疏叶骆驼刺和多枝柽柳不同时期光合特性日变化及其与环境因子的关系
2015-07-04郭自春曾凡江李尝君
郭自春,曾凡江,刘 波,李尝君,3,张 波
(1 中国科学院新疆生态与地理研究所/新疆策勒荒漠草地生态系统国家野外科学观测试验站/中国科学院干旱区生物地理与生物资源重点实验室/中国科学院荒漠与绿洲生态国家重点实验室,乌鲁木齐830011;2中国科学院大学,北京100049;3新疆大学资源与环境科学学院,乌鲁木齐830046)
光合特性是植物适应环境最重要的特征和物质积累代谢的基本单元,也是植物生长发育的基础和生产力高低的决定性因素[1-3]。它不仅受到植物本身遗传特性的影响[4],同时还受到环境条件的制约[3]。在自然条件下,环境因子对光合作用的影响不是单一的,而是多因素相互联系、相互制约的结果[5-6]。比如光照强度、空气温度、CO2浓度和空气相对湿度等环境因子不仅直接影响着光合作用,而且还通过植物的生理因子来间接地影响植物的光合作用。此外,植物生长的不同时期内各环境因子之间存在着相互作用和较大差异[5,7],而这些不断变化的环境因子可能导致植物在不同生长时期表现出不同的光合特征,继而导致植物在不同时期内表现出不同的生长特性。因此,有必要对不同时期植物光合特性及其与环境因子之间关系进行研究,以期更好地理解在特定生境下各个环境因子对植物光合作用的综合影响,确定对光合作用影响较为显著的环境因子,找出植物适应该环境条件的主要原因,进而为深入研究植物适应环境机制提供理论基础。
骆驼刺是生长于荒漠半荒漠地区的多年生豆科木质化的草本植物,含有较高的蛋白质,既是防风固沙的自然植被,又是牲畜不可缺少、也无法替代的饲草之一[8],具有重要的经济价值。柽柳是分布最为广泛的柽柳属植物之一,是维持干旱区荒漠生态系统的关键树种,目前已成为新疆南部管花肉苁蓉栽培选用的主要寄主之一[9],具有很高的经济价值。近年来,随着当地居民对骆驼刺过度采伐和放牧[10],以及对肉苁蓉的过度采挖,过渡带生态环境面临着越来越巨大的压力,使得该地区本来就已经十分严峻的生境进一步恶化。刈割和放牧不仅会影响牧草质量和产量[11],还对牧草的光合特性有一定的影响[12];管花肉苁蓉的寄生也会影响柽柳的光合特性[13]。每种植物在特定的环境中都有复杂的适应机制来生存和发展[14],而骆驼刺和柽柳作为策勒绿洲过渡带上所形成的优势植物群落,必然在其独特的环境条件中产生了一定的适应性,这对于维护脆弱的过渡带地区生态环境有着重要的意义[15]。
目前,关于骆驼刺和柽柳在不同水分胁迫[16-17]和盐胁迫[18-19]等控制条件下的光合特性已有大量的报道,但对自然条件下尤其是绿洲与荒漠过渡带生境下,骆驼刺和柽柳不同时期内光合特性的日变化及其与同期内环境因子的日变化的综合分析还未见报道。为此,本研究通过对骆驼刺和柽柳不同生长时期光合参数日变化的测定及与其环境因子之间相互关系的分析,寻找影响植物光合作用的主导环境因子,为以后更好有效地恢复和重建过渡带植被提供理论依据。
1 材料和方法
1.1 实验区概况
该实验依托中国科学院新疆生态与地理研究所策勒国家野外科学观测研究站(简称策勒站)进行。策勒站位于塔克拉玛干沙漠南缘中段,该地区属典型的大陆性干旱气候,年平均温度11.9 ℃;多年平均降水量仅35.1mm,且主要集中在5月和7月;平均蒸发量2 595.3mm,干燥度20.8。夏季炎热,日照时间长,昼夜温差大,极端最高气温41.9℃,极端最低气温-23.9 ℃。区域内风沙灾害频繁,多年平均沙尘日数25.2d,最多高达89d。土壤以风沙土和棕漠土为主,物质组成以极细砂和粉砂为主,质地轻,粒径细,易形成风沙流。过渡带生境内自然植被种类组成单一,主要以骆驼刺、柽柳为建群种。
该区域内生态环境脆弱,降水稀少,蒸发强烈,地下水矿化度高和土壤盐渍化问题[20],以及人口的增加和不合理利用自然植物的破坏[21],使得该地区受到非常严重的荒漠化危害。骆驼刺为多年生豆科木质化草本植物,它可通过深根系与地下水连接,并利用地下水维持生存,并以克隆分株进行无性繁殖为主,当地居民通常在秋冬季刈割骆驼刺作为牲畜越冬饲料贮藏。柽柳具有耐干旱、耐盐碱、耐贫瘠、耐风蚀和沙埋等特征,目前作为当地管花肉苁蓉栽培选用的主要寄主之一。长期以来,人们对骆驼刺过度放牧和刈割以及对肉苁蓉的过度采伐,导致过渡带生态环境恶化,极易遭受流沙侵袭。
1.2 实验方法
观测实验在中国科学院新疆策勒荒漠草地生态系统国家野外研究站天然荒漠综合实验场进行。在长期进行刈割和放牧的样地内,选健康、长势基本一致的6株标准样株。由于该地区风沙降尘较多,所以在测定前一天,先将待测向阳面叶片表面的尘土用喷水壶清洗、擦干。分别于2012年7月、8月、9月份用Li-6400 便携式光合系统测定仪(LICOR,USA)对骆驼刺和柽柳叶片的净光合速率(Pn)蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等生理因子,以及光合有效辐射(PAR)、空气温度(Ta)、大气CO2浓度(Ca)、相对湿度(RH)等环境因子进行测定。并在以上参数的基础上计算气孔限制值(Ls)和水分利用效率(WUE),公式分别为:
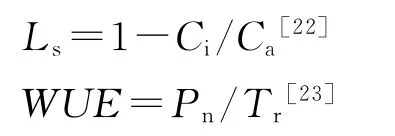
由于柽柳叶片退化,因此选择具有光合作用的同化枝进行测定;骆驼刺叶片极小,选择连同叶轴一起测定。光合参数测定完之后,采集测定的叶片带回实验室,用扫描仪进行扫描,再经DT Scan软件(Techne,Cambridge,UK)计算出实际的光合叶面积,并利用Li-6400提供的重算功能重新计算,得出实际的观测数据。参数测定时间为8:00~20:00,每隔3h测定1次,测定时待数据稳定后读取5个测量数据,取平均值作为该时刻的实测值。
1.3 数据处理
所有的数据均用Excel 2003 进行整理。首先利用SPSS 17.0进行不同时期光合特征参数的单因素方差分析(One-way ANOVA),然后进行环境因子与净光合速率的相关性分析,最后用SPSS 17.0进行多元回归分析和通径分析。当许多自变量共同影响一个因变量时,其中一个自变量可通过其他自变量对因变量起作用,间接通径系数可以表示为:Pij=rij·Pjy(其中,rij是自变量i和自变量j的简单相关系数,Pjy是自变量j对因变量Y 的直接作用)[6,24]。决策系数R2=2Pi·riy-Pi2(Pi为自变量i对因变量的直接通径系数,riy为自变量i与因变量的Pearson 相关系数)是通径分析中的决策指标[25]。所有的作图均在Origin 8.0完成。
2 结果与分析
2.1 实验区环境因子日变化特征
实验区自然条件下主要环境因子日变化如图1所示。其中,各月份PAR都是从日出后逐渐增大,在14:00达到最大值后则逐渐减小,变化幅度很大;各月份RH、Ca均随着PAR和Ta的升高而逐渐下降,并在早晨达到最大值,午间降至最低,而后由于PAR和Ta的降低,RH先降低后升高,Ca的变化幅度不大;3个月份的平均气温分别为32.4 ℃、35.4 ℃和31.3℃,均高出荒漠植物最适宜温度[26]。
2.2 骆驼刺和柽柳气体交换参数的日变化特征
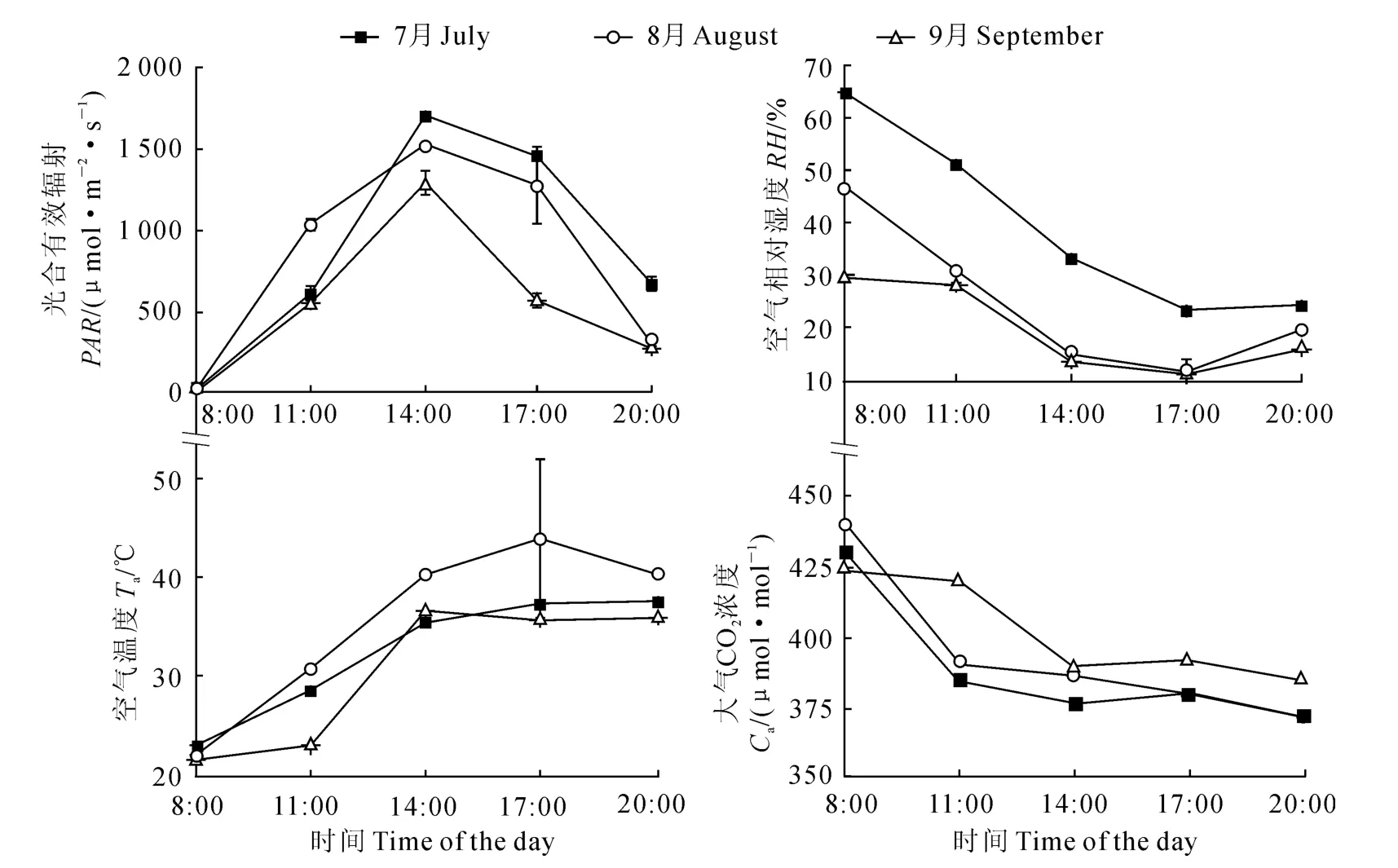
图1 实验区不同月份主要环境因子日变化(平均值±标准误)Fig.1 Diurnal changes of environmental factors at different months of experimental area(mean±SE)
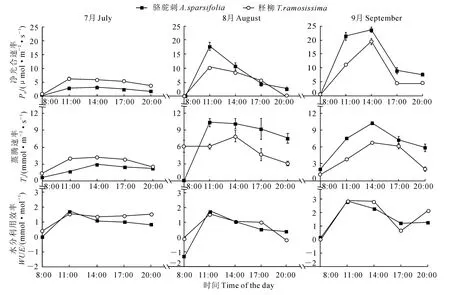
图2 不同月份骆驼刺和柽柳光合特性的日变化(平均值±标准误)Fig.2 Diurnal changes of photosynthetic characteristics of A.sparsifoliaand T.ramosissimaat different months(mean±SE)
2.2.1 净光合速率、蒸腾速率和水分利用效率 图2显示,骆驼刺Pn日变化在3个月份均为单峰曲线,而柽柳Pn的日变化在7月和8月份为单峰曲线,在9月份却为双峰曲线;骆驼刺和柽柳7月和8月份Pn均在11:00左右达到峰值,但峰值有所差异,7月份分别为2.84和6.10μmol·m-2·s-1,8月份分别为1 7.7 4和10.21μmol·m-2·s-1;而 它 们 在9月份于14:00 左右达到峰值,在17:00左右达到谷值;就3个月整体来看,骆驼刺Pn的平均值(7.08 μmol·m-2·s-1)高于柽柳(5.54μmol·m-2·s-1)。同时,3个月份骆驼刺和柽柳的Tr日变化均表现为单峰曲线,这是因为PAR和Ta的增大,有利于气孔开张,Tr也随之增大;而午后Tr下降可能是由于RH的降低从而引起气孔关闭,这也是骆驼刺和柽柳减少水分散失的一种适应方式;就3个月整体来看,骆驼刺Tr平均值(5.46 mmol·m-2·s-1)高于柽柳(4.40mmol·m-2·s-1)。
另外,骆驼刺和柽柳的WUE日变化进程与各自Pn日变化规律基本一致,除9月份柽柳最大值出现在14:00时外,其它时期骆驼刺和柽柳WUE的最大值都出现在11:00时左右;就3个月份整体来看,柽柳WUE日均值(1.21mmol·mol-1)明显高于骆驼刺(0.97mmol·mol-1)。2种植物相比较而言,骆驼刺的Pn和Tr高于柽柳,而WUE低于柽柳,这一结果与骆驼刺属于低光合低蒸腾、柽柳属于高光合高蒸腾的结论不一致[25]。
2.2.2 气孔导度、胞间CO2浓度和气孔限制值 骆驼刺和柽柳的气孔导度(Gs)除8月份以外的日变化趋势均表现为单峰曲线;8月份的Gs日变化相差较大,从早晨气孔逐渐开放,于11:00 左右达到峰值,之后开始下降,在17:00时出现低谷,这可能与午后光照加强引起叶片水分蒸腾强烈有关。骆驼刺在3个月内Gs变化非常明显,表现为7月最低,8月次之,9月最高,而柽柳Gs在3个月间的变化基本不明显,这也是骆驼刺对于极限逆境的一种自我调整[26]。同时,骆驼刺和柽柳叶片胞间CO2浓度(Ci)的日变化趋势呈“倒钟型”(图3),与Pn的日变化趋势相反,这是因为当Pn较大时,固定的CO2较多,进而引起了Ci的降低。根据Farquhar等[27]的理论用Ls和Ci的变化来判断Pn下降的原因,7月份14:00之后骆驼刺和柽柳的Pn和Gs下降,而Ci降低、Ls增大,所以Pn下降主要因素为气孔限制;8月和9月14:00之后骆驼刺和柽柳的Pn和Gs下降,而Ci增大、Ls减小,则Pn下降归因于叶肉细胞同化能力的降低,即非气孔限制因素所致。
2.3 骆驼刺和柽柳净光合速率与环境因子之间的关系
2.3.1 净光合速率与环境因子的相关性 不同月份骆驼刺和柽柳光合特性与环境因子相关性分析结果表明(表1),叶片的Pn与其它环境因子在不同月份有一定的相关性,但相关关系各异,相关程度也各不相同。从简单相关系数分析来看,7月份骆驼刺和柽柳的Pn与RH、Ca呈显著负相关(P<0.01),与PAR、Ta呈显著正相关(P<0.01);8月份Pn与RH、Ca仍呈显著负相关(P<0.01),与PAR呈显著正相关(P<0.01),而与柽柳的Ta呈显著正相关(P<0.01),与骆驼刺的Ta无显著相关;9月份Pn与PAR呈显著正相关(P<0.01),与RH呈显著负相关(P<0.05),而与柽柳的Ta呈显著正相关(P<0.01),与骆驼刺的Ta无显著相关。
由于各环境因子之间具有相互作用,Pn与各环境因子之间的相关系数并未完全反映出各因子之间的真实情况,应该看偏相关的结果找到真实联系最为密切的因子。从偏相关系数来看,骆驼刺和柽柳Pn与PAR呈显著正相关关系,而与RH达到显著负相关关系。
2.3.2 环境因子对净光合速率的逐步线性回归分析 运用多元回归分析方法,挑选对Pn影响较大的因子PAR、RH、Ca和Ta进行逐步多元回归分析,得到了不同月份骆驼刺和柽柳的Pn与环境因子的线性回归方程(表2)。从表中可以看出,不同月份骆驼刺和柽柳的回归模型均达到显著水平(P<0.01),回归模型的可信度分别为59.7%(R2=0.356)和65.1%(R2=0.424),这说明骆驼刺和柽柳Pn日变化的变异平方和分别有35.6%和42.4%是由环境因子的日变化造成的。
2.3.3 环境因子对净光合速率的通径分析 为进一步了解各环境因子作用大小,又对Pn与各个因子进行通径分析。结果发现,影响骆驼刺和柽柳各月Pn的环境因子及其影响程度均存在一定的差异(表3)。其中,骆驼刺7~9月份Pn主要受PAR和Ca的影响,且具有较大的间接通径系数;柽柳7~9月份Pn受PAR、Ca和Ta的影响较大;就这3个月整体来看,各环境因子对Pn都具有显著的影响,其大小顺序分别为:Ta>RH>PAR>Ca(骆驼刺)和PAR>Ca>Ta>RH(柽柳)。
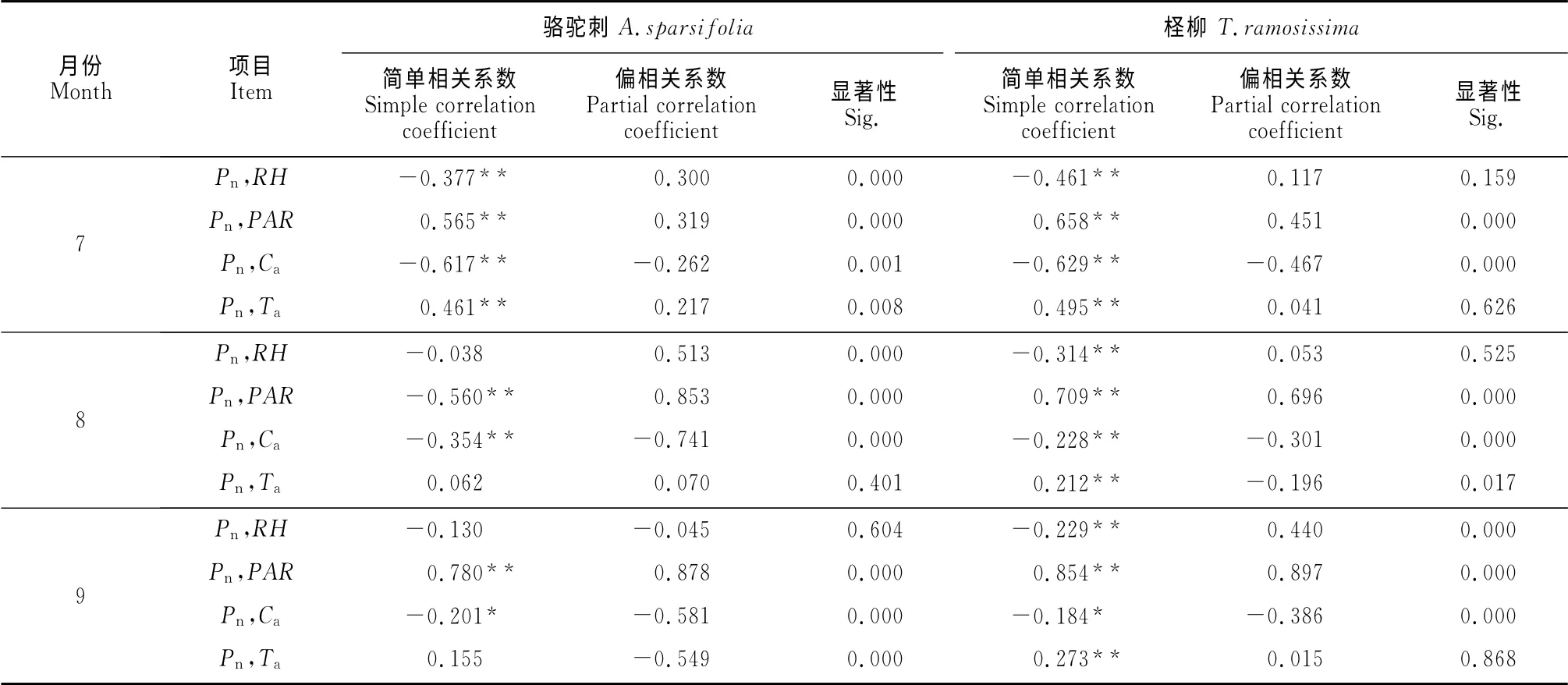
表1 不同月份骆驼刺和柽柳净光合速率与环境因子相关性分析Table1 Correlation analysis of A.sparsifoliaand T.ramosissimaat different months between environmental factors and Pn
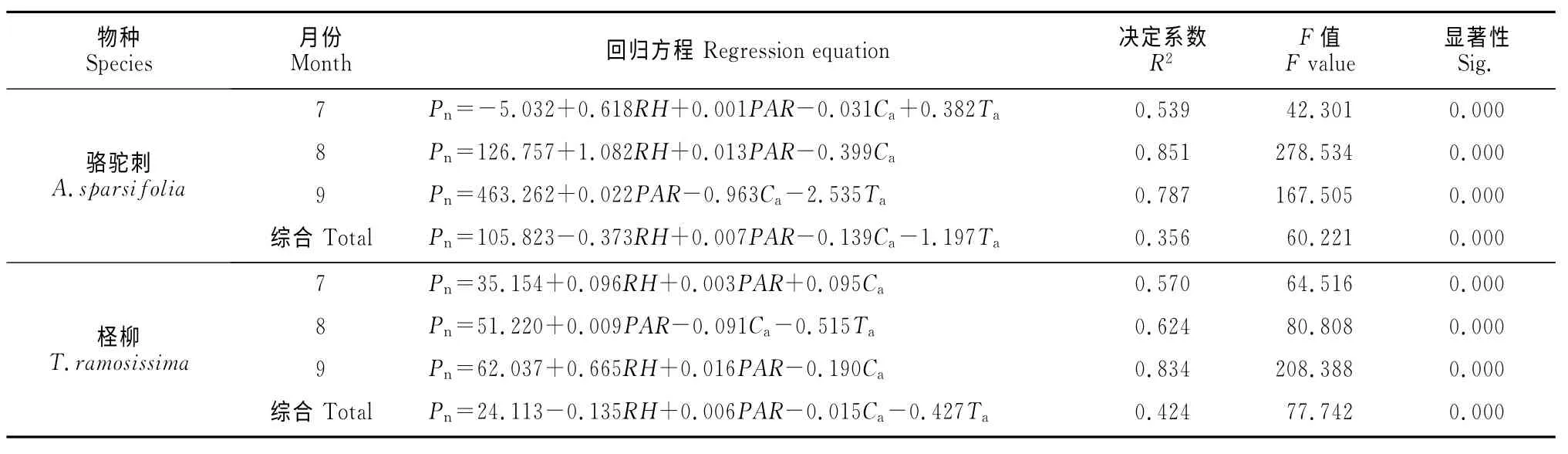
表2 不同月份骆驼刺和柽柳净光合速率多元线性回归方程Table2 Multiple linear regression equations of A.sparsifoliaand T.ramosissima for net photosynthetic rate at different months
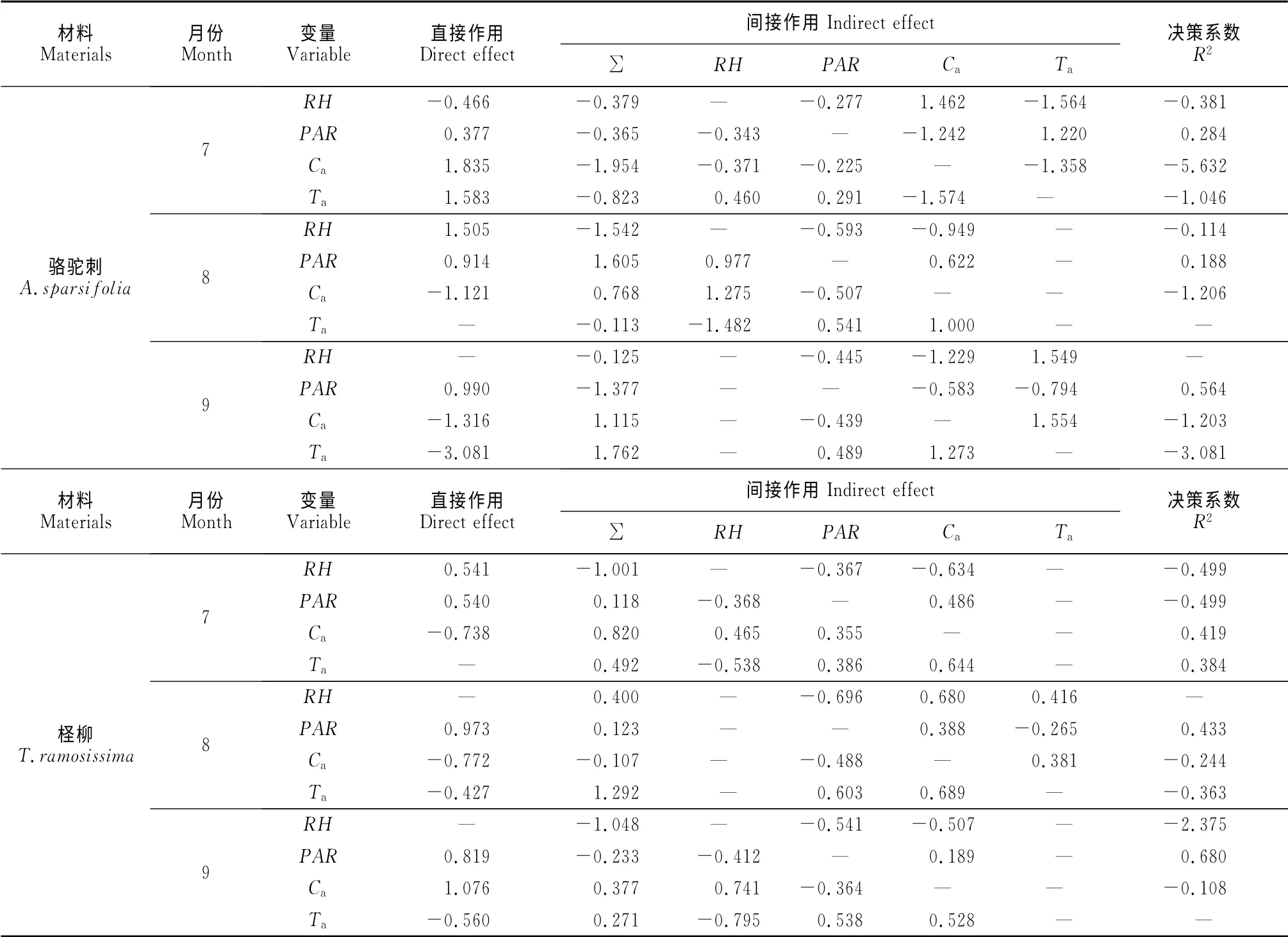
表3 不同月份环境因子对骆驼刺和柽柳净光合速率的通径分析Table3 Path analysis of A.sparsifoliaand T.ramosissimafor net photosynthetic rate at different months
通过各环境因子对Pn的综合作用(决策分析)进行排序后,最终确定主要决策变量和限制变量。骆驼刺在这3个月份内PAR均是决策变量,RH均是限制变量(除7月份外);而柽柳在8月和9月份内PAR均是决策变量,RH、Ca是限制变量。
3 讨 论
3.1 骆驼刺和柽柳不同时期光合参数日变化特征
光合特性日变化是植物生产过程物质积累与代谢的基本单元[2],也是分析环境因子影响植物生长和代谢的重要手段。一般而言,荒漠植物光合作用的日变化趋势都呈“单峰”和“双峰”变化。已有研究表明,骆驼刺和柽柳的净光合速率和蒸腾速率的日变化曲线特征都属于双峰型,其中骆驼刺属于低光合低蒸腾型,而柽柳属于高光合高蒸腾型[16,27]。不同植物由于本身的遗传特性和环境因子的不同,对环境的适应能力也存在着较大的差别[28]。但在相同生境下,骆驼刺和柽柳的光合作用的日变化类型和大小表现出明显的不同,说明这2种植物的光合特性在生境条件发生改变的情况下进行了自我调节,以适应环境的变化。本研究中骆驼刺Pn的日变化在3个月份都表现为单峰曲线;而柽柳Pn的日变化在7月和8月份为单峰曲线,9月份为双峰曲线;相比于柽柳,整个观测时期内骆驼刺Pn和Tr的平均值均较大,属于高光合高蒸腾型,这说明在同一生境中骆驼刺具有较高的CO2固定能力,这可能是由于人为干扰导致Pn,Tr和Gs发生明显的变化[12,29-30],从而影响荒漠生态系统的第一性生产力[31]。
有学者认为,光合作用可以作为判断植物生长和抗逆的指标[32]。然而也有学者认为,光合和蒸腾是一个复杂耦合的过程,并不能定量地来评价植物适应环境的能力[33],而WUE可以有效地评价植物适应环境的能力。在干旱半干旱极端环境下,WUE是决定植物生存和发展的一个关键因素[34]。在相同的环境条件下,WUE越大,表明固定单位质量CO2所需要的水分越少,植物抗旱能力越强[35]。本研究发现,虽然骆驼刺的Pn和Tr比柽柳的大,但是就整体WUE的日均值来看,柽柳的WUE明显高于骆驼刺,所以由此推断出柽柳的抗旱能力强于骆驼刺。因此,本研究也认为判断植物适应环境的能力需要综合考虑多方面因素。
气孔是植物叶片与大气之间进行水气交换的通道,主要是通过气体的调节来影响光合作用的。一般认为Pn的下降主要是由于气孔限制和非气孔限制两种原因造成的,当两种因素同时存在时,以Ci的变化方向为主,而不是以Ci的变化幅度作为标准[36]。本研究中,7月份骆驼刺和柽柳Pn的下降主要是由于气孔限制的,而8月和9月主要是由于非气孔因素限制。这与邓雄等[26]的结论不一致,除了与测定的时间不同以外,与植物目前所生存的环境有很大的关系,因为植物通过调节气孔大小在光合所需要的CO2和可利用的水资源之间寻求折中点[37],从而使得植物气孔导调节更加倾向于植物适应环境的方向发展。
3.2 骆驼刺和柽柳不同时期环境因子对净光合速率的影响
植物叶片Pn的大小是衡量植物光合作用能力强弱的关键指标[38]。影响植物叶片Pn的环境因子主要有PAR、RH、Ca和Ta,这些因子对Pn的影响既有直接作用,也有间接作用。而且不同时期对植物叶片的Pn起主导作用的环境因子有所差异,同一环境因子在不同时期对Pn的影响程度各异[39]。在本研究中,在7月份,骆驼刺和柽柳处于生长初期,Ta和PAR相对较小,且直接通径系数较小,因此该时期叶片Pn相对较低;而8月份和9月份2种植物处于生长季旺盛期,Ta和PAR相对较高,且直接通径系数较大,Pn显著大于7月份,这也同时说明强光环境并未对骆驼刺和柽柳造成胁迫。一定温度范围内,Ta和Pn呈正比例关系,这是由于25~30 ℃是Rubisco最为适宜温度范围[40]。在本研究中,2种植物的平均温度均高出荒漠植物的最适宜温度,甚至日最高温度达到40 ℃以上(8月份),这就说明植物适应生境变化的过程中,温度生态位的范围变得更大,这一结果与邓雄等[41]的研究结果一致。PAR是植物光合作用能量的最终来源,也是影响光合作用的主导因子[6],在一定范围和时间内,PAR和Pn之间有较好的正比例关系[30]。而在本研究中由于柽柳9月的Pn有光合“午休”现象,所以在一定程度上消弱了这种相关关系。
在自然条件下,环境因子对植物Pn的影响不是单一不变的,而是综合作用的结果。关于Pn与各环境因子的关系目前存在不同的观点。吴统贵等[42]通过研究旱柳(Salixmatsudana)整个生长季的Pn与环境因子之间的关系发现,各因子对Pn变化的影响显著,且Ta是主要的影响因子;邓雄等[41]的研究认为,在外界环境因子中PAR是影响骆驼刺和柽柳Pn变化最重要的主导因子,其次是Ta;许皓等[43]的研究表明,PAR是光合作用物候学特征的一个最主要的影响因素。本研究通过相关分析、多元回归分析以及通径分析发现,PAR和RH是影响骆驼刺和柽柳Pn最重要的环境因子,其次是Ca,而Ta在不同的时期影响程度不同,这与其他研究的成果不尽一致。这主要有以下3个方面的原因:首先,与物种有关。本实验所选用的过渡带地区生境中的2种优势种,而前人的结论是在同一属植物得到的结论,所以不同的品种生物学特性具有很大的差异;其次,与柽柳所处的生境有关系。本实验是在过渡带生境中进行的,而许皓等[43]是在盐生荒漠中进行的,所以不同的生境下环境因子对植物的光合特性的影响也是不同的;最后,与人为干扰有关。本研究中,过渡带生境中的骆驼刺受到了不同程度的刈割和放牧,对生境变化有了一定的生态适应特征,而邓雄等[41]所研究的骆驼刺生长在绿洲边缘,人为干扰小,但是骆驼刺在不同干扰程度下的光合特性的适应机制还需要进一步的研究分析。
综合上述结果表明,骆驼刺属于高光合高蒸腾低水分利用效率型,而柽柳属于低光合低蒸腾高水分利用效率型;7月份骆驼刺和柽柳Pn的下降主要是由于气孔限制引起,而8月和9月主要是由于非气孔因素限制所致;PAR和RH是影响骆驼刺和柽柳最重要的环境因子,其次是Ca,而Ta在不同的时期影响程度不同,且PAR均是决策变量,RH均是限制变量。
致谢:感谢罗维成、宋聪在实验和取样上给予的帮助。感谢唐钢梁对论文撰写提供的宝贵意见。
[1] GAGO J,COOPMAN R E,CABRERA H M,etal.Photosynthesis limitations in three fern species[J].PhysiologiaPlantarum,2013,149(4):599-611.
[2] WAN S M(万素梅),JIA ZH K(贾志宽),YANG B P(杨宝平).Relationship between diurnal changes of alfalfa net photosynthetic rate and environmental factors[J].ActaAgrestiaSinica(草地学报),2009,17(1):27-31(in Chinese).
[3] COLLATZ G J.Influence of certain environmental factors on photosynthesis and photorespiration inSimmondsiachinensis[J].Planta,1977,134(2):127-132.
[4] LEWIS L D,LUCASH M,OLSZYK D,etal.Seasonal patterns of photosynthesis in douglas fir seedlings during the third and fourth year of exposure to elevated CO2and temperature[J].Plant,CellandEnvironment,2001,24(5):539-548.
[5] ZHANG J L(张津林),ZHANG ZH Q(张志强),ZHA T G(查同刚),etal.Physiological regulations of photosynthesis in a popular plantation on a sandy soil[J].ActaEcologicaSinica(生态学报),2006,26(5):1 523-1 532(in Chinese).
[6] JIN T T(靳甜甜),FU B J(傅伯杰),LIU G H(刘国华),etal.Diurnal changes of photosynthetic characteristics ofHippophaerhamnoidesand the relevant environment factors at different slope locations[J].ActaEcologicaSinica(生态学报),2011,31(7):1 783-1 793(in Chinese).
[7] COXSON D S,COYLE M.Niche partitioning and photosynthetic response of alectorioid lichens from sublpine spruce-fir forest in northcentral British Columbia:the role of canopy microclimate gradients[J].TheLichenologist,2003,35(2):157-175.
[8] ZENG F J(曾凡江),ZHANG X M(张希明),LI X M(李小明).Study on the characteristics ofAlhagiand its impact on resources protection and development[J].AridLandGeography(干旱区地理),2002,25(3):286-288(in Chinese).
[9] ZHANG Y(张 媛),TU P F(屠鹏飞).Chemical constituents ofTamarixramosissima[J].ChineseTraditionalandHerbalDrugs(中草药),2006,37(12):1 764-1 768(in Chinese).
[10] GRIES D,FOETZKI A,AMDT S K,etal.Production of perennial vegetation in an oasis-desert transition zone in NW China-allometric estimation,and assessment of flooding and use effects[J].PlantEcology,2005,181(1):23-43.
[11] ZHANG L Y(张立运),ANWAR M(安尼瓦尔·买买提),XIA Y(夏 阳).Effects of the morphological characteristics,ecological structure,natural regeneration ofAlhagisparsifoliato kharif irrigation[J].AridZoneResearch(干旱区研究),1995,12(4):34-40(in Chinese).
[12] JIANG H(姜 华),BI Y F(毕玉芬),HE CH G(何承刚).Effects of different timing cutting on production,crude protein content and photosynthetic rate ofRyegrass[J].JournalofYunnanAgriculturalUniversity(云南农业大学学报),2003,18(2):149-152(in Chinese).
[13] GUO D F(郭东锋),GUO Y H(郭玉海),HUANG Y(黄 勇).Influence on the photosynthesis ofTamarixchinensisparasited byCistanchetubulosa[J].JournalofAnhuiAgricultureScience(安徽农业科学),2009,37(23):11 293-11 295(in Chinese).
[14] GUTTEMAN Y.Seed germination in desert plants[M].Springer-Verlag GmbH &Co.KG,1993.
[15] THOMAS F M,ARADT S K,BRUELHEIDE H,etal.Ecological basis for a sustainable management of the indigenous vegetation in a Central-Asian Desert:presentation and first results[J].JournalofAppliedBotany,2000,74(5-6):212-219(in Chinese).
[16] ZHU J T(朱军涛),LI X Y(李向义),ZHANG X M(张希明),etal.Effects of irrigation on photosynthetic physiology characteristics and osmolytes ofAlhagisparsifolia[J].JournalofDesertResearch(中国沙漠),2009,29(4):697-702(in Chinese).
[17] WANG SH SH(王珊珊),CHEN X(陈 曦),WANG Q(王 权),etal.Ecological adaptability of photosynthesis and water use forTamarixramosissimain the southern periphery of Gurbantunggut Desert,Xinjiang[J].ActaEcologicalSinica(生态 学 报),2011,31(11):3 082-3 089(in Chinese).
[18] WANG W H(王伟华),ZHANG X M(张希明),YAN H L(闫海龙),etal.Effects of salt stress on photosynthesis and osmoregulation substance ofTamarixramosissimaLedeb.[J].AridZoneResearch(干旱区研究),2009,26(4):561-568(in Chinese).
[19] UMETHAN K(吾木提汗·卡克木),HALIL K(库尔班·海利力),CHEN Q J(陈其军),etal.Study on daily change and midday depression of photosynthesis ofAlhagipseudoalhagiandVignaradiataunder salt stress[J].AridZoneResearch(干旱区研究),2012,29(6):1 039-1 045(in Chinese).
[20] 张希明,MICHAEL R.塔克拉玛干沙漠边缘植被可持续管理的生态学基础[M].北京:科学出版社,2006.
[21] XIA X CH(夏训诚),LIU X M(刘新民),LI CH SH(李崇舜),etal.A study on the mechanism of desertification[J].AridZoneResearch(干旱区研究),1993,10(1):1-14(in Chinese).
[22] BERRY J A,DOWNTON W J S.Environmental regulation of photosynthesis[J].Photosynthesis,1982,2:263-343.
[23] FISCHER R A,TURNER N C.Plant productivity in the arid and semiarid zones[J].AnnualReviewofPlantPhysiology,1978,29(1):277-317.
[24] DU J(杜 鹃).Path analysis implement in Excel and SPSS[J].JournalofShaanxiMeteorology(陕西气象),2012,1:15-18(in Chinese).
[25] YUAN ZH F(袁志发),ZHOU J Y(周静芋),GUO M C(郭满才),etal.Decision coefficient the decision index of path analysis[J].Jour-nalofNorthwestA&FUniversity(西北农林科技大学学报),2001,29(5):131-133(in Chinese).
[26] DENG X(邓 雄),LI X M(李小明),ZHANG X M(张希明),etal.A study of the gas exchange characteristics of four desert plants[J].ActaPhytoecologicaSinica(植物生态学报),2002,26(5):605-612(in Chinese).
[27] HUANG Z CH(黄子琛),PU J CH(蒲锦春),WANG J H(王继和),etal.Net photosynthesis and CO2compensation points of desert plants in Minqin and Shapotou region[J].JournalofDesertResearch(中国沙漠),1984,4(1):23-30(in Chinese).
[28] ZHANG SH Y(张淑勇),ZHOU Z F(周泽福),ZHANG G C(张光灿),etal.Change of gas exchange parameters in leaves of natural secondary shrubsPrunusdavidianaandPrunussibiricaL.in semi-arid Loess Hilly region[J].ActaEcologicaSinica(生态学报),2009,29(1):499-507(in Chinese).
[29] ZHOU X H(周晓红),WANG G X(王国祥),YANG F(杨 飞),etal.Effects of cutting on photosynthesis and purification efficiencies on nitrogen and phosphorus of theLoliummultiflorum[J].EnvironmentalScience(环境科学),2008,29(12):3 393-3 399(in Chinese).
[30] LIU ZH(刘 震),LIU J X(刘金祥),ZHANG SH W(张世伟).Effect of defoliation on photosynthetic characteristics ofStylosanthes guianenesiscv.Reyan No.13[J].GrassLandandTurf(草原与草坪),2009,(1):55-57(in Chinese).
[31] DU ZH CH(杜占池),YANG Z G(杨宗贵).The effect of cutting on the photosynthetic characteristics ofAneurolepidiumchinense[J].ActaPhytoecologicaetGebotanicaSinica(植物生态学与地植物学学报),1989,13(4):317-324(in Chinese).
[32] UEDA Y,NISHIHARA S,TOMITA H,etal.Photosynthetic response of Japanese rose speciesRosabracteataandRosarugosato temperature and light[J].ScientiaHorticulturae,2000,84(3):365-371.
[33] SUN J(孙 建),LIU M(刘 苗),LI SH G(李胜功),etal.Survival strategy ofStipakryloviiandAgropyroncristatumin typical steppe of Inner Mongolia[J].ActaEcologicaSinica(生态学报),2011,31(8):2 148-2 158(in Chinese).
[34] GUO Z CH(郭自春),ZENG F J(曾凡江),LIU B(刘 波),etal.Effects of irrigations on photosynthetic characteristics and water use efficiency ofCalligonummongolicumandHedysarumscoparium[J].JournalofDesertResearch(中国沙漠),2014,34(2):448-455(in Chinese).
[35] ZHAO Y M(赵育民),NIU SH K(牛树奎),WANG J B(王军邦),etal.Light use efficiency of vegetation:A review[J].ChineseJournal ofEcology(生态学杂志),2007,26(9):1 471-1 477(in Chinese).
[36] FARQUHAR G D,SHARKEY T D.Stomatal conductance and photosynthesis[J].AnnualReviewofPlantPhysiology,1982,33(1):317-345.
[37] JARVIS A J,MANSFIELD T A,DAVIES W J.Stomatal behavior,photosynthesis and transpiration under rising CO2[J].Plant,Celland Environment,1999,22(6):639-648.
[38] DING Y F(丁友芳),ZHANG X X(张晓霞),SHI L L(史玲玲),etal.Diurnal change of net photosynthetic rates inPuerarialobataand its relation with environmental factors[J].JournalofBeijingForestryUniversity(北京林业大学学报),2010,5:132-137(in Chinese).
[39] WANG J L(王建丽),ZHANG Y L(张永亮),ZHU ZH L(朱占林),etal.Analysis on ecological characteristics ofMedicagovariaCv.Gannong No.1[J].ActaAgrestiaSinica(草地学报),2006,14(2):138-141(in Chinese).
[40] SALVUCCI M E,PORTIS J A R,OGREN W L.Purification of ribulose-1,5-bisphosphate carboxylase/oxygenase with high specific activity by fast protein liquid chromatography[J].AnalyticalBiochemistry,1986,153(1):97-101.
[41] DENG X(邓 雄),LI X M(李小明),ZHANG X M(张希明),etal.Relationship between gas exchange for desert plants and environmental factors in Taklimakan[J].ChineseJournalofApplicationEnvironmentalBiology(应用与环境生物学报),2002,8(5):445-452(in Chinese).
[42] WU T G(吴统贵),ZHOU H F(周和锋),WU M(吴 明),etal.Dynamics ofSalixmatsudanaphotosynthesis and its relations to environmental factors[J].ChineseJournalofEcology(生态学杂志),2008,27(12):2 056-2 061(in Chinese).
[43] XU H(许 皓),LI Y(李 彦),XIE J X(谢静霞),etal.Influence of solar radiation and groundwater table on carbon balance of phreatophytic desert shrubTamarix[J].ChineseJournalofPlantEcology(植物生态学报),2010,34(4):375-386(in Chinese).
