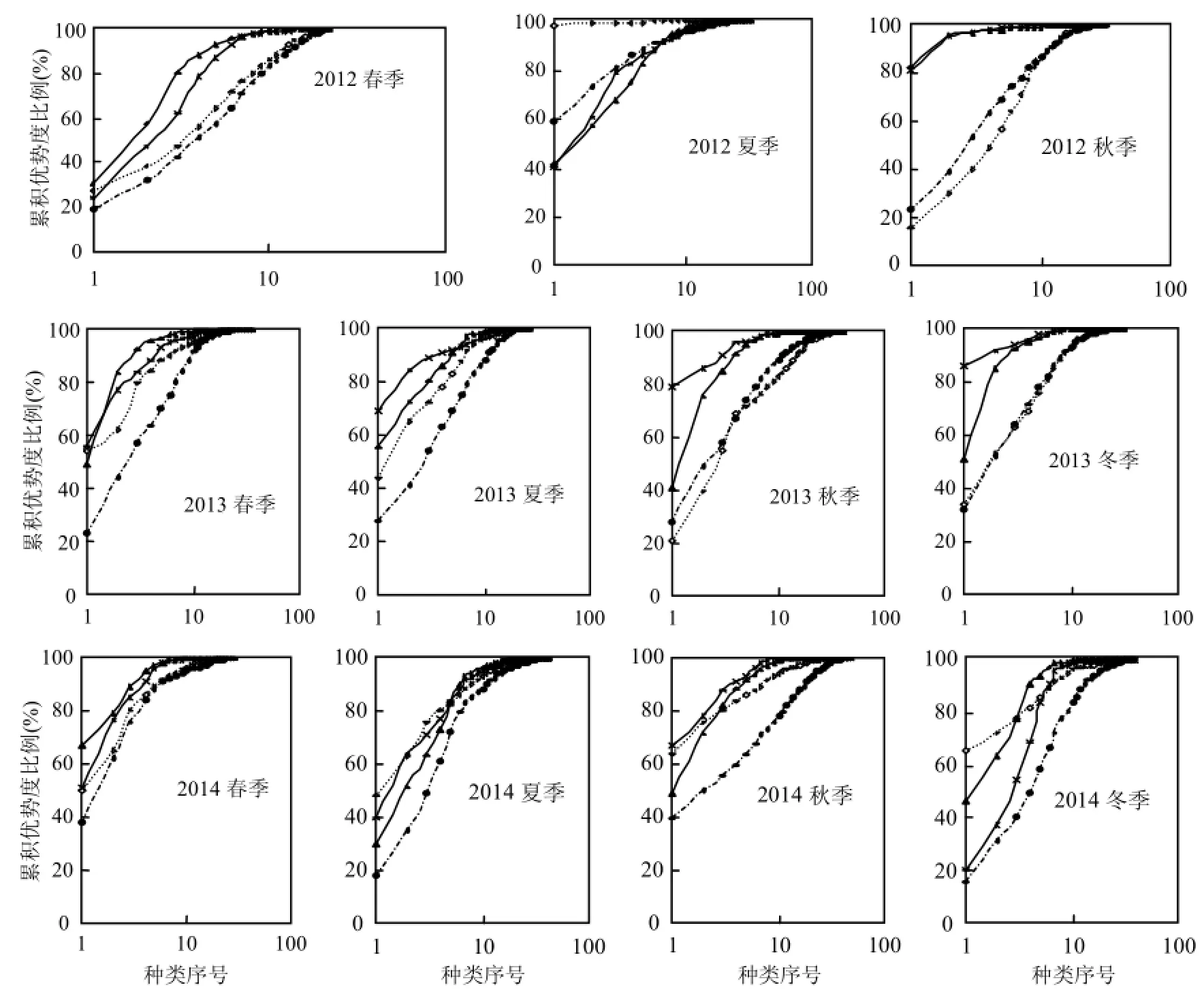
图4 各季节浮游植物群落的ABC曲线以及W统计值Fig.4 ABC curves and W value of phytoplankton community in each season

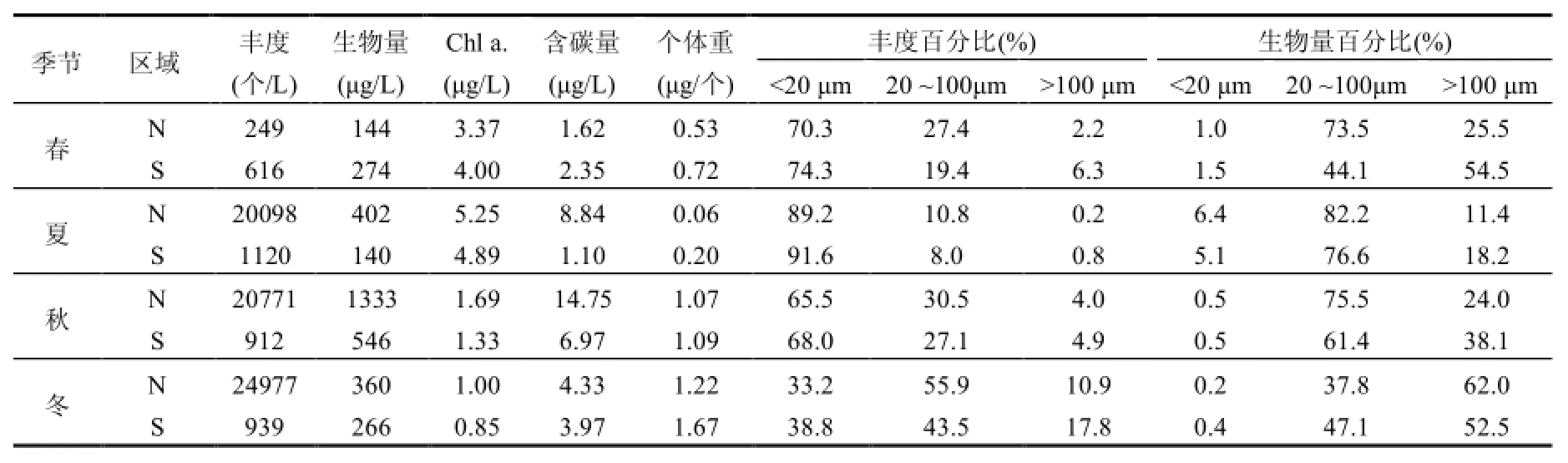
表4 研究区浮游植物粒径组成Table 1 Phytoplankton size composition of the study area
2.3粒级结构变化特征
N、S区ESD全年分布在3~245µm,N区浮游植物总丰度、生物量、Chl a.、含碳量在夏、秋、冬季平均值均高于S区,而浮游植物平均个体重量全年S区均高于N区,表明S区整体上粒径大于N区(表4).从各粒径丰度、生物量组成可见,N、S区春、夏、秋季浮游植物丰度主要集中在 ESD<20μm小粒径区域,冬季主要集中在20μm<ESD<100μm中粒径区域;生物量春、夏、秋季主要集中在20μm<ESD<100μm中粒径区域,冬季主要集中在 ESD>100μm大粒径区域.S区ESD<20μm小粒径组丰度全年高于 N区,而ESD>100μm大粒径组生物量整体高于N区(除冬季外).丰度主要贡献者为中肋骨条藻、具槽帕拉藻、薄壁几内亚藻(Guinardia flaccida)、三角角藻、伏氏海线藻(Thalassionema frauenfeldii)、角毛藻等,而生物量主要贡献者为粒径较大的圆筛藻属.
2.4ABC曲线特征
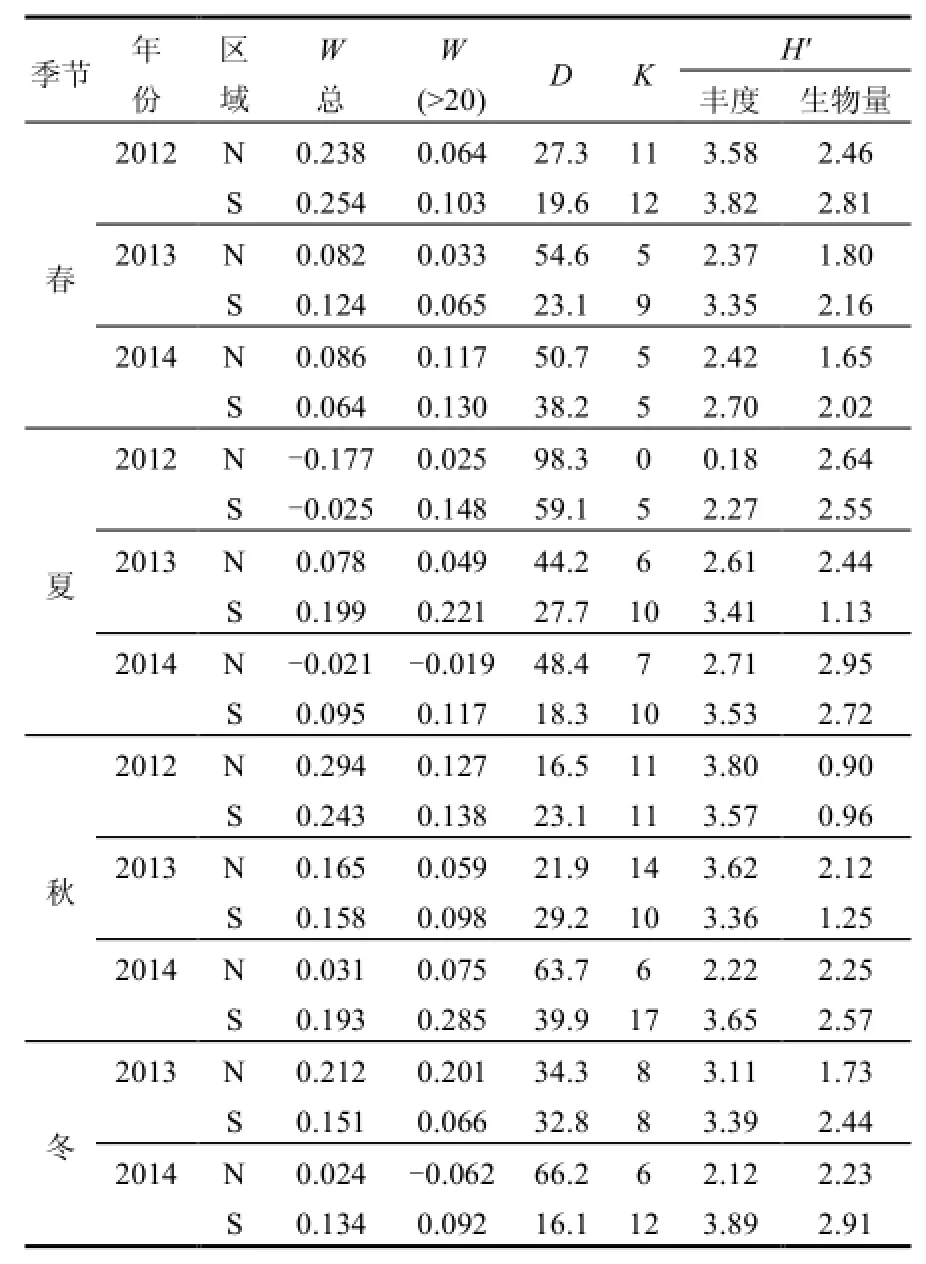
表5 ABC曲线及多样性水平统计参数Table 1 The statistical parameter of ABC curves and diversity level
N、S区2012~2014年4个季节浮游植物群落的ABC曲线变化如图4所示,总体而言,N、S区生物量优势度曲线基本都在丰度优势度曲线之上,仅2012年夏季N、S区丰度优势度曲线居上,N区的2013年春季和2014年夏、秋、冬季2个优势度曲线更替相交.说明N、S区春、秋、冬季浮游植物群落结构整体上比较稳定,仅夏季偏离平衡点.
从W总统计量来看(表5),数值基本为正,仅在夏季(2012年N、S区和2014年N区)出现负值.S区W统计量整体上高于N区,尤其ESD>20μm粒径组(除2013年冬季外)均高于N区,说明S区群落结构整体上比N区稳定.
2.5多样性水平
图4中的丰度优势度曲线即为k-优势度曲线(k-dominance curve),用来检验种类丰度组成中包含的优势种类的多少,可进行多样性水平的分析与评价,曲线位置越低代表群落多样性水平越高[25-26],而第一优势度决定了k-优势度曲线的起始位置.总体来看,N区浮游植物群落多样性水平依次为秋>春>冬>夏,与多样性指数(H′)统计的规律一致,而 S区多样性水平依次为冬>春>秋>夏,与多样性指数(H′)计算的结果基本一致(表5),总体而言,S区多样性水平高于N区.
3 讨论
本研究选取辽东湾南(S区)、北(N区)2个区域进行浮游植物群落及粒级结构分析,结果显示不仅丰度与生物量上存在差异,其粒级结构和群落稳定性也存在明显差异,说明环境因子的不同是其主要的影响因素.
3.1粒级结构的环境胁迫响应
N、S区中心相距仅150km,但盐度、水温、DIN、COD、Cd、Hg、As差异较为显著,为避免浮游植物低盐种类的影响,N区选取的10个站位盐度均在20以上,S区选取的10个站位盐度均在28以上,调查中N区也未检出淡水种类.由于N区水深相对较浅,同期调查水温在春、夏季略高,秋、冬季略低.重金属 Cd、Hg、As含量在两区之间虽有差异,但差值较小,所测阈值也均在第二类海水水质标准范围内,对浮游植物的群落结构影响较小.而营养盐水平是影响浮游植物粒度的主要因素,调查发现,N区富营养化水平较高,S区则为寡营养水平.
研究发现,N区浮游植物生物量在 20μm<ESD<100μm中粒径区域较S区有明显优势,而在ESD>100μm大粒径区域比例却相对较低.由于是网采样品,ESD<20μm粒径组主要是由小粒径的链状或杆状的骨条藻、帕拉藻、角毛藻、菱形藻等组成,导致低营养水平的S区ESD<20μm小粒径组丰度组成全年高于N区,但在以圆筛藻等短圆柱形为主的ESD>100μm大粒径组生物量整体高于N区.本研究于2014年夏、秋季对同期采集的水样品进行了叶绿素 a分级测定(20、3、0.22μm的滤膜逐级过滤),结果表明,S区<20μm叶绿素a组成(夏88.4%、秋76.5%)均高于N区(夏49.2%、秋72.6%).这与Grover[27]研究结果相似,高营养盐环境更能促进小粒径球形细胞和大粒径细长型细胞的繁殖.许多研究也表明,寡营养海域,微微型浮游植物(0.2~3μm)是初级生产力的主要贡献者,营养丰富的水域,初级生产力主要由粒径较大的小型浮游植物(20~200μm)贡献[2].
另外,温度也是影响浮游植物粒径分布的重要因素,在温度较低的南极普里兹湾,各粒级浮游植物对初级生产力的贡献分别为小型浮游植物69%、微型浮游植物 21%、微微型浮游植物10%[28].本研究在冬季水温较低环境下,浮游植物生物量主要由ESD>100 μm大粒径组贡献,也验证了低水温环境下更适于大粒径浮游植物生存的结论.
由于本研究针对的浮游植物粒级研究是网采样品,不可避免漏掉大部分微型及微微型浮游植物,可能无法体现各粒级浮游植物的实际分布特征,但本研究利用同种方法分析的2个区域,可以反映不同环境胁迫下网采浮游植物的粒级结构响应规律.
3.2群落结构的稳定性评估
ABC曲线法具有生态学中r-选择和k-选择策略的理论基础,可比较分析不同时期,不同胁迫情况下浮游植物群落的响应[29-31].
春季,由于光照充足、温度适宜和营养盐的蓄积给浮游植物生长带来了有利条件,使得中肋骨条藻、具槽帕拉藻、角毛藻等以 r-对策者为主的小个体细胞迅速繁殖,生物量曲线暂时位于丰度曲线之上;夏季,海水营养盐大部分被浮游植物消耗,由于降雨量较小,营养盐没有及时从陆源得到补充(N、S区2012年和2014年无机氮检测含量相对其他季节偏低),导致群落结构偏离平衡点,生物量曲线位于丰度曲线之下;秋季,由于营养盐消耗大于补充,以r-对策者为主的小个体细胞逐渐被 k-对策者的圆筛藻属等大个体细胞演替,群落逐渐恢复到平衡点;到了冬季,主要受水温影响,群落主要由k-对策者的圆筛藻属完成反馈调节.
浮游植物群落结构稳定性评估结果显示,营养化程度较低的S区群落结构整体上比N区稳定,与生物多样性评估结果基本一致.因此,可以尝试利用ABC曲线法定量评估浮游植物群落结构稳定性状况,而对于网采样品,统计 ESD>20 μm粒径组的W统计量评估群落结构稳定性相对更加合理和符合实际情况.
另外,研究区浮游植物类群主要以硅藻为主,甲藻为辅.N区硅/甲藻丰度(生物量)比例在夏、秋、冬季平均值均高于S区,而春季比例接近,说明甲藻更适于S区的生存环境.因为与硅藻相比,甲藻更耐受较低的营养盐,但需要较高的温度和较强的光照.
4 结论
4.1辽东湾海域网采浮游植物小粒径优势种主要由中肋骨条藻、具槽帕拉藻、角毛藻组成,中粒径优势种主要由格式圆筛藻、威氏圆筛藻组成,大粒径优势种主要由星脐圆筛藻、虹彩圆筛藻组成,由于种群分布不均,仅分布于南部区域的物种数量相对于只分布在北部区域的物种数量偏多.
4.2低营养水平的南部区域浮游植物粒径整体上大于北部区域,虽小粒径组丰度比例较高,但大粒径组生物量比例却高于北部区域,群落结构整体上比北部区域稳定.寡营养水域以及较低的水温环境更适于k-对策者的大粒径浮游植物生存.
4.3可以尝试利用网采ESD>20μm粒径组的W统计量作为浮游植物群落结构稳定性的定量评估指标之一.
[1] 马艳娥,柯志新,黄良民,等.夏冬两季大亚湾典型海域浮游植物粒级结构特征 [J]. 热带海洋学报, 2013,32(3):40-46.
[2] 杨茹君,王修林,韩秀荣,等.海洋浮游植物粒径组成及其生物粒径效应研究 [J]. 海洋科学, 2003,27(11):5-9.
[3] 周林滨,谭烨辉,黄良民,等.水生生物粒径谱/生物量谱研究进展[J]. 生态学报, 2010,30(12):3319-3333.
[4] 李清雪,陶建华.应用浮游植物群落结构指数评价海域富营养化[J]. 中国环境科学, 1999,19(6):548-551.
[5] 孙 军,刘东艳,钱树本.浮游植物生物量研究I.浮游植物生物量细胞体积转化法 [J]. 海洋学报, 1999,21(2):75-85.
[6] 左 涛,王 俊,金显仕,等.春季长江口邻近外海网采浮游生物的生物量谱 [J]. 生态学报, 2008,28(3):1174-1182.
[7] 左 涛,王 俊,唐启升,等.秋季南黄海网采浮游生物的生物量谱 [J]. 海洋学报, 2008,30(5):71-80.
[8] 吴金浩,徐雪梅,杨 爽,等.2007年春、秋辽东湾北部营养盐及影响因素 [J]. 海洋科学进展, 2012,30(4):477-486.
[9] 周遵春,马志强,薛 克,等.对辽东湾夜光藻赤潮和叉状角藻赤潮成因的初步研究 [J]. 水产科学, 2002,21(2):9-12.
[10] 栾 莎,宫相忠,双秀芝,等.2009年春季辽东湾网采浮游植物群落结构 [J]. 海洋科学, 2012,36(5):57-64.
[11] 康元德,王 俊.渤海浮游植物的数量分布和季节变化 [J]. 海洋水产研究, 1991,12(12):31-54.
[12] 俞建銮,李瑞香.渤 海,黄海浮游植物生态的研究 [J]. 黄渤海海洋, 1993,11(3):52-59.
[13] 王 俊,康元德.渤海浮游植物种群动态的研究 [J]. 海洋水产研究, 1998,19(1):51-59.
[14] 宋 伦,周遵春,王年斌,等.辽东湾浮游植物多样性与海洋环境的关系 [J]. 海洋环境科学, 2007,26(4):365-368.
[15] 耿文华,陈继淼,冯剑丰,等.辽河浮游植物群落及生物多样性基准验证 [J]. 中国环境科学, 2014,34(1):239-245.
[16] 宋 伦,王年斌,宋永刚,等.辽宁近岸浑浊海域网采浮游生物的粒径结构特征 [J]. 应用生态学报, 2013,24(4):900-908.
[17] GB/T 12763-2007 海洋调查规范 [S].
[18] GB 17378-2007 海洋监测规范 [S].
[19] 孙 军,刘东艳.中国海区常见浮游植物种名更改初步意见 [J].海洋与湖沼, 2002,33(3):271-286.
[20] Eppley RW, Reid F M, Strickland J D. The ecology of the plankton off La Jolla, Callifornia, in the period April through September 1967Par Ⅱ. Estimates of phytoplankton crop size, growth rate and primary production [J]. Bulletin of the Scripps Institution of Oceanography, 1970,17:33-42.
[21] Pinkas L, Oliphant M S, Iverson I L K. Food habits of albacore, bluefin tuna, and bonito in California waters [J]. California Department of Fish and Game Fish Bulletin, 1971,152:1-105.
[22] 郭卫东,章小明,杨逸萍,等.中国近岸海域潜在性富营养化程度的评价 [J]. 台湾海峡, 1998,17(1):64-70.
[23] GB3097-1997 海水水质标准 [S].
[24] Warwick R M. A new method for detecting pollution effects on marine macrobenthic communities [J]. Marine Biology, 1986, 92(4):557-562.
[25] Ellis J R, Rogers S I, Freeman S M. Demersal assemblages in the Irish Sea, St George's Channel and Bristol Channel [J]. Estuarine, Coastal and Shelf Science, 2000,51(3):299-315.
[26] Platt H M, Shaw K M, Lambshead P J D. Nematode species abundance patterns and their use in the detection of environment perturbations [J]. Hydrobiologia, 1984,118(1):59-66.
[27] Grover JP. Influence of cell shape and size on algal competitive ability [J]. Journal of Phycology, 1989,25:402-405.
[28] 刘子琳,蔡昱明,陈中元,等.1998/1999年南极夏季普里兹湾及北部海区叶绿素 a和初级生产力的分布特征 [J]. 极地研究, 2002,14(1):12-21.
[29] Yemane D, Field J G, Leslie R W. Exploring the effects of fishing on fish assemblages using Abundance Biomass Comparison (ABC) curves [J]. ICES Journal of Marine Science, 2005,62(3):374-379.
[30] 曲方圆,于子山,隋吉星,等.丰度生物量比较法应用局限性 [J].海洋科学, 2009,33(6):118-121.
[31] 宋 伦,王年斌,杨国军,等.鸭绿江口及邻近海域生物群落的胁迫响应 [J]. 生态学报, 2013,33(9):2790-2802.
The stress response of net phytoplankton biomass size structure in Liaodong Bay.
SONG Lun1,2*, SONG Guang-jun1,2, WANG Nian-bin1,2, ZHAO Hai-bo1,2, TIAN Jin1,2, YANG Shuang1,2, DU Jing1,2(1.Liaoning Ocean and Fisheries Science Research Institute, Dalian 116023, China;2.Liaoning Ocean Environment Monitoring Station, Dalian 116023, China).
China Environmental Science, 2015,35(9):2764~2771
Based on net collection of phytoplankton samples in Liaodong Bay in all four seasons during 2012~2014, the distribution pattern of size structure of phytoplankton and its response to environmental stress were analyzed in the northern and southern areas of the bay. The stability of the phytoplankton community was estimated and was found to be dominated by small species in spring, summer and autumn, and by medium-sized species in winter. Phytoplankton biomass was controlled by medium-sized species in spring, summer and autumn, and by large species in winter. In general, phytoplankton sizes in the southern area were greater than those in the northern area. Large phytoplankton species were much more adaptive to oligotrophic water and a colder environment. The phytoplankton community in spring, autumn and winter was relatively stable; however, it deviated from equilibrium in summer. Phytoplankton community stability in the southern area was slightly better than in the northern area. The W statistical value of the group of large species can be used as an evaluation index of phytoplankton community stability.
phytoplankton;size structure;ABC curves;bionomic strategy;ecological stability;Liaodong Bay
X171.1;Q89;P735
A
1000-6923(2015)09-2764-08
2015-01-11
辽宁省自然科学基金资助项目(2014020182);辽宁省海洋与渔业科研项目(201415);国家自然科学基金资助项目(31400406)
*责任作者, 副研究员, songlun@lnshky.com
宋 伦(1980-),男,辽宁大连人,副研究员,硕士,主要从事海洋生物生态学研究.发表论文30余篇,专著2部.
