染色质解旋酶/ATP酶的DNA结合蛋白1样基因致瘤机制研究进展
2015-03-04综述审校
傅 点,王 玲,田 丰 (综述),程 文(审校)
(南京军区南京总医院泌尿外科,南京210002)
染色质解旋酶/ATP酶的DNA结合蛋白1样基因致瘤机制研究进展
傅点△,王玲,田丰 (综述),程文※(审校)
(南京军区南京总医院泌尿外科,南京210002)
摘要:染色质解旋酶/ATP酶的DNA结合蛋白1样基因(CHD1L)是一种新近被证实与在许多实体瘤中表达增多的致癌基因,它定位于第1号染色体q21区。CHD1L在肝细胞癌和其他肿瘤中的功能性研究提示,该基因在肿瘤形成过程中可以引起细胞增殖、调节G1/S过渡期并且可以抑制细胞凋亡。CHD1L活化的潜在机制可能是通过结合凋亡蛋白Nur77,或通过上调CHD1L调控的靶基因(如ARHGEF9、SPOCK1或TCTP)受体以激活蛋白激酶B通路,从而干扰细胞死亡程序。CHD1L目前已经被认为是一种新的可独立评价一些实体性肿瘤进展、预后以及生存率的生物标志物。现有的对CHD1L的认识在将来也许可以激励大家进行对一些特定肿瘤的靶向治疗研究。
关键词:染色质解旋酶/ATP酶的DNA结合蛋白1样基因;致癌基因;染色体1q21;扩增; ρ鸟嘌呤核苷酸交换因子9;蛋白聚糖1;Nur77
癌症是一种基因水平的疾病。国际合作的癌症基因组学研究已经发现许多肿瘤存在多种体细胞突变、染色体重排和结构变异[1]。大概需要3~6个基因改变即可使一个正常细胞变异为肿瘤细胞[2]。在某一特定肿瘤中,平均有2~8种体细胞变异,而其余的突变均无法提供选择性的生长优势,为无效突变。关键的基因变化(联合效应)通过改变正常细胞生长过程中的几个核心信号通路来影响细胞命运、细胞存活以及基因修护[1]。驱动基因目前被分为两种,一种是mut驱动基因,另一种是epi驱动基因。mut驱动基因包含了足够数量或种类的驱动基因突变,而epi驱动基因在肿瘤中有异常表达,但却常常不发生突变。它们通过DNA甲基化或核染色体的改变在肿瘤细胞分裂过程中发生变化[1]。通过传统的分子生物学或全基因组测序技术收集所有恶性转化(也被称为癌症起源[3])的因果关系,将找到战胜癌症的解决方案。肿瘤发生过程中的染色体重排已经发现了一种常见的基因异常(包括扩增、缺失或易位),它可能由一条或多条染色体的破碎引起,然后就是在修复时的碎片错接,接着就是kataegis突变模式的发生[4]。
染色体1q21的扩增是一个在许多实体肿瘤中最常见的遗传改变,包括膀胱癌[5]、乳腺癌[6]、鼻咽癌[7]、肝细胞癌[8]、食管癌[9]、骨纤维肉瘤[10]、结直肠癌[11]等。染色质解旋酶/ATP酶的DNA结合蛋白1样(chromodomain helicase/ATPase DNA binding protein 1-like gene,CHD1L)是最近发现的致癌基因,它定位于第1号染色体q21区,该区域是肝细胞癌中常见的扩增区域[12]。CHD1L在肿瘤发生过程中的致癌性和CHD1L蛋白在肿瘤中的过度表达,使它成为一种能够提示肿瘤预后差、无瘤生存时间短的生物标志物。通过了解更多关于CHD1L基因及其相关的分子机制,肿瘤发生的结构和功能来讨论开发CHD1L抑制剂作为治疗肿瘤潜在治疗策略的可能性。现就CHDIL致瘤机制的研究进展予以综述。
1CHD1L基因的结构
人类CHD1L基因,也被称为肝癌扩增基因1,由Ma等[12]于2008年第1次确定,它位于染色体1q21.1上,基因组坐标:146714292-146767443,strand (+)。CHD1L的基因组大小为53 152碱基对,包含23个外显子。其上游是含黄素的单加氧酶5(MFO5)基因,下游是前列腺素还原酶伪基因(LOC10030018)及长基因间的非编码RNA642 (LINC 00624)。其启动子包含可与转录因子xxxx结合的xxx模体。目前已发现有6种选择性剪接的变种基因。有趣的是,第6号变种是一个非编码转录变异体,这可以作为编码和非编码分子依赖于细胞的情况下编码RNA的一个例子。相应的编码蛋白的转录变异体在表1中列出。
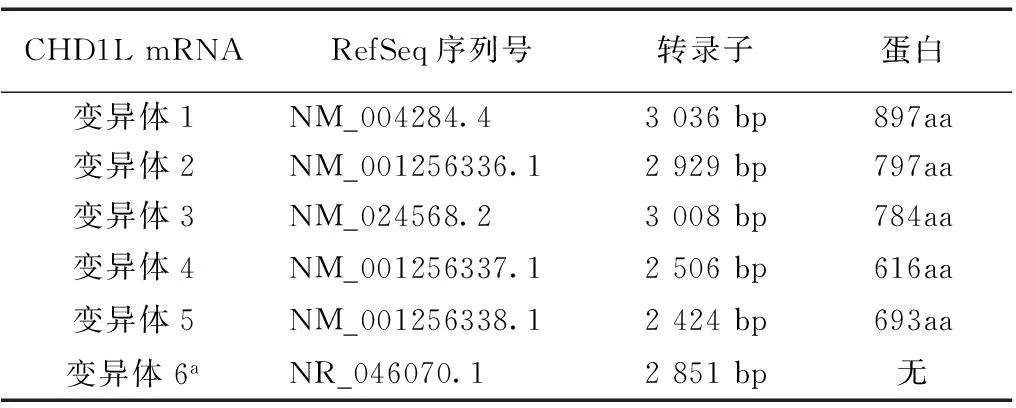
表1 CHD1L的转录变异体
GHD1L:染色质解旋酶/ATP酶的DNA结合蛋白1样基因;a为非编码转录
CHD1L信使RNA的全长为2980个碱基对(最近的数据为3036个碱基对),包含一个假定的开放阅读框,编码1个包含蛋白897aa[12]。序列分析表明,CHD1L属于SNF2家族,含有一个保守的SNF2-N结构域,一个解旋酶超家族结构域——解旋酶超家族C端结构域(helicase superfamily c-terminal domain,HELICc),以及一个宏结构域,见图1A[12]。该SNF2-N结构域是由280个氨基酸组成,并且ALC1的SNF2-N结构域与另一个SNF2样家族成员(染色质解旋酶DNA结合蛋白1,CHD1)间,两者存在序列同源性,其序列约有45%是相同的。CHD1L与CHD1中的HELICc结构域(包含107个氨基酸),其序列同源性约为59%[12]。因此,给这些基因起名为染色质解旋酶DNA结合蛋白1类似基因,即CHD1L。多种基因突变已在癌症的体细胞突变中有过报道 (http://cancer.sanger.ac.uk/cosmic/gene/analysis?ln=CHD1L#dist),有约56.67%为替代错义突变,见图1B。最近,在肾脏和泌尿系统先天性畸形患者中又检出了CHD1L突变[13]。各种不同的组织中均检出有CHD1L的表达,其表达的蛋白位于细胞核。
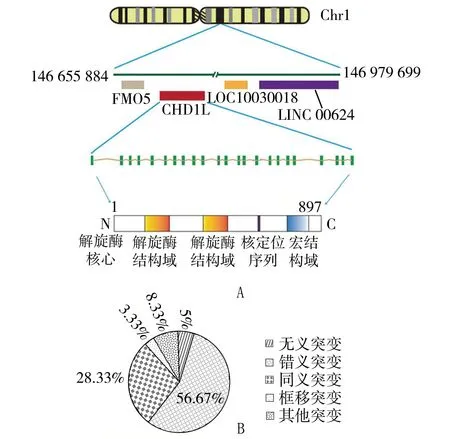
A:CHD1L全长转录(nm_004284.2)和所编码蛋白质的结构。CHD1L蛋白由N端的解旋酶结构域、C端的宏结构域以及核定位序列组成。B.CHD1L中突变种类的分布。CHD1L:染色质解旋酶/ATP酶的DNA结合蛋白1样基因;Chr1:1号染色体;FMO5:含黄素的单加氧酶5;LOC10030018:前列腺素还原酶伪基因;LINC 00624:长基因间的非编码RNA642图1 人类的CHD1L基因组信息
2CHD1L的功能
CHD1L是一种最近发现的致癌基因,其位于染色体1q21区,该区域常在肝细胞癌中扩增[12]。其蛋白质结构与CHD1类似,以此推测其具有与CHD1相近的生化功能。CHD1蛋白家族的特点是存在染色体结构域(染色质的结构修饰)和与SNF2相关的解旋酶/ATP酶结构域。
CHD1能够与DNA结合并调节ATP依赖的核小体组装,修改染色质结构并通过其固有的双染色质结构域和SNF2解旋酶/ATP酶结构域活化[14]。序列比较表明,CHD1L包含SNF2-N结构域和解旋酶超家族结构域。因此,CHD1L也被假设在转录调控、维持染色体的完整性和DNA修复中发挥着重要的作用。但是,与CHD1不同的是,CHD1L不含可识别甲基化组蛋白尾的染色质域。相反,CHD1L包含一个宏结构域,这是一个腺苷5′-去磷酸盐ADP-核糖/聚ADP核糖(poly-ADP-ribose,PAR)结合元件[15],因此,CHD1L具有PAR依赖的染色质重塑活性,并能在染色质中促进DNA修复[16]。CHD1L的ATP酶和染色质重塑活性受聚ADP核糖聚合酶PARP1和其底物NAD+调节,通过完整的宏结构域和染色质相关蛋白(包括组蛋白和PARP1)之间的短暂的相互作用而被强烈的激活[17]。CHD1L的这种核小体重塑活性取决于一个稳定的CHD1L PAR相关的PARP1-核小体中间体的形成[18]。此外,CHD1L所包含的C末端的宏观域(含600~897个残基)能够结合Nur77蛋白(该蛋白是一个非p53依赖的细胞凋亡通路的关键成员),并且抑制Nur77的核线粒体转位(这是Nur77介导的细胞凋亡的关键步骤),从而抑制细胞色素C的释放和阻断细胞凋亡的启动[19]。综上所述,染色质重塑因子CHD1L在哺乳动物发育的早期细胞分裂中起着重要的作用[20]。CHD1L赋予的DNA结合能力,通过染色质免疫为基础的克隆策略,来调节下游基因的表达。
ρ鸟嘌呤核苷酸交换因子9(Rho guanine nucleotide exchange factor 9,ARHGEF9)被确定为CHD1L的靶基因[21],它编码ρ小GTP酶Cdc42的特定的鸟嘌呤核苷酸交换因子。CHD1L蛋白直接结合到翻译控制肿瘤蛋白(translationally controlled tumor protein,TCTP)的启动子区(-1027~-733 nt)[22]和蛋白聚糖1(Sparc/osteonectin,cwcv,and kazal-like domains proteoglycan 1,SPOCK1)的启动子区(-1662~+ 34 nt)[23],随后激活这些靶基因的转录。这些CHD1L介导转录调控的上调目标可以部分解释CHD1L在癌症发展过程中的致癌作用机制。CHD1L通过与其他蛋白质的相互作用或调节其他基因的表达来执行其生物学效应。
3CHD1L与癌症
有许多文献报道,染色体1q21区的扩增在多种实体肿瘤中发现[5-7,9-11]。在肝细胞癌中,1q21区的扩增是最常见的遗传变异,占原发性肝癌的58%~78%[8]。这个发现驱使肿瘤生物学家去探索为什么这个区域会发生扩增,以及在这个区域中存在什么样的基因。2008年,Ma等[12]利用染色体显微切割/混合选择法第一次从扩增的1q21区中分离出了靶基因CHD1L。最近,有学者利用比较基因组杂交的方法进行高分辨率放大寡核苷酸序列分析,在尿路上皮癌中发现了包括CHD1L在内的一些存在于1q21~24区域内的扩增基因[24]。从肝癌研究中发现,CHD1L不仅被检测到通过荧光原位杂交扩增,而且其mRNA和蛋白也在检测样品中出现了过度表达[12]。CHD1L基因转染的细胞具有强烈的致癌能力,它能增加软琼脂集落形成并增加裸鼠成瘤性,这些效应能被抗CHD1L的小干扰RNA有效抑制[12]。为了进一步探讨在CHD1L体内的致癌作用,学者们建立了一个能够广泛表达CHD1L/ALC1的转基因小鼠模型。转基因小鼠中自发性肿瘤形成的概率为24.4%(10/41),包括4例肝细胞癌,但野生型小鼠中却没有发生。此外,CHD1L在肝细胞中的过度表达可能促进CHD1L转基因小鼠的肿瘤易患性[25]。CHD1L/ALC1在体外和体内肿瘤发生的致癌作用同样也在结肠癌中被观察到[11]。CHD1L同样也在用亚硝胺化学转化的人乳头瘤病毒感染的永生化宫颈细胞中发生表达[26]。这些证据有力地表明,CHD1L在癌症的发展中起到了一个驱动基因的作用。CHD1L扩增和过度表达的临床意义已经用于在实体性肿瘤的评价,包括肝细胞癌[27]、卵巢癌[28]、结肠癌[11]和膀胱癌[29]。这些均表明,CHD1L是一种新的生物标志物,可以用于预测肿瘤的进展和预后。另外,还有学者对CHD1L在肝癌患者的化疗反应中的作用进行了研究[30]。CHD1L能选择性地抑制5-氟尿嘧啶诱导的细胞凋亡,但无法抑制阿霉素。体外细胞培养和体内小鼠模型实验中均发现,化疗耐药表型可被抗CHD1L的短发卡RNA所逆转[30]。综合以上两个信息可以说明,CHD1L是一种新的致癌基因,可作为肿瘤预后差及化疗耐药的一个指标。综上所述,上述研究强烈支持CHD1L是一种新的癌基因,其在肿瘤发生中发挥着重要的作用。
4CHD1L驱动的致癌机制
驱动基因(mut驱动基因和epi驱动基因)赋予细胞选择性生长优势,可以分为12个信号通路,调节3个核心的细胞过程:细胞命运,细胞存活和染色体修复[1]。功能研究表明,CHD1L的过度表达能促进细胞增殖,促进G1/S期的过渡和抑制细胞凋亡[11-12]。在转基因小鼠模型中,CHD1L可以上调细胞周期蛋白A、周期蛋白 D1、周期蛋白 E、细胞周期蛋白依赖性激酶2和CDK4,下调成视网膜细胞瘤蛋白、细胞周期依赖性激酶特异性抑制蛋白p27(kip1)、p53,以此促进DNA合成和G1/S期过渡[25]。小鼠体外和体内的功能研究表明,CHD1L能增加细胞活力,并通过ARHGEF9介导的Cdc42的活化诱导丝状伪足的形成及上皮间质转化(epithelial mesenchymal transition,EMT),从而促进肿瘤细胞的移动、侵袭和转移。因此,CHD1L-ARHGEF9-Cdc42-EMT可能是肝癌进展和转移的一条新途径[21]。在人类肝癌样本中,有40.7%的样本检测出了TCTP,并与CHD1L的过度表达呈正相关。临床上,TCTP过表达与肿瘤分期和肝细胞癌患者的总体生存时间均显著相关(P<0.05)。在多变量分析中,TCTP被确定为一个与预后不佳有关的独立标志物。小鼠体外和体内的功能研究表明,TCTP具有致瘤能力,并且由CHD1L诱导的TCTP过表达可以导致肿瘤细胞的有丝分裂缺陷。进一步的机制研究表明,TCTP促进有丝分裂过程中的Cdc25C泛素-蛋白酶体降解,这导致CDK1的15位酪氨酸去磷酸化失败,从而降低了Cdk1的活性。结果就是有丝分裂过程中Cdk1活性的突然下降,诱导更快的退出有丝分裂过程并出现染色体分离现象,从而导致染色体的不稳定。耗竭试验证明,TCTP的致瘤性与它能导致有丝分裂缺陷的作用相关。总的来说,CHD1L-TCTP-Cdc25C-Cdk1是一种新型的分子途径,可以加速有丝分裂,产生异倍体标星,从而使肝细胞发生恶性转化[22]。SPOCK1通过激活蛋白激酶B抑制肝癌细胞凋亡,阻止细胞色素C、细胞凋亡胱天蛋白酶9(caspase-9)和caspase-3的活化释放。这些效应可被蛋白激酶B抑制剂所逆转。过度表达spock1的肝癌细胞中同样出现了高水平的基质金属蛋白酶9的表达,在基膜实验中表现出了更高的侵袭性,并且在免疫缺陷小鼠中,比对照的肝癌细胞形成了更多的转移性结节[23]。综上所述,CHD1L能激活细胞生存途径和抑制细胞程序性死亡信号,导致细胞命运的改变(出现恶性转化),见图2。
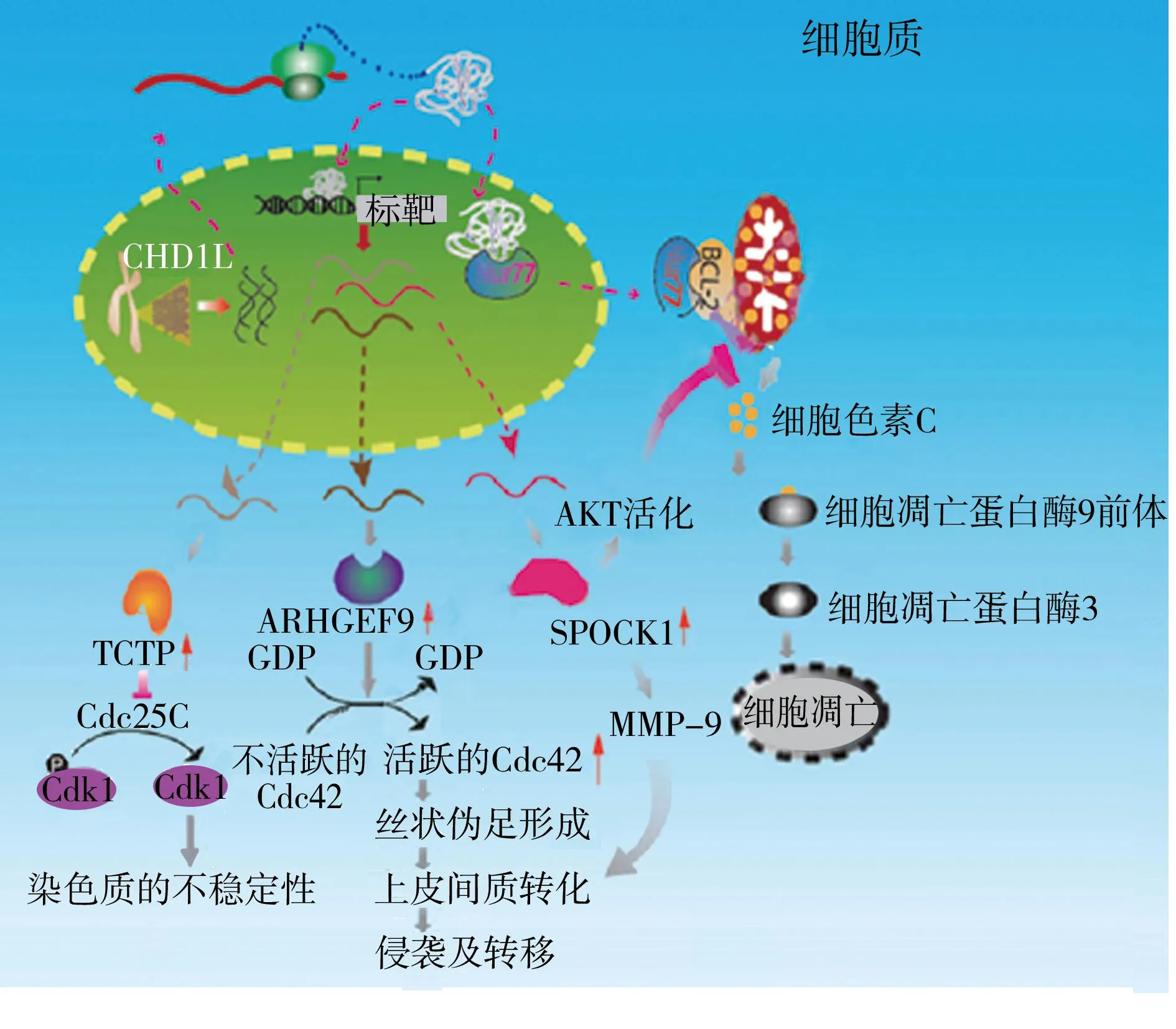
CHD1L:染色质解旋酶/ATP酶的DNA结合蛋白1样基因;ARHGEF9:ρ鸟嘌呤核苷酸交换因子9;SPOCK1:蛋白聚糖1;AKT:丝氨酸/苏氨酸蛋白激酶;TCTP:翻译控制肿瘤蛋白;GTP:三磷酸鸟苷;GDP:二磷酸鸟苷;MMP-9:基质金属蛋白酶9;Cdc:细胞分裂周期蛋白图2 CHD1L致癌作用的机制
5CHD1L的靶向治疗潜力
将癌症驱动基因转变为诊疗手段的成功范例促进了以CHD1L为靶向的潜在治疗研究。通过粉碎致癌基因及其编码产物来干扰其调控通路,以此来回复正常的细胞功能。首先,体外及体内实验数据表明,使用特定的RNA干扰分子,诱导细胞凋亡以改变癌细胞的行为后,CHD1L基因表达出现了显著的下调。用相应的shRNA使CHD1L表达沉默这一治疗方法,在肝癌治疗中具有巨大的治疗潜力,特别是能够增加联合5-氟尿嘧啶方案的化疗敏感性[30]。以siRNA 为基础的治疗是一种新兴的有前景的治疗方法。另一方面,由于CHD1L蛋白的宏结构域与多种配对物相互作用来执行其生物学效应,因此以宏结构域为靶向的治疗可能会提高放疗和化疗的有效性[31]。聚ADP核糖聚合酶1抑制剂和抑制Nur77结合的小分子可以反过来增加凋亡途径,或者通过抑制CHD1L的靶基因以使下游通路失活。理想状态下,组合所有这些策略可能有相加或协同作用。
6结论
由于CHD1L基因是从肿瘤中染色体1q21扩增子中分离出来的,功能性研究指出其在实体性肿瘤中具有致癌作用,特别是肝细胞癌。包含宏结构域的CHD1L独特的蛋白质结构与其他蛋白质一起执行多种生物学功能,如DNA损伤修复和抗细胞凋亡。此外,CHD1L介导的基因活化可能在肿瘤恶性转化中具有调节功能。更好地了解CHD1L基因组功能可能有益于研究癌症关键信号通路,从而为提出新的癌症治疗策略铺平道路。
参考文献
[1]Vogelstein B,Papadopoulos N,Velculescu VE,etal.Cancer genome landscapes[J].Science,2013,339(6127):1546-1558.
[2]Hahn WC,Counter CM,Lundberg AS,etal.Creation of human tumour cells with defined genetic elements[J].Nature,1999,400(6743):464-468.
[3]Wu W,Choudhry H.Next Generation Sequencing in Cancer Research-Decoding Cancer Genome,1st ed[M],New York:Springer,2013(1):199-215.
[4]Stephens PJ,Greenman CD,Fu B,etal.Massive genomic rearrangement acquired in a single catastrophic event during cancer development[J].Cell,2011,144(1):27-40.
[5]Simon R,Burger H,Brinkschmidt C,etal.Chromosomal aberrations associated with invasion in papillary superficial bladder cancer[J].J Pathol,1998,185(4):345-351.
[6]Tirkkonen M,Tanner M,Karhu R,etal.Molecular cytogenetics of primary breast cancer by CGH[J].Genes Chromosome Cancer,1998,21(3):177-184.
[7]Fang Y,Guan X,Guo Y,etal.Analysis of genetic alterations in primary nasopharyngeal carcinoma by comparative genomic hybridization[J].Genes Chromosome Cancer,2001,30(3):254-260.
[8]Guan XY,Fang Y,Sham J,etal.Recurrent chromosome alterations in hepatocellular carcinoma detected by comparative genomic hybridization[J].Genes Chromosome Cancer,2001,30(1):110.
[9]Kwong D,Lam A,Guan X,etal.Chromosomal aberrations in esophageal squamous cell carcinoma among Chinese:gain of 12p predicts poor prognosis after surgery[J].Hum Pathol,2004,35(3):309-316.
[10]Niini T,Lopez-Guerrero JA,Ninomiya S,etal.Frequent deletion of CDKN2A and recurrent coamplification of KIT,PDGFRA,and KDR in fibrosarcoma of bone--an array comparative genomic hybridization study[J].Genes Chromosome Cancer,2010,49(2):132-143.
[11]Ji X,Li J,Zhu L,etal.CHD1L promotes tumor progression and predicts survival in colorectal carcinoma[J].J Surg Res,2013,185(1):84-91.
[12]Ma NF,Hu L,Fung JM,etal.Isolation and characterization of a novel oncogene,amplified in liver cancer 1,within a commonly amplified region at 1q21 in hepatocellular carcinoma[J].Hepatology,2008,47(2):503-510.
[13]Brockschmidt A,Chung B,Weber S,etal.CHD1L:a new candidate gene for congenital anomalies of the kidneys and urinary tract (CAKUT)[J].Nephrol Dial Transplant,2012,27(6):2355-2364.
[14]Stokes DG,Perry RP.DNA-binding and chromatin localization properties of CHD1[J].Mol Cell Biol,1995,15(5):2745-2753.
[15]Karras GI,Kustatscher G,Buhecha HR,etal.The macro domain is an ADP-ribose binding module[J].EMBO J,2005,24(11):1911-1920.
[16]Ahel D,Horejsi Z,Wiechens N,etal.Poly(ADP-ribose)-dependent regulation of DNA repair by the chromatin remodeling enzyme ALC1[J].Science,2009,325(5945):1240-1243.
[17]Gottschalk AJ,Timinszky G,Kong SE,etal.Poly(ADP-ribosyl)ation directs recruitment and activation of an ATP-dependent chromatin remodeler[J].Proc Natl Acad Sci U S A,2009,106(33):13770-13774.
[18]Gottschalk AJ,Trivedi RD,Conaway JW,etal.Activation of the SNF2 family ATPase ALC1 by poly(ADP-ribose) in a stable ALC1.PARP1.nucleosome intermediate[J].J Biol Chem,2012,287(52):43527-43532.
[19]Chen L,Hu L,Chan TH,etal.Chromodomain helicase/adenosine triphosphatase DNA binding protein 1-like (CHD1l) gene suppresses the nucleus-to-mitochondria translocation of nur77 to sustain hepatocellular carcinoma cell survival[J].Hepatology,2009,50(1):122-129.
[20]Snider AC,Leong D,Wang QT,etal.The chromatin remodeling factor Chd1l is required in the preimplantation embryo[J].Biol Open,2013,2(2):121-131.
[21]Chen L,Chan TH,Yuan YF,etal.CHD1L promotes hepatocellular carcinoma progression and metastasis in mice and is associated with these processes in human patients[J].J Clin Invest,2010,120(4):1178-1191.
[22]Chan TH,Chen L,Liu M,etal.Translationally controlled tumor protein induces mitotic defects and chromosome missegregation in hepatocellular carcinoma development[J].Hepatology,2012,55(2):491-505.
[23]Li Y,Chen L,Chan TH,etal.SPOCK1 is regulated by CHD1L and blocks apoptosis and promotes HCC cell invasiveness and metastasis in mice[J].Gastroenterology,2013,144(1):179-191.
[24]Eriksson P,Aine M,Sjodahl G,etal.Detailed Analysis of Focal Chromosome Arm 1q and 6p Amplifications in Urothelial Carcinoma Reveals Complex Genomic Events on 1q,and as a Possible Auxiliary Target on 6p[J].PLoS One,2013,8(6):e67222.
[25]Chen M,Huang JD,Hu L,etal.Transgenic CHD1L expression in mouse induces spontaneous tumors[J].PLoS One,2009,4(8):e6727.
[26]Prokopczyk B,Sinha I,Trushin N,etal.Gene expression profiles in HPV-immortalized human cervical cells treated with the nicotine-derived carcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone[J].Chem Biol Interact,2009,177(3):173-180.
[27]Hyeon J,Ahn S,Park CK.CHD1L Is a Marker for Poor Prognosis of Hepatocellular Carcinoma after Surgical Resection[J].Korean J Pathol,2013,47(1):9-15.
[28]He WP,Zhou J,Cai MY,etal.CHD1L protein is overexpressed in human ovarian carcinomas and is a novel predictive biomarker for patients survival[J].BMC cancer,2012,10(12):437.
[29]Tian F,Xu F,Zhang ZY,etal.Expression of CHD1L in bladder cancer and its influence on prognosis and survival[J].Tumour Biol,2013,34(6):3687-3690.
[30]Chen L,Yuan YF,Li Y,etal.Clinical significance of CHD1L in hepatocellular carcinoma and therapeutic potentials of virus-mediated CHD1L depletion[J].Gut,2011,60(4):534-543.
[31]Han W,Li X,Fu X.The macro domain protein family:structure,functions,and their potential therapeutic implications[J].Mutat Res,2011,727(3):86-103.
A Review of CHD1L Tumorigenic MechanismFUDian,WANGLing,TIANFeng,CHENGWen.(DepartmentofUrology,NanjingMilitaryCommandNanjingGeneralHospital,Nanjing210002,China)
Abstract:Chromodomain helicase/ATPase DNA binding protein 1-like (CHD1L) gene is a newly identified oncogene located at 1q21 amplification in many solid tumors.The functional studies of CHD1L in hepatocellular carcinoma and other tumors strongly suggested its oncogenic role in tumoregenesis for unleashed cell proliferation,G1/S transition and inhibition of apoptosis.The underlying mechanisms of CHD1L activation may disrupt the cell death program via binding apoptotic protein Nur77 or through activation of AKT pathway by up-regulation of CHD1L-mediated target genes(such as ARHGEF9,SPOCK1 or TCTP).CHD1L is now considered to be a novel independent biomarker for progression,prognosis and survival in several solid tumors.The accumulated knowledge of CHD1L will invigorate the search for targeted treatment in specific subtype of tumors in the future.
Key words:Chromodomain helicase/ATPase DNA binding protein 1-like gene; Oncogene; Chr1q21; Amplification; Rho guanine nucleotide exchange factor 9; Sparc/osteonectin,cwcv,and kazal-like domains proteoglycan 1; Nur77
收稿日期:2014-08-04修回日期:2015-01-05编辑:薛惠文
doi:10.3969/j.issn.1006-2084.2015.14.025
中图分类号:R34
文献标识码:A
文章编号:1006-2084(2015)14-2562-04
