希金斯炭疽菌GPCR蛋白生物信息学分析
2015-03-21韩长志
韩 长 志
(西南林业大学 林学院, 云南省森林灾害预警与控制重点实验室, 昆明 650224)
希金斯炭疽菌GPCR蛋白生物信息学分析
韩 长 志*
(西南林业大学 林学院, 云南省森林灾害预警与控制重点实验室, 昆明 650224)
希金斯炭疽菌可以侵染诸多十字花科植物引起炭疽病,给各国农业生产造成了巨大经济损失.GPCR作为生物体内G蛋白信号转导途径中的重要感知蛋白,在信号传递过程中发挥着重要作用.本研究基于酿酒酵母中已经报道的3个典型GPCR序列,利用Blastp以及关键词对炭疽菌蛋白质数据库进行比对、搜索,以及通过TMHMM、HMMTOP跨膜结构域分析,明确该菌存在4个典型的GPCR;同时,通过对上述氨基酸序列进行细胞信号肽、亚细胞定位以及二级结构等生物信息学分析,明确上述GPCR均具有较高比例的α螺旋结构以及均不含有明显的信号肽序列;在定位方面,4个GPCR均定位在质膜上.此外,通过对希金斯炭疽菌中的4个GPCR与其他物种中的23个同源序列进行遗传关系比较分析,发现该菌中的GPCR与C.graminicola、C.fioriniae等炭疽菌属中的病菌具有较高的同源序列以及较近的亲缘关系.该研究为深入开展希金斯炭疽菌GPCR功能研究打下坚实的理论基础,同时,也为进一步开展其他炭疽菌的研究提供重要的理论指导.
希金斯炭疽菌; G蛋白偶联受体; 信号肽; 二级结构; 炭疽菌属
希金斯炭疽菌(ColletotrichumhigginsanumSacc.)可以侵染诸如小油菜、菜心、羽衣甘蓝、结球甘蓝、大白菜、萝卜等多种十字花科蔬菜植物而引起炭疽病[1-2],是一类重要的世界性植物真菌病害.该菌又称为希金斯刺盘孢,其主要分布于美国以及中国、日本、印度等东南亚国家[3].在我国,由该菌侵染菜心引起的炭疽病是菜心上最常见和发生最严重的病害之一[4].国内外对该病菌的研究主要集中在病菌的生物学特性、生防菌筛选以及遗传转化[5-6]、防治方法[7-8]等方面,随着该基因组序列的释放[9],目前,关于该菌MAPK途径蛋白预测[10]、致病基因鉴定[11-12]、基因功能[13-14]、分泌蛋白预测以及RGS[15]、14-3-3蛋白[16]、磷酸二酯酶[16]、septin[17]等蛋白生物信息学分析等已见报道,而关于G蛋白信号通路效应酶的报道尚不多见.GPCR作为G蛋白信号转导途径上接收外源信号、传递信号重要的表面受体,在植物与病原菌互作过程中发挥着诸多重要的作用[18-19].
本研究利用模式生物酿酒酵母SaccharomycescerevisiaeS288c中已经报道的3个典型GPCR氨基酸序列[20],通过在炭疽菌属蛋白质数据库中进行Blastp比对分析、关键词搜索,获得与酿酒酵母GPCR同源的C.higginsianum序列,选择TMHMM和HMMTOP对GPCR进行跨膜结构预测,并通过理化性质、疏水性分析、二级结构预测以及信号肽、亚细胞定位等生物信息学分析,以期明确该菌中所存在的GPCR数量、理化性质、结构特征以及定位情况,同时,基于上述发现的GPCR氨基酸序列,在美国国家生物信息中心(NCBI)在线进行同源序列搜索,通过遗传关系分析,以期为进一步开展同属于炭疽菌属但其基因组序列尚未公布的其他炭疽菌的研究提供重要的理论指导.
1 材料与方法
1.1 材料
利用关键词“G-protein coupled receptor”以及“GPCR”等对炭疽菌属蛋白数据库在线进行搜索,同时,以S.cerevisiaeS288c中3个GPCR(GCR1、STE2、STE3)蛋白序列为基础,进行Blastp进行比对分析(参数选择默认).另外,利用NCBI明确该菌中GPCR蛋白质登录号信息.
1.2 方法
1.2.1 保守结构域及跨膜区结构预测 利用SMART网站[21]在线分析GPCR所具有的保守结构域特征.利用TMHMM Server v. 2.0[22]和HMMTOP version 2.0[23]等跨膜网站对GPCR的跨膜区结构进行预测.
1.2.2 蛋白质理化性质分析 利用Protscale程序[24]对GPCR进行理化性质测定.
1.2.3 蛋白质信号肽预测 利用SignalP 3.0 Server[22]在线分析实现对蛋白质信号肽的预测.
1.2.4 蛋白质二级结构预测 采用PHD[25]在线分析实现对蛋白质二级结构预测.
1.2.5 亚细胞定位分析 对GPCR进行亚细胞定位分析,利用ProtComp v9.0实现(http://linux1.softberry.com/berry.phtml?topic=protcompan&group=programs&subgroup=proloc),以期获得蛋白质的定位情况.
1.2.6 系统进化树构建 在NCBI中,以C.higginsianum中GPCR氨基酸序列为基础,在线进行Blastp同源搜索,获得来自于不同物种的同源蛋白质序列.对所获得的同源序列,利用ClustalX[26]进行多重比对分析,随后利用MEGA 5.2.2软件[27]构建系统进化树:采用邻近法构建系统发育树,各分支之间的距离计算采用p-distance模型,系统可信度检测采用自举法重复1 000次进行.
2 结果与分析
2.1 希金斯炭疽菌4个GPCR均具有典型的七次跨膜结构域
通过同源比对搜索,结果显示,与S.cerevisiaeS288c中STE3、GPR1同源的C.higginsianumGPCR蛋白ID分别为CH063_13574.1、CH063_13120.1,并未发现与STE2同源的C.higginsianumGPCR蛋白;另外,通过关键词搜索,结果显示,共获得C.higginsianum中两个GPCR,其ID分别为CH063_00913.1、CH063_10797.1(表1).根据序列彼此之间的同源结果,将上述所获得的的GPCR蛋白分别命名为ChGPR1、ChGPR2、ChSTE3、ChGPR4.

表1 希金斯炭疽菌GPCR基本情况及获取方法
基于TMHMM跨膜结构域分析,结果显示,除ChSTE3外,其他GPCR均具有七次跨膜结构域(图1).同时,利用HMMTOP进行预测,与上述结果相同,而对于具体的跨膜起始位置、终止位置预测并不相同(表2).
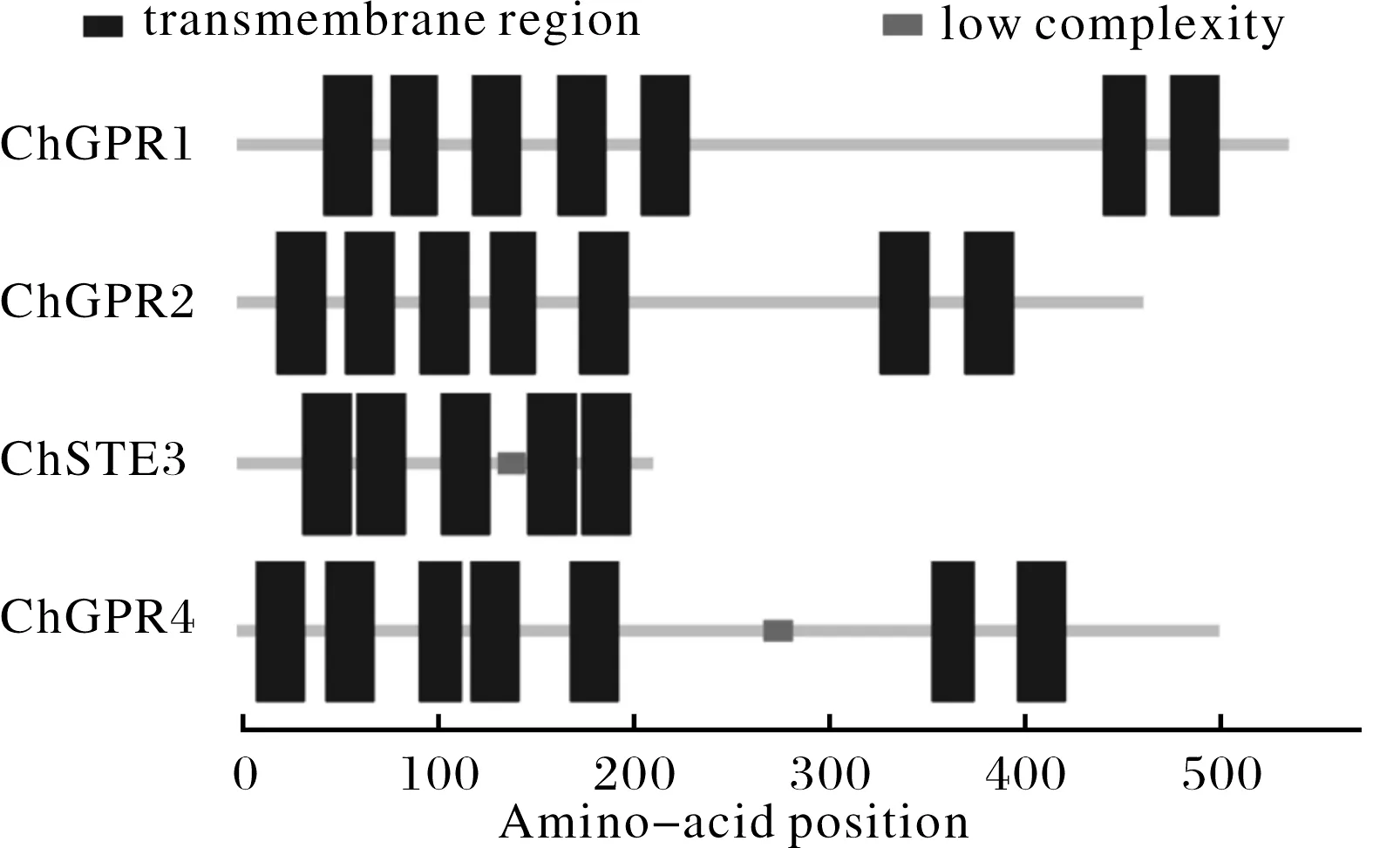
图1 希金斯炭疽菌GPCR的保守结构域分析Fig.1 The conserved domain of GPCRs in C. higginsianum
2.2 GPCR蛋白质理化性质及疏水性预测
C.higginsianum中所含GPCR彼此之间在酸性氨基酸、碱性氨基酸以及非极性R基氨基酸、不带电荷的极性R基氨基酸方面组成及所占比例方面均存在不同(表3),同时,在相对分子质量、理论等电点、负电荷氨基酸残基数、正电荷氨基酸残基数、分子式、原子质量以及不稳定性系数、脂肪族氨基酸指数、总平均亲水性等方面均存在着一定差异(表4).此外,除ChGPR4不稳定性系数小于40外,其他均大于40,属于不稳定蛋白;ChGPR1、ChGPR2、ChGPR4总平均亲水性(GRAVY)小于0,为亲水性蛋白,而CgSTE3为疏水性蛋白(表4).C.higginsianum中4个GPCR在亲(疏)水性最强氨基酸残基及位置方面也存在着较大的差异(数据未显示).
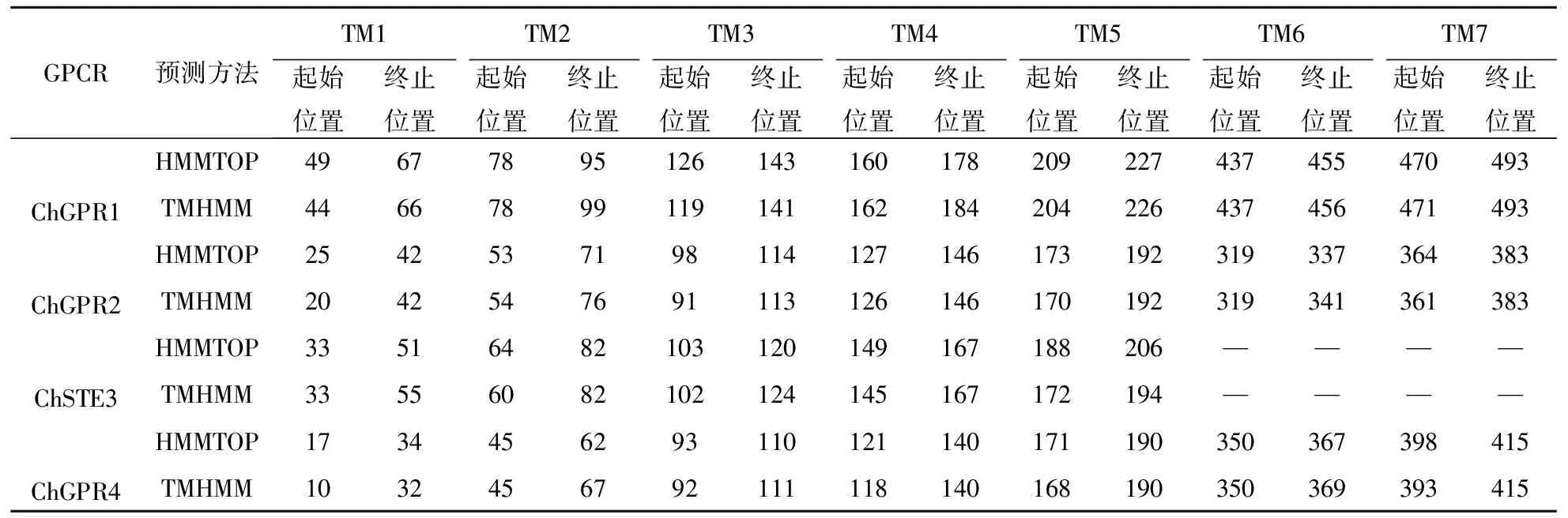
表2 希金斯炭疽菌GPCR跨膜情况预测
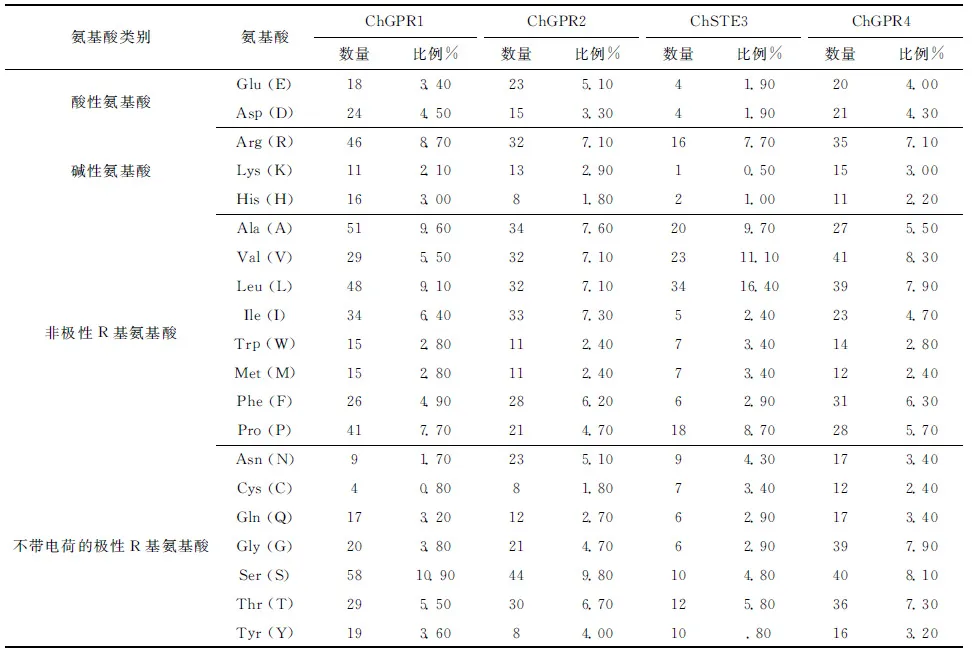
表3 希金斯炭疽菌GPCR氨基酸组成情况
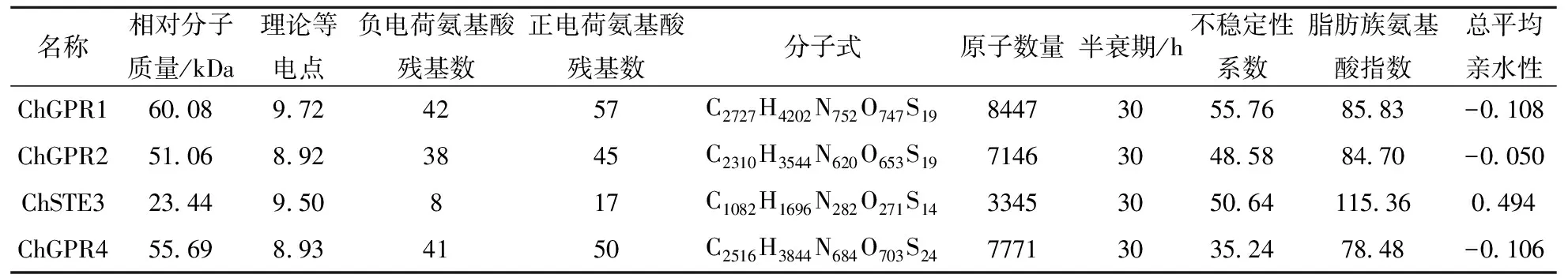
表4 希金斯炭疽菌GPCR蛋白质基本理化性质
2.3 信号肽特征
经过SingnalP 3.0分析,无论是经NN计算还是经HMM分析,4个GPCR均未发现有明显的信号肽序列(数据未显示).
2.4 二级结构预测
希金斯炭疽菌C.higginsianum中所含的4个GPCR均含有较高比例的α螺旋,除ChSTE3外,其他GPCR所含有的α螺旋、β卷曲以及无规卷曲所占比例较为相近(图2).
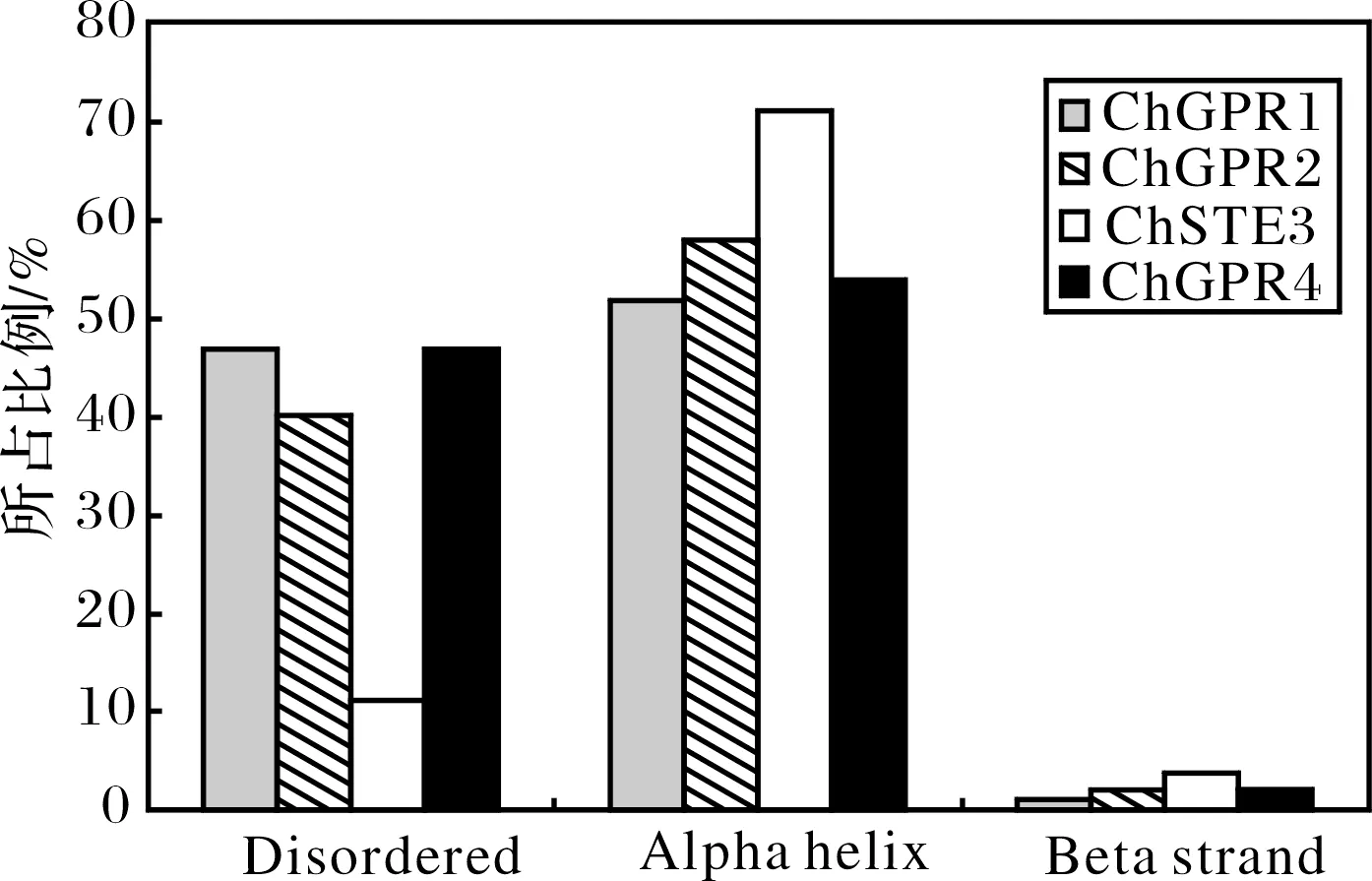
图2 希金斯炭疽菌GPCR的二级结构分析Fig.2 The secondary structure character of GPCRs in C. higginsianum
2.5 亚细胞定位分析
C.higginsianum中所含的4个GPCR亚细胞定位情况均为质膜(表5),该结果与通过SMART分析以及跨膜结构域分析结果一致,同时,也符合GPCR所具有的功能特征.
2.6 遗传关系
通过对C.higginsianum中的4个GPCR序列及其同源序列进行聚类分析,结果显示,分别以ChGPR1、ChGPR2、ChSTE3以及ChGPR4为核心,分为明显的4大类,就上述四个GPCR亲缘关系而言,ChGPR1、ChGPR2和ChGPR4彼此之间亲缘关系较近,而ChGPR2和ChGPR4彼此之间亲缘关系更近.同时,发现该菌中的GPCR与C.graminicola、C.fioriniae等炭疽菌属中的病菌具有较高的同源序列以及较近的亲缘关系(图3).
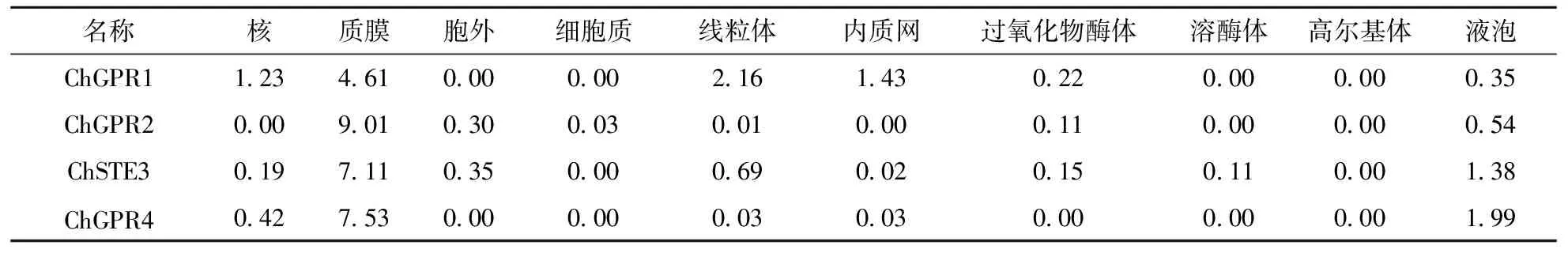
表5 希金斯炭疽菌GPCR亚细胞定位情况
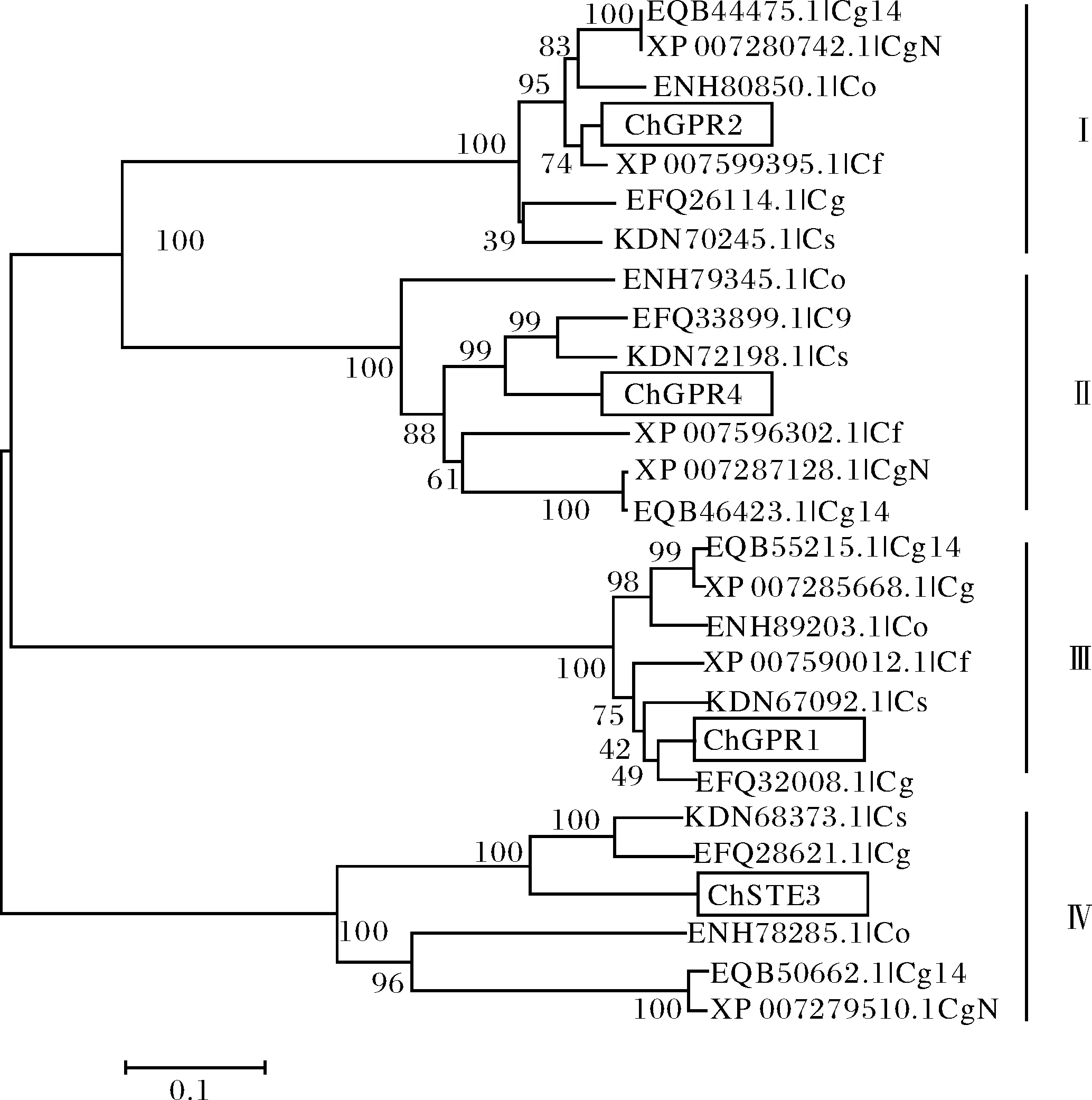
Cg、CgN、Cg14、Co、Cf、Cs分别为Colletotrichum graminicola、Colletotrichum gloeosporioides Nara gc5、Colletotrichum gloeosporioides Cg-14、Colletotrichum orbiculare MAFF 240422、Colletotrichum fioriniae PJ7、Colletotrichum sublineola等物种的缩写.
3 结论与讨论
前人研究发现,基于HMM算法的TMHMM和HMMTOP对GPCR预测具有非常好的评价准确性,正确率在85%[28].本研究通过对炭疽菌属蛋白数据库进行Blastp比对分析以及关键词搜索,并利用TMHMM、HMMTOP程序对所获得的候选GPCR进行跨膜结构域预测,明确C.higginsianum中存在4个GPCR,同时,利用SMART、SignalP、PHD、Protscale、TargetP等生物信息学分析网站,明确其保守结构域、信号肽、理化性质、疏水性、亚细胞定位等情况;此外,通过对上述GPCR及其同源序列进行遗传关系分析,明确彼此之间存在的亲缘关系.
通过对S.cerevisiaeS288c中GPR1、STE2、STE3保守结构域分析,结果显示GPR1仅具有六次跨膜结构域,其他STE2、STE3均具有典型的七次跨膜结构域,而本研究中所获得的4个GPCR中,除ChSTE3外,均具有典型的七次跨膜结构域,因此,七跨膜结构域是否是GPCR所具有的唯一典型特征有待于进一步试验验证.
[1] 梁惠凌, 唐 辉. 广西常见花卉真菌性病害的防治[J]. 广西园艺, 2002(2): 18-19.
[2] 沈瑞清, 张 萍, 郭成瑾, 等. 宁夏炭疽菌属真菌资源研究[J]. 河南农业科学, 2012(5): 100-102, 149.
[3] Hyde K, Cai L, Cannon P, et al. Colletotrichum-names in current use[J]. Fungal Diversity, 2009, (39): 147-182.
[4] 卢博彬, 杨 暹. 菜心炭疽病研究进展[J]. 长江蔬菜, 2009(24): 1-5.
[5] 周而勋, 杨 媚, 张 华, 等. 菜心炭疽病菌菌丝生长、产孢和孢子萌发的影响因素[J]. 南京农业大学学报, 2002(2): 47-51.
[6] 况福元, 吴小丽, 吕风青, 等. 菜心炭疽病菌拮抗细菌的筛选及鉴定[J]. 微生物学通报, 2009(9): 1350-1355.
[7] 郭巨先, 刘玉涛, 杨 暹. 钾营养对菜薹(菜心)炭疽病发生和植株防御酶活性的影响[J]. 中国蔬菜, 2012(14): 86-89.
[8] 康云艳, 周小萌, 杨 暹, 等. pHBA对菜心炭疽病的诱导抗性及植株生理特性的影响[J]. 植物病理学报, 2014, 44(4): 393-404.
[9] O’Connell R J, Thon M R, Hacquard S, et al. Lifestyle transitions in plant pathogenicColletotrichumfungi deciphered by genome and transcriptome analyses[J]. Nat Genet, 2012, 44(9): 1060-1065.
[10] 林春花, 蔡志英, 黄贵修. 全基因组法绘制禾谷炭疽菌和希金斯炭疽菌中MAPK级联信号途径简图[J]. 热带作物学报, 2012(4): 674-680.
[11] Huser A, Takahara H, Schmalenbach W, O’Connell R. Discovery of pathogenicity genes in the crucifer anthracnose fungusColletotrichumhigginsianum, using random insertional mutagenesis[J]. Molecular plant-microbe interactions, 2009, 22(2): 143-156.
[12] Ushimaru T, Terada H, Tsuboi K, et al. Development of an efficient gene targeting system inColletotrichumhigginsianumusing a non-homologous end-joining mutant andAgrobacteriumtumefaciens-mediatedgene transfer[J]. Molecular Genetics and Genomics, 2010, 284(5): 357-371.
[13] 周 鹏. 希金斯刺盘孢T-DNA插入体库的构建、筛选及相关突变体基因的克隆[D]. 武汉: 华中农业大学, 2011.
[14] 赵 典. 希金斯刺盘孢T-DNA插入体库的筛选及相关突变体基因的克隆[D]. 武汉: 华中农业大学, 2012.
[15] 韩长志. 希金斯炭疽菌RGS蛋白生物信息学分析[J]. 生物技术, 2014(1): 36-41.
[16] 韩长志. 希金斯炭疽菌14-3-3蛋白生物信息学分析[J]. 湖北农业科学, 2014, 3(15): 3669-3672.
[17] 韩长志. 希金斯炭疽菌中五个典型septin生物信息学分析[J]. 河南农业科学, 2014, 43(8): 91-96.
[18] Li L, Wright S J, Krystofova S, et al. Heterotrimeric G protein signaling in filamentous fungi[J]. Annu Rev Microbiol, 2007, 61:423-452.
[19] 韩长志. 植物病原丝状真菌G蛋白偶联受体的研究进展[J].微生物学通报, 2015, 42(2):374-383.
[20] Versele M, Lemaire K, Thevelein J M. Sex and sugar in yeast: two distinct GPCR systems[J]. EMBO reports, 2001, 2(7): 574-579.
[21] Letunic I, Doerks T, Bork P. SMART 7: recent updates to the protein domain annotation resource[J]. Nucleic acids research, 2012, 40(D1): D302-D305.
[22] Bendtsen J D, Nielsen H, von Heijne G, et al. Improved prediction of signal peptides: SignalP 3.0[J]. Journal of Molecular Biology, 2004, 340(4): 783-795.
[23] Tusnady G E, Simon I. The HMMTOP transmembrane topology prediction server[J]. Bioinformatics, 2001, 17(9): 849-850.
[24] Gasteiger E, Hoogland C, Gattiker A, et al. Protein identification and analysis tools on the ExPASy server[M]. The proteomics protocols handbook: Springer, 2005:571-607.
[25] Kelley L A, Sternberg M J. Protein structure prediction on the Web: a case study using the Phyre server[J]. Nat Protoc, 2009, 4(3): 363-371.
[26] Thompson J D, Gibson T J, Higgins D G. Multiple sequence alignment using ClustalW and ClustalX[J]. Current Protocols in Bioinformatics, 2002, Chapter 2(Unit 2.3).
[27] Tamura K, Peterson D, Peterson N, et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J]. Mol Biol Evol, 2011, 28(10): 2731-2739.
[28] Möller S, Croning M D, Apweiler R. Evaluation of methods for the prediction of membrane spanning regions[J]. Bioinformatics, 2001, 17(7): 646-653.
Bioinformatics analysis on G protein-coupled receptors inColletotrichumhigginsanum
HAN Changzhi
(College of Forestry, Southwest Forestry University, The Key Laboratory of Forest Disaster Warning and Control of Yunnan Province, Kunming 650224)
Colletotrichumhigginsanumcan infect cruciferous vegetables plants, causing tremendous economic losses in agricultural production in many countries. GPCR as a perception protein of G protein signal transduction in vivo plays an important role in the signal transduction process. Based on the three typical GPCR sequences had been reported inSaccharomycescerevisiae, to search GPCR-like protein sequence from the protein databases ofColletotrichumspp. with the Blastp as well as the use of keywords, and the seven trans-membrane domain of four typical GPCRs were analyzed by using the TMHMM. Meanwhile, through bioinformatics analysis including the signal peptide, subcellular location and the secondary structure, there is high proportion of helical structure and not contains significant signal peptide sequence in every GPCR protein, and which positioned in the plasma membrane. In addition, analysis of genetic relationships through comparative four GPCRs in C. graminicola with 23 homologous sequences in other species, bothC.graminicolaandC.fioriniaehave a high sequence homology and close genetic relationship withC.higginsanum. This study could provide strong theoretical foundation to the function of GPCR, and provided an important theoretical guidance to clarify the other pathogen inColletotrichumspp.
Colletotrichumhigginsanum; GPCR; signal peptide; secondary structure;Colletotrichumspp
2014-12-29.
云南省优势特色重点学科生物学一级学科建设项目(50097505);云南省高校林下生物资源保护及利用科技创新团队(2014015);云南省教育厅科学研究基金项目(2014Y330);国家自然科学基金青年科学基金项目(31200488).
1000-1190(2015)02-0246-06
S435.1
A
*E-mail: hanchangzhi2010@163.com.
