叶面喷施亚精胺对盐碱胁迫下番茄幼苗生长及其叶绿素合成前体含量的影响
2015-03-03徐志然胡晓辉胡立盼邹志荣潘雄波
张 丽,徐志然,胡晓辉,胡立盼,邹志荣,潘雄波
(西北农林科技大学 园艺学院,农业部西北设施园艺工程重点实验室,陕西杨陵 712100)
叶面喷施亚精胺对盐碱胁迫下番茄幼苗生长及其叶绿素合成前体含量的影响
张丽,徐志然,胡晓辉*,胡立盼,邹志荣,潘雄波
(西北农林科技大学 园艺学院,农业部西北设施园艺工程重点实验室,陕西杨陵 712100)
摘要:以盐碱敏感型番茄品种‘中杂9号’幼苗为试验材料,研究叶面喷施0.25 mmol·L-1亚精胺(Spd)对75 mmol·L-1盐碱溶液胁迫下番茄幼苗生长、净光合速率及叶绿素合成前体物质含量的影响,探讨Spd在缓解番茄盐碱胁迫伤害的生理机制。结果显示:(1)盐碱胁迫下番茄叶片叶绿素合成前体物质原卟啉Ⅸ(Proto Ⅸ)、镁-原卟啉Ⅸ(Mg-proto Ⅸ)、原叶绿素酸(Pchl)含量均显著降低,而δ-氨基酮戊酸(ALA)、胆色素原(PBG)、尿卟啉原Ⅲ(Uro Ⅲ)显著积累,Uro Ⅲ到Proto Ⅸ的转化受阻,引起叶片叶绿素a(Chl a)、叶绿素b(Chl b)和总叶绿素(Chl)含量显著降低,以及幼苗叶片净光合速率(Pn)、叶面积、叶片相对含水量、地上和地下部干鲜重的生长指标均显著降低。(2)盐碱胁迫下,叶面喷施Spd可显著促进番茄幼苗的生长,抑制叶片内ALA、PBG、Uro Ⅲ的积累,并提高Proto Ⅸ、Mg-proto Ⅸ、Pchl、Chl a、Chl b、Chl含量和相应Pn值。研究表明,盐碱胁迫显著抑制番茄幼苗的生长,叶面喷施Spd可通过缓解Uro Ⅲ到Proto Ⅸ的转化受阻程度,促进盐碱胁迫下番茄叶片的叶绿素合成,提高叶绿素含量和净光合速率,减轻盐碱胁迫对幼苗生长的伤害。
关键词:叶面喷施;亚精胺;番茄;盐碱胁迫;叶绿素合成前体物质
随着人类活动对环境破坏的加剧,日益严重的全球土壤盐碱化已成为一个世界性的环境、资源和生态问题,近年来引起了人们的普遍关注[1]。当前,全球盐碱地面积已达9.5×108hm2[2],中国的盐渍土总面积约3.6×107hm2,并且有9.2×107hm2的耕地正在面临盐碱化[3],主要分布在西北、华北及东北等地区,严重影响作物的产量、品质和效益。土壤盐碱化是限制植物生长和生产的重要因素,已经成为世界灌溉农业可持续发展的资源制约因素。
多胺(PAs)是植物体内普遍存在的一类具有强烈生物活性的低分子量脂肪族含氮碱,主要包括腐胺(Put)、亚精胺(Spd)和精胺(Spm),PAs通过促进生物大分子合成、稳定生物膜、清除活性氧自由基、影响跨膜质子电化学梯度及渗透调节等作用,参与植物体内许多重要的代谢过程及提高植物抗逆性反应[4]。近年来,Spd对植物抗盐性的影响机理研究日趋深入,但已有研究主要集中在NaCl上[5-6],而对碱性盐或混合盐碱逆境的相应研究报道相对较少。本课题组前期研究表明,外源Spd可提高盐碱胁迫下番茄幼苗的光合作用效率,减轻盐碱胁迫对番茄幼苗抑制效应,且对敏感品种的作用效果更为显著[4],但Spd对番茄等蔬菜叶绿素(Chl)合成调控的机理尚不清楚。因此,为进一步探讨Spd对盐碱胁迫下影响番茄幼苗叶绿素合成的机理,本试验以盐碱敏感的‘中杂9号’番茄为试验材料,采用水培的方式,研究叶面喷施外源Spd对盐碱胁迫下番茄幼苗生长及叶绿素合成前体物质含量的影响,旨在探讨Spd在缓解番茄盐碱胁迫伤害的生理机制,以期为进一步研究和利用Spd提供参考依据。
1材料和方法
1.1试验材料
试验于2013年4~11月在西北农林科技大学科研温室中进行,以‘中杂9号’(盐碱敏感型)番茄(Solanumlycopersicum)[4,7]为试验材料。
1.2试验处理
采用穴盘育苗,待番茄幼苗长至三叶一心时,选取长势一致健壮的幼苗定植于盛有1/2剂量Hoagland营养液的水培槽中预培养,用气泵通气。幼苗长至六叶一心时进行不同试验处理。试验共设4个处理:(1)对照(CK):1/2剂量Hoagland营养液+叶面喷施蒸馏水;(2)Spd处理(Spd):1/2剂量Hoagland营养液+叶面喷施0.25 mmol·L-1Spd[7];(3)盐碱处理(MS):75 mmol·L-1盐碱溶液(NaCl:Na2SO4:NaHCO3:Na2CO3=1:9:9:1)+叶面喷施蒸馏水;(4)盐碱+Spd处理(MS+Spd):75 mmol·L-1盐碱溶液+叶面喷施0.25 mmol·L-1Spd。每天18:00~18:30,叶片正反面均匀喷施Spd(喷施前加入2滴Tween-20)或等量的蒸馏水,以叶面产生水膜为止。处理第6天时选取幼苗生长点下第2片完全展开叶片测定叶绿素含量、净光合速率、叶绿素合成前体物质含量及生长指标。每处理48株,4次重复。
1.3测定指标与方法
1.3.1干重、鲜重、叶面积、叶片相对含水量选取各处理10株植株,用蒸馏水冲洗干净,吸水纸吸干表面水分称鲜重,再经115 ℃杀青15 min,75 ℃烘干至恒重,称干重;叶面积测定参照高建昌[8]的方法,用台式扫描仪(EPSON V700)扫描番茄叶片图像,然后用Image J软件进行叶面积分析;叶片相对含水量参照杨国会等[9]的方法测定。
1.3.2净光合速率采用Li-6400便携式光合仪(Li-Cor Inc,USA),上午9:00~11:30对各处理植株生长点下第2片完全展开功能叶的净光合速率进行测定。设定叶室温度为26 ℃,照射光强为800 μmol·m-2·s-1,CO2浓度为400 μmol·mol-1。
1.3.3叶绿素和叶绿素合成前体物质含量叶绿素含量测定参照张宪政[10]的方法。原卟啉Ⅸ(Proto Ⅸ)、镁-原卟啉Ⅸ(Mg-Proto Ⅸ)和原叶绿素酸酯(Pchl)含量测定按文献[11]进行;尿卟啉原Ⅲ(Uro Ⅲ)和胆色素原(PBG)含量按Bogorad[12]方法测定;δ-氨基酮戊酸(ALA)含量按文献[13]进行。
1.4数据处理和分析
试验数据采用Excel 2010绘图和SAS 8.1软件进行方差分析和Duncan’s多重比较。
2结果与分析
2.1叶面喷施Spd对盐碱胁迫下番茄幼苗生长的影响
由表1可知,正常栽培条件下,叶面喷施Spd处理显著提高了番茄幼苗的叶面积、地上部鲜重、干重和地下部干重;盐碱胁迫处理显著抑制番茄幼苗的生长,使番茄的叶面积、叶片相对含水量、地上部鲜重、地下部鲜重、地上部干重、地下部干重分别比对照降低了48.90%、19.89%、58.13%、56.18%、60.92%和37.50%;与单纯盐碱处理相比,盐碱胁迫下叶面喷施Spd处理的番茄幼苗的叶面积、相对含水量、地上部鲜重和干重、地下部鲜重和干重分别增加了20.33%、10.27%、28.21%、30%、44.07%和20%,且除地下部干重外,其余各指标均达到显著水平。表明叶面喷施Spd可有效缓解盐碱胁迫对“中杂9号”番茄幼苗生长的抑制作用。
2.2叶面喷施Spd对盐碱胁迫下番茄叶绿素含量的影响
表2显示,正常栽培条件下,叶面喷施Spd使番茄叶片总叶绿素和叶绿素a含量分别显著提高7.95%和8.56%,但对叶绿素b无明显提高;在处理第6天时,单纯盐碱处理的番茄叶片总叶绿素、叶绿素a、叶绿素b含量均比对照显著降低;与单纯盐碱胁迫相比,盐碱胁迫下叶面喷施Spd显著提高了番茄总叶绿素、叶绿素a、叶绿素b含量。表明叶面喷施Spd能有效降低盐碱胁迫对幼苗叶绿素的破坏作用,并且对叶绿素b的作用效果更显著。
2.3叶面喷施Spd对盐碱胁迫下番茄幼苗叶片叶绿素合成前体物质含量的影响
2.3.1ALA、PBG、Uro Ⅲ含量由图1可知,正常栽培下,叶面喷施Spd对番茄幼苗叶片叶绿素合成前体物质ALA和Uro Ⅲ含量无显著影响,而使PBG含量有所降低;盐碱胁迫下番茄叶片ALA、PBG、Uro Ⅲ含量较对照均有大幅度显著增加,增幅分别达47.05%、110.07%和150.42%;盐碱胁迫下喷施Spd显著降低了单纯盐碱胁迫引起的ALA、PBG、Uro Ⅲ累积,但仍显著高于对照。说明番茄幼苗叶片ALA、PBG、UroⅢ含量在盐胁迫逆境下显著增加,而喷施Spd能有效降低它们的积累量。
2.3.2Proto Ⅸ、Mg-proto Ⅸ、Pchl含量图1还显示,正常栽培条件下,叶面喷施Spd对番茄幼苗叶片叶绿素合成前体物质Proto Ⅸ、Mg-proto Ⅸ、Pchl含量也无显著影响;与对照处理相比,盐碱胁迫下番茄叶片的Proto Ⅸ、Mg-proto Ⅸ、Pchl含量显著降低,表明盐碱胁迫减弱了Proto Ⅸ、Mg-proto Ⅸ、Pchl的合成能力;与单纯盐碱胁迫相比,盐碱胁迫下叶面喷施Spd能显著提高Proto Ⅸ、Mg-protoⅨ和Pchl含量,增幅分别为11.31%、12.29%和12.64%,且达到与对照接近的水平,表明Spd能有效促进盐碱胁迫下番茄叶片叶绿素的合成。
2.4叶面喷施Spd对盐碱胁迫下番茄叶片净光合速率的影响
从图2可知,正常栽培条件下,叶面喷施Spd对番茄叶片的净光合速率Pn无显著影响;盐碱胁迫显著抑制了幼苗光合作用,番茄叶片Pn比对照显著降低了56.81%;同时,盐碱胁迫下叶面喷施Spd能显著提高番茄Pn,与单纯盐碱胁迫处理相比显著提高了43.37%,但仍低于对照水平。这表明盐碱胁迫下外源Spd可显著提高‘中杂9号’番茄幼苗的净光合速率,在一定程度上缓解盐碱胁迫对幼苗光合作用的抑制。
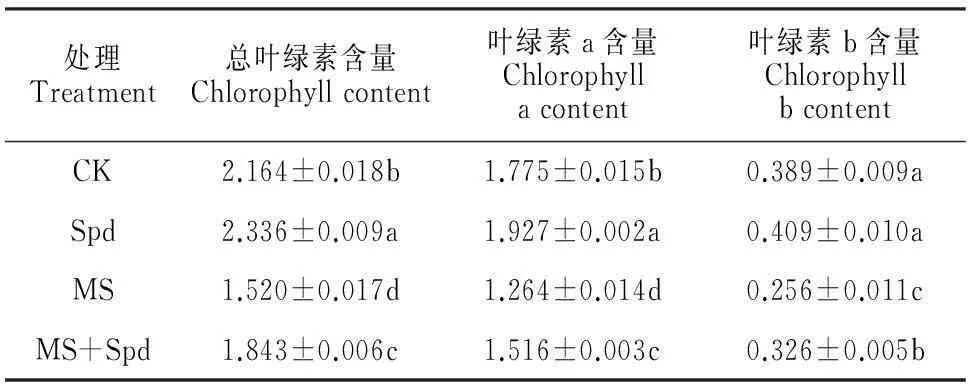
表2 叶面喷施Spd对盐碱胁迫下番茄叶绿素含量的影响

表1 叶面喷施Spd对盐碱胁迫下番茄幼苗生长的影响
注:CK.对照(1/2 Hoagland);Spd.叶面喷施亚精胺;MS.盐碱胁迫;MS+Spd.盐碱胁迫后叶面喷施亚精胺;表中同列不同字母表示处理间在0.05水平差异显著;数值为平均值±SE;下同。
Note:CK.Control(1/2 Hoagland);Spd.Foliar-spraying spermidine;MS.Saline-alkaline stress;MS+Spd.Foliar-spraying spermidine after saline-alkaline stress;Different letters in the same column meant significant difference among treatments at 0.05 level;Values were means±SE;The same as below.

图1 叶面喷施Spd对盐碱胁迫下番茄幼苗叶片叶绿素合成前体物质含量的影响
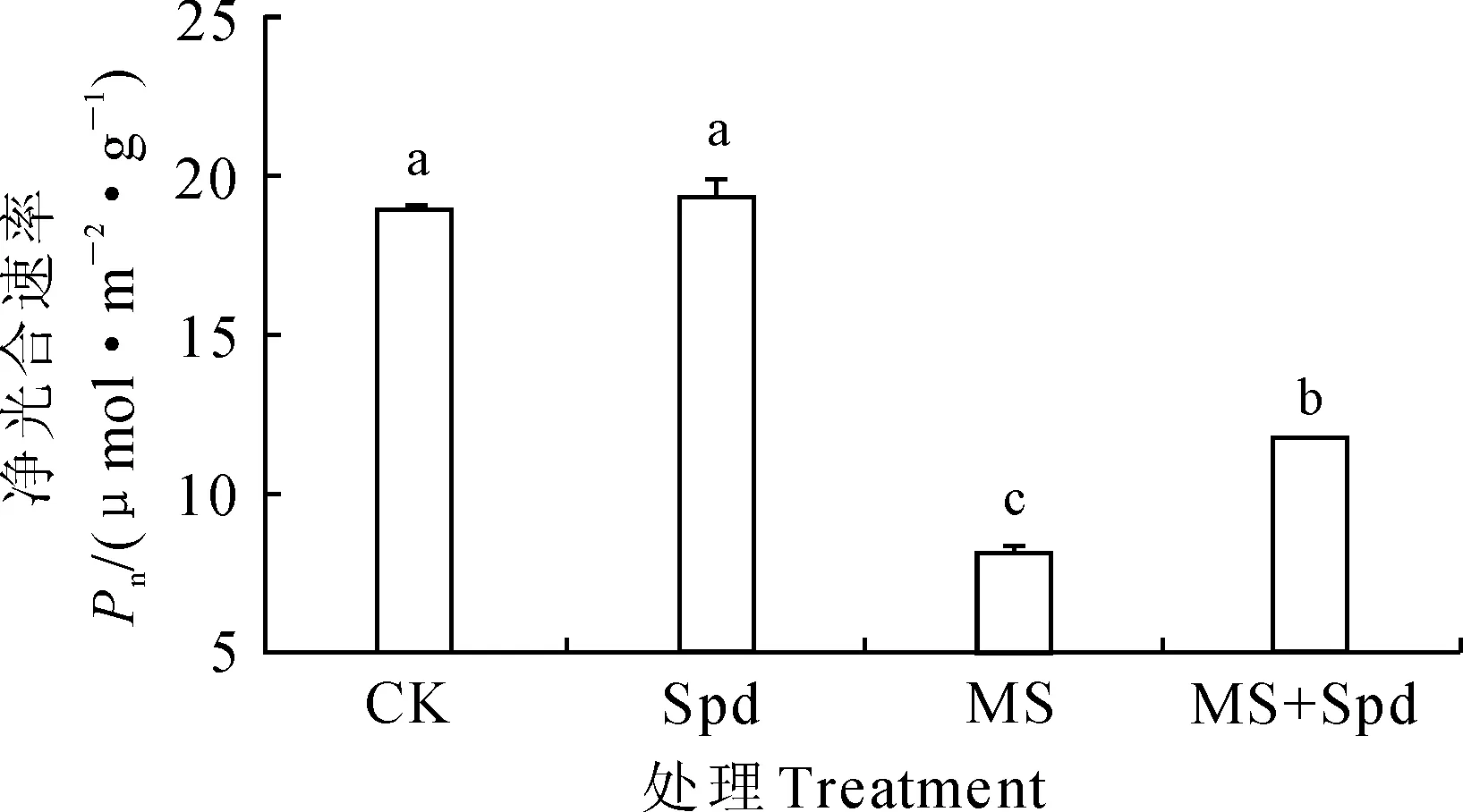
图2 叶面喷施Spd对盐碱胁迫下番茄
3讨论
生物量变化是植物对盐碱胁迫的综合反应,是评估胁迫程度和植物抗盐能力的可靠指标[14]。本研究表明番茄各项生长指标在盐碱处理6 d时均降低,其中番茄植株干重尤为显著;盐碱胁迫下,叶面喷施Spd处理能够显著改善番茄幼苗地上部的生长状况,缓解盐碱胁迫对地上部生物量积累的抑制作用,但对植株地下部生长的促进效应并不明显。
叶片光合色素含量是反映植物光合能力的一个重要指标,环境因子的改变会引起叶绿体色素含量的变化,进而引起光合性能的改变[15]。本研究结果表明,盐碱胁迫后,番茄幼苗叶片叶绿素含量显著降低,而叶面喷施Spd可以显著提高番茄幼苗的叶绿素含量,且对叶绿素b含量影响更为明显,可能是因为Spd处理后,一部分叶绿素a先与反应中心蛋白结合形成特殊状态下的叶绿素a,达到饱和后剩余的叶绿素a在脱植基叶绿素a加氧酶的作用下经过两步氧化反应最终生成叶绿素b,这与DA-6处理桃树叶片的结果一致[16]。本研究中盐碱胁迫下番茄叶绿素含量的高低与净光合速率高低变化趋势相一致,说明叶绿素含量的变化可影响净光合速率的变化,这与华春等[17]的研究结果相一致。Spd可通过参与膜构成阻止膜脂过氧化及膜蛋白的水解作用、稳定类囊体膜组成、阻止叶绿素损失[18],进而显著提高盐碱胁迫下番茄幼苗的净光合速率。
叶绿素的生物合成主要通过以下途径:谷氨酸(Glu)→δ-氨基酮戊酸(ALA)→胆色素原(PBG)→尿卟啉原Ⅲ(Uro Ⅲ)→原卟啉Ⅸ(Proto Ⅸ)→镁-原卟啉Ⅸ(Mg-proto Ⅸ)→原叶绿素酸(Pchl)→叶绿素a(Chl a)→叶绿素b(Chl b),Mg-proto Ⅸ形成之前的合成过程均在叶绿体基质进行,而后发生在类囊体膜上[13]。其中,任何一步出现障碍都会导致受阻碍部位以前的中间产物显著积累,受阻位点之后的产物则显著降低[19]。本试验结果表明,盐碱胁迫下叶绿素合成前体物质ALA、PBG、UroⅢ大量积累,而Proto Ⅸ、Mg-proto Ⅸ、Pchl含量显著降低,说明盐碱胁迫造成番茄叶绿素合成受阻发生在Uro Ⅲ向Proto Ⅸ的转化,这与郝树芹等[20]关于西葫芦银叶病发病叶片叶绿素合成受阻位点的研究结果相一致,发生此变化的原因可能与类囊体膜受损有关。叶绿素合成途径的很多酶属脂溶性,与叶绿体类囊体膜的发育密切相关[21]。多胺作为多聚阳离子,生理pH条件下带正电荷可与带电荷的磷酸基团、蛋白质及核酸等相互作用,稳定类囊体组成并参与膜构建[22]。本试验中,盐碱胁迫下,叶面喷施Spd降低了Uro Ⅲ的积累,促进了Proto Ⅸ向叶绿素的合成,提高了净光合速率,有效缓解盐碱胁迫对Uro Ⅲ到Proto Ⅸ转化的阻碍作用,这可能是Spd有助于稳定逆境胁迫下叶绿体类囊体膜的发育[23],从而保证了叶绿素合成由Uro Ⅲ到Proto Ⅸ转化过程中有充足的酶参与催化,促进了叶绿素的合成。但Spd对盐碱胁迫下番茄幼苗叶绿体结构以及合成关键酶的影响仍需要进一步深入研究。
综上所述,盐碱胁迫阻碍番茄幼苗叶片叶绿素合成中间产物Uro Ⅲ到Proto Ⅸ的转化,影响叶绿素的合成,造成叶绿素含量降低,进而降低其净光合速率,抑制植株生长。叶面喷施Spd能有效缓解盐碱胁迫下叶绿素合成步骤中Uro Ⅲ向Proto Ⅸ的受阻程度,促进其合成转化,提高叶绿素含量。
参考文献:
[1]JIA N E·A H(贾娜尔·阿汗),YANG CH W(杨春武),SHI D CH(石德成),etal.Physiological response of an alkali resistant halophyteKochiasieversianato salt and alkali stresses[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2007,27(1):79-84(in Chinese).
[2]WANG Y CH(王宇超),WANG D X(王得祥),PENG SH B(彭少兵),etal.Effects of salt stress on cell membrane permeability and physiological property in woody saltbush[J].AgriculturalResearchintheAridAreas(干旱地区农业研究),2007,25(4):225-229(in Chinese).
[3]ZHANG H H(张会慧),ZHANG X L(张秀丽),LI X(李鑫),etal.Effects of NaCl and Na2CO3stresses on the growth and photosynthesis characteristics ofMorusalbaseedlings[J].ChineseJournalofAppliedEcology(应用生态学报),2012,23(3):625-631(in Chinese).
[4]HU X H,ZHANG Y,SHI Y,etal.Effect of exogenous spermidine on polyamine content and metabolism in tomato exposed to salinity-alkalinity mixed stress[J].PlantPhysiologyandBiochemistry,2012,57:200-209.
[5]DUAN J J,LI J,GUO S R,etal.Exogenous spermidine affects polyamine metabolism in salinity-stressedCucumissativusroots and enhances short-term salinity tolerance[J].JournalofPlantPhysiology,2008,165(15):1 620-1 635.
[6]ROYCHOUDHURY A,BASU S,SENGUPTA D N.Amelioration of salinity stress by exogenously applied spermidine or spermine in three varieties of indica rice differing in their level of salt tolerance[J].JournalofPlantPhysiology,2011,168(4):317-328.
[7]LI J(李俊),LI J M(李建明),HU X H(胡晓辉),etal.Effects of pre-soaked seeds with spermidine on tomato seedling salinity-alkalinity tolerance[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2012,32(9):1 788-1 795(in Chinese).
[8]GAO J CH(高建昌),GUO G J(郭广君),GUO Y M(国艳梅),etal.Measuring plant leaf area by scanner and Image J Software[J].ChinaVegetables(中国蔬菜),2011,(2):73-77(in Chinese).
[9]YANG G H(杨国会),SHI D CH(石德成).Effects of NaCl stresses on relative water contents and defense enzymes activities in leaves ofGlycyrrhizauralensisFisch[J].JournalofHenanAgriculturalSciences(河南农业科学),2009,(12):104-106(in Chinese).
[10]ZHANG X ZH(张宪政).The Determination of chlorophyll content in plant-the method that use mixture containing acetone and ethanol[J].LiaoningAgriculturalSciences(辽宁农业科学),1986,(3):26-32(in Chinese).
[11]HODGINS R R,VAN HUYSTEE R B.Rapid simultaneous estimation of protoporphyrin and Mg-protophyrinsin higherplants[J].JournalofPlantPhysiology,1986,125(3-4):311-323.
[12]BOGORAD L.Porphyrin synthesis [M]//DARON H H,GUNSALUS I C.Methods in Enzymology.New York:New York Academic Press,1962:885-895.
[13]YU M(喻敏),HU CH X(胡承孝),WANG Y H(王运华).Effects of molybdenum on the precursors of chlorophyll biosynthesis in winter wheat cultivars under low temperature[J].ScientiaAgriculturaSinica(中国农业科学),2006,39(4):702-708(in Chinese).
[14]HE L(何磊),LU ZH H(陆兆华),GUAN B(管博),etal.Seed germination and seedling growth of the two sorghums under saline-alkaline stress[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2012,32(2):362-369(in Chinese).
[15]WANG S P(王素平),GUO SH R(郭世荣),HU X H(胡晓辉),etal.Effects of NaCl stress on the content of photosynthetic pigments in the leaves of cucumber(CucumissativusL.) seedlings[J].ActaAgriculturaeUniversitatisJiangxiensis(江西农业大学学报),2006,28(1):32-38(in Chinese).
[16]YANG Q(杨清),AI S J M M T(艾沙江 · 买买提),WANG ZH X(王志霞),etal.Effects of DA-6 on chlorophyll biosythesis pathway in peach leaves[J].ActaHorticulturaeSinica(园艺学报),2012,39(4):621-628(in Chinese).
[17]HUA CH(华春),ZHOU F(周峰),DING CH X(丁春霞),etal.Effects of exogenous spermidine on photosynthetic parameters and chloroplast ultrastructure ofSalicorniabigeloviiunder NaCl stress[J].JournalofPlantResourcesandEnvironment(植物资源与环境学报),2012,21(2):89-95(in Chinese).
[18]ZHANG CH M(张春梅),ZOU ZH R(邹志荣),HUANG ZH(黄志),etal.Effects of exogenous spermidine on photosynthesis of tomato seedlings under drought stress[J].AgriculturalResearchintheAridAreas(干旱地区农业研究),2010,28(3):182-187(in Chinese).
[19]LÜ M(吕明),LIU H H(刘海衡),MAO H D(毛虎德),etal.Changes of chlorophyll synthesis metabolismin chlorophyll-deficient mutant inBrassicajuncea[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2010,30(11):2 177-2 183(in Chinese).
[20]HAO SH Q(郝树芹),LIU SH Q(刘世琦),ZHANG Z K(张自坤),etal.Characteristics of chlorophyll metabolism and chlorophyll fluorescence in the silvered leaf of summer squash[J].ActaHorticulturaeSinica(园艺学报),2009,36(6):879-884(in Chinese).
[21]喻敏.冬小麦不同基因型的钼效率及其生理基础[D].武汉:华中农业大学,2000.
[22]TIAN J(田婧),GUO SH R(郭世荣),SUN J(孙锦),etal.Effects of exogenous spermidine on lipid peroxidation and membrane proton pump activity of cucumber seedling leaves under high temperature stress[J].ChineseJournalofAppliedEcology(应用生态学报),2011,22(12):3 252-3 258(in Chinese).
[23]XUE SH Y(薛淑媛),ZHU SH D(朱世东),LI X(李雪),etal.Effects of exogenous spermidine on photosynthetic characteristics and ultrastructure of salt-stressed muskmelon seedlings[J].JiangsuJournalofAgriculturalSciences(江苏农业学报),2013,29(3):613-618(in Chinese).
(编辑:裴阿卫)
Introduction of the Plant Front Cover:PetrocosmeaqinlingensisW.T.Wang
PetrocosmeaqinlingensisW.T.Wang belongs to the Family Gesneriaceae.Among all the species ofPetrocosmea,P.qinlingensisis on the northest edge of the distribution area of the genus.It is a perennial herbaceous plant with 7-12 leaves on each individual.Petioles short or long.Leaf blade papyraceous,wide ovate,rhombic or nearly rotund,0.7-3 cm in length,0.7-2.8 cm in width.Apex rotund or truncate.Basal part cuneate.Margin repand or unobvious creanate.Pubrulent on both side.Inflorecence 2-6.1 flower on each inflorescence.Calyx 5 lobed.Pubescent on absixal surface.Corolla labiate,light purple.Sparsely pubescent on out surface.Pilose on inner surface.Corolla tube about 2.8mm.Upper lobe 4.8mm in length and 2 divided.Lower lobes almost the same length as the upper ones,3 divided.Lobes oblong.Apex rotund.Fertile stamens 2,nearly basal.Pistil about 5mm long.Ovary and style villous.Stigma small and spherical.Flowering period August to September.The nearly related species,P.sinensisOliv.,is differed fromP.qinlingensisin having smooth inner surface of corolla.
The species was first descripted by the famous taxonomist,Wang Wencai,in 1981.The type specimen was collected from Mianxian County,Shaanxi Province.In the following 30 years,people tried to look for the wild population of the species but failed.Until recently,when carrying out the program ‘Second Investigation of wild plant resources in Shaanxi Province’,P.qinlingensiswas rediscovered near the location where it has been first found.
P.qinlingensisis an endemic species in Qinling Mountains area.It is usually found on moist rock in deciduous forest valley.The plant is a rare species and its distribution area is very small.It has been listed in the first group of national endangered wild plant species (class Ⅱ).
(Photographed and introduced by WANG Yong,YANG Peijun,LI Changbo and FAN Rong)
Effect of Foliar-spraying Spermidine on Seedlings Growth and
Contents of Chlorophyll Biosynthesis Precursors in
Leaves of Tomato under Saline-alkaline Stress
ZHANG Li,XU Zhiran,HU Xiaohui*,HU Lipan,ZOU Zhirong,PAN Xiongbo
(College of Horticulture,Northwest A&F University,Key Laboratory of Horticultural Engineering in Northwest China,Ministry of Agriculture,Yangling,Shaanxi 712100,China)
Abstract:With the salinity-alkalinity-sensitive tomato cultivar ‘Zhongza No.9’ as the experimental material,the effects of foliar-spraying Spd (0.25 mmol·L-1) on the growth paramaters,the net photosynthetic rate (Pn) and contents of chlorophyll biosynthesis precursors in tomato seedlings grown hydroponically under 75 mmol·L-1mixed saline-alkaline solution were studied.The results showed that:(1)Under saline-alkaline stress,the contents of chlorophyll biosynthesis precursors-protoporphyrin Ⅸ(Proto Ⅸ),Mg-Protoporphyrin Ⅸ(Mg-Proto Ⅸ),protochlorophyll (Pchl) were significantly decreased,while the contents of δ-aminolevulinic acid (ALA),porphobilinogen(PBG),uroorphyrinogen Ⅲ(Uro Ⅲ) were significantly accumulated in leaves of tomato seedlings.It was showed that stress restrained the transformation obstruction from Uro Ⅲ to Proto Ⅸ and lead to decrease in contents of Chl a,Chl b and total Chl,and reduced thePn,leaf area,leaf relative water content,fresh and dry weight of shoot and root.(2)Under saline-alkaline stress,foliar-spraying Spd could effectively promote tomato seedlings growth and suppress the accumulation of ALA,PBG,URO Ⅲ and also increase contents of Proto Ⅸ,Mg-proto Ⅸ and Pchl,and enhance thePnvalue.These results indicated that exogenous Spd could significantly relieve the degree of transformation obstruction from Uro Ⅲ to Proto Ⅸ,promote the tomato leaf chlorophyll synthesis and increase the chlorophyll content and net photosynthetic rate and also alleviate the stress damage under saline-alkaline stress.
Key words:foliar-spraying;spermidine;tomato;saline-alkaline stress;chlorophyll synthesis precursor
中图分类号:Q945.78
文献标志码:A
作者简介:张丽(1989-),女,在读硕士研究生,主要从事设施园艺研究。E-mail:precious0028@126.com*通信作者:胡晓辉,博士,副教授,主要从事设施园艺研究。E-mail:hxh1977@163.com
基金项目:国家“十二五”科技支撑计划(2011BAD29B01);国家星火计划(2012GA850001);西北农林科技大学校基金(QN2013018)
收稿日期:2014-07-02;修改稿收到日期:2014-10-06
文章编号:1000-4025(2015)01-0125-06
doi:10.7606/j.issn.1000-4025.2015.01.0125
