胡萝卜低温胁迫转录因子DcICE1基因克隆与非生物逆境响应分析
2015-03-03徐志胜李梦瑶熊爱生
黄 莹,徐志胜,王 枫,李梦瑶,马 静,熊爱生
(南京农业大学 作物遗传与种质创新国家重点实验室,园艺学院,农业部华东地区园艺作物生物学与种质创制重点实验室,南京 210095)
胡萝卜低温胁迫转录因子DcICE1基因克隆与非生物逆境响应分析
黄莹,徐志胜,王枫,李梦瑶,马静,熊爱生*
(南京农业大学 作物遗传与种质创新国家重点实验室,园艺学院,农业部华东地区园艺作物生物学与种质创制重点实验室,南京 210095)
摘要:植物ICE1基因是调控CBF基因表达的上游调控因子,在植物抵抗逆境胁迫中具有重要的作用。该实验以2个胡萝卜品种 ‘黑田五寸’和‘君川红’为实验材料,分别克隆出DcICE1转录因子基因,并通过荧光定量PCR方法测定了4种不同逆境胁迫下(4 ℃低温、38 ℃高温、0.2 mol·L-1NaCl及200 g·L-1PEG)DcICE1基因的表达情况,探讨DcICE1转录因子在植物抵抗非生物胁迫下的功能。序列分析显示,该基因全长1 458 bp,编码485个氨基酸。2个胡萝卜品种的DcICE1基因在核苷酸水平上有2个位点的差异,分别为第139位的G/A和第475位的A/G,导致编码的氨基酸在第47位的E/K和第159位的N/D差异,该差异可能与DcICE1转录因子在2个不同品种应对逆境胁迫下的响应不同有关。胡萝卜DcICE1转录因子有一个保守的ICE1功能结构域。实时定量PCR检测DcICE1基因在不同逆境胁迫下的响应表明,低温(4 ℃)处理能明显地诱导DcICE1基因的表达,但盐(0.2 mol·L-1NaCl)和干旱(200 g·L-1PEG)处理下的诱导效果均不明显。
关键词:胡萝卜;DcICE1转录因子;基因克隆;非生物胁迫;表达分析
低温是一类重要的环境因子,在很大程度上影响着植物的生长、发育,并且会给植物造成不同程度的伤害,严重影响着植物的产量与品质[1-2]。低温胁迫时,植物体内会发生许多生理生化和分子变化[3],包括细胞水平和基因水平[3-4]。因此,研究植物的抗寒性、探究植物的抗寒性机制,对提高植物的抗寒能力,提高农作物的产量和品质具有重要意义[5]。
在遭遇低温胁迫时,植物体内与冷有关的基因表达会被激活,从而提高植物的抗寒性,这种现象称为冷驯化,其中由冷胁迫诱导而表达的基因称为冷诱导基因(cold regulated gene,COR)[6]。冷诱导基因是一类编码调控性蛋白的基因,可调控相关抗寒基因的表达及相关抗寒蛋白的活性,如CBF/DREB、ICE基因[7]。ICE是在低温胁迫下诱导CBF家族转录因子基因表达的转录激活因子,在低温胁迫时能特定地结合CBF的启动子序列,诱导CBF基因表达,而CBF基因同样会诱导下游相关基因的表达,从而产生一系列适应低温的变化,增强植物的抗寒性[8]。ICE1基因编码的蛋白具有MYC转录因子家族特有的bHLH(碱性-螺旋-环-螺旋)DNA结合域,能够与CBF3启动子上的MYC顺式作用元件结合,激活CBF3基因的表达,提高抗寒性[8]。目前已经从许多植物中发现了ICE基因,包括拟南芥[9]、赤桉[10]、小麦[11]、油菜[5]、萝卜[7]、茶树[12]、苹果[13]、蝴蝶兰[14]、菊花[15],并且获得了转ICE基因的拟南芥[9,16]、黄瓜[17]、柠檬[18]、水稻[19]、烟草[20]和马铃薯[21],转基因植株抗寒性均有提高。
胡萝卜(DaucuscarotaL.)属于伞形科二年生草本植物,栽培面积非常广泛,主要以肉质根做食用材料。其肉质根含有丰富的营养物质,如葡萄糖、胡萝卜素、维生素等,在人们健康生活中发挥着重要的作用。胡萝卜生长发育大致可以分为发芽期、幼苗期、莲座叶生长期、肉质根膨大期以及抽薹开花期[22],且其生长发育需要一定的温度条件,播苗期温度太低,会影响出苗率以及幼苗生长;若在肉质根膨大期遭遇低温,会使胡萝卜提前经历春化作用而抽薹开花,严重影响胡萝卜的产量及品质[23-24]。近年来许多地区初春及秋冬季不乏极端天气的出现,严重影响胡萝卜的生长发育,给胡萝卜生产带来极大的损失。胡萝卜品种有多种,根据肉质根形状,可分为短圆锥形、长圆锥形和圆柱形三大类型,‘黑田五寸’和‘君川红’ 2个品种所食用肉质根分别为圆锥、圆柱类型,‘黑田五寸’品种引自日本,属中早熟品种,抗热抗病性强,产量高;‘君川红’品种属早熟品种,抗病性强,抗寒、抗热性好。‘黑田五寸’和‘君川红’ 均有很高的商品性,是生产中常用的胡萝卜栽培种。
近年来人们对ICE基因及其低温调控机制比较关注,研究重点主要集中在拟南芥等模式植物中,且对除低温胁迫外的其他非生物胁迫的研究也相对较少。本研究从胡萝卜品种‘黑田五寸’及‘君川红’中分别克隆了DcICE1基因,并对DcICE1基因进行了较为详细的生物信息学分析,利用荧光定量PCR检测其在38 ℃高温、4 ℃低温、0.2 mol·L-1NaCl、200 g·L-1PEG等胁迫处理不同时间段的表达情况,以了解胡萝卜抗逆转录调控的作用机制,为通过遗传方法提高作物抗逆性奠定基础。
1材料和方法
1.1植物材料
植物材料胡萝卜品种 ‘黑田五寸’和‘君川红’于2014年03月份种植于南京农业大学作物遗传与种质创新国家重点实验室人工气候室。待长至2月龄时,分别进行低温(4 ℃)、高温(38 ℃)、盐(0.2 mol·L-1NaCl)、干旱(200 g·L-1PEG)处理,在处理的0、1、2、4、8和12 h分别取叶片样品,按照RNA simple Total RNA Kit试剂盒(北京天根公司)说明书分别提取总RNA,并用PrimeScript RT reagent Kit(大连TaKaRa公司)将提取的总RNA反转录成cDNA,用于荧光定量PCR。
1.2胡萝卜DcICE1基因克隆
基于本实验室胡萝卜转录组数据,拼接得到胡萝卜DcICE1基因的序列,并设计1对引物,DcA13F(5′-CAAAATGCTCCCCACTTCCAACGGCG-3′)和DcA13R(5′-AGCTAAATTGCTCCATGAAAACCAG-C-3′)。分别以‘黑田五寸’和‘君川红’未经处理的叶片的cDNA为模板进行扩增,PCR反应条件为:94 ℃预变性 5 min;94 ℃变性 30 s,54 ℃退火 30 s,72 ℃延伸 1 min 45 s,共35个循环;72 ℃延伸10 min。反应产物经质量浓度12 g/L琼脂糖凝胶电泳分离,利用AxyPrep DNA凝胶回收试剂盒(美国Axygen公司)回收后,连接pMD19-T载体(大连TaKaRa公司)并转化到大肠杆菌DH5α(由本实验室保存),提取质粒经PCR鉴定后委托南京金斯瑞生物科技有限公司测序。
1.3胡萝卜DcICE1基因序列分析
各植物相关基因和氨基酸序列均来自NCBI数据库,多序列比对采用Clustal X软件,用MEGA 5构建分子进化树,生成报告图形[25],蛋白质基本性质分析使用http://www.expasy.org网站相关软件完成[26],利用DNAMAN软件进行蛋白质疏水性/亲水性分析。
1.4胡萝卜DcICE1转录因子基因实时定量PCR表达分析
荧光定量 PCR(Quantitative real time RT-PCR)采用SYBR PremixExTaq试剂盒(大连TaKaRa公司),在iQTM5 Real-time PCR System完成,采用iQTM5 software进行分析,按照操作说明进行。相对定量使用参照基因的ΔΔCT法,表达差异等于2-ΔΔCT,ΔCT=CT目标基因-CTactin。相对定量是基于处理和对照之间,目标基因相对内参基因的表达量的比较[27]。以胡萝卜Tubulin基因为参照基因,引物为DcA15F(5′-GAGTGGAGTTACCTGCTGCCTTC-3′)和DcA15R(5′-ATGTAGACGAGGGAACGGAATCAAG-3′)。用于DcICE1基因表达分析的引物为DcA14F(5′-GCTTCCATACTTGGCGATGCTAT-TG-3′)和DcA14R(5′-CTCTTGCCCTTACCTC-GACCCTTGC-3′)。
2结果与分析
2.1胡萝卜DcICE1基因克隆
以胡萝卜‘黑田五寸’和‘君川红’未经处理叶片的cDNA为模板,以DcA13F和DcA13R为引物进行扩增,分别得到长约1 500 bp的扩增产物(图1)。将回收产物分别进行连接、转化、序列测定,获得2个扩增产物序列。对胡萝卜‘黑田五寸’和‘君川红’扩增产物核苷酸序列推导的氨基酸序列进行BLAST检索与比对,发现其均有ICE1转录因子所具有的保守功能域,且具有ICE1蛋白具有的bHLH(碱性-螺旋-环-螺旋)DNA结合域,属于ICE1家族。序列测定与分析表明,‘黑田五寸’和‘君川红’中的DcICE1基因(GenBank登录号分别为KM487593和KM487594)均含有一个1 458 bp的开放阅读框,编码485个氨基酸,两者在核苷酸水平上有2个碱基的差异,分别为第139位的G/A,第475位的A/G;氨基酸水平上有2个位点的差异,分别为第47位的E/K,第159位的N/D。对2个品种的DcICE1转录因子进行氨基酸亲水性/疏水性分析,结果表明,胡萝卜DcICE1转录因子中大部分氨基酸属于亲水性氨基酸(图2)。
2.2胡萝卜DcICE1转录因子氨基酸序列分析及理化性质分析
通过氨基酸序列检索和比对得知,胡萝卜ICE1转录因子与大白菜(Brassicapekinensis)、大豆(Glycinemax)、拟南芥(Arabidopsisthaliana)等的相似性均较高,且均具有bHLH保守区域及ICE1转录因子具有的功能域(图3)。对上述植物的氨基酸组成成分及理化性质进行分析(表1),各植物的氨基酸残基数在420~550之间不等,其理论相对分子质量在45~61 kD之间,碱性氨基酸比例高于酸性氨基酸比例,个别植物差异较明显,脂肪族氨基酸、芳香族氨基酸的比例均有一定的差异,可能与其在非保守区域的氨基酸差异较大有关。

图1 胡萝卜DcICE1基因克隆图谱

图2 胡萝卜DcICE1氨基酸序列疏水性(A)
2.3胡萝卜DcICE1转录因子的进化分析
为了进一步分析胡萝卜‘黑田五寸’和‘君川红’中DcICE1转录因子与其他物种中相关蛋白的进化关系,选取了相似性较高的几种植物进行进化分析。如图4所示,胡萝卜与大豆中的ICE转录因子属于同一个分支,亲缘关系较近;十字花科的油菜、大白菜、萝卜、拟南芥同属于一个分支,而葡萄科的葡萄、蔷薇科的苹果、茄科的番茄属于不同的分支,说明同科的植物进化关系最近。

图3 胡萝卜DcICE1转录因子与其他物种氨基酸序列的多重比对
2.4胡萝卜DcICE1基因在不同逆境处理下表达情况
为了分析DcICE1基因在胡萝卜非生物胁迫中的响应情况,对胡萝卜‘黑田五寸’和‘君川红’2个品种进行了38 ℃高温、4 ℃低温、0.2 mol·L-1NaCl、200 g·L-1PEG不同时间段的处理。结果(图5)表明,胡萝卜DcICE1转录因子基因在4 ℃低温处理时表达,‘黑田五寸’品种在2 h时表达量达到最高水平,是对照的2.3倍,而‘君川红’品种在处理1 h后表达量稍增加,随着处理时间的延长,表达量均有所下降;38 ℃高温处理下,DcICE1基因在2个胡萝卜品种中的表达量均较对照低,其中处理2 h和4 h,‘君川红’和‘黑田五寸’的表达量又相对高于其他处理时间;0.2 mol·L-1NaCl处理时,2个品种DcICE1基因的表达量均下降;200 g·L-1PEG处理下,2个品种各个时间段的表达量同样与对照相比,均有所下降,其中‘黑田五寸’在处理的各时间段表达量呈连续下降趋势,而‘君川红’各处理时间变化更加平缓。DcICE1转录因子基因在2个胡萝卜品种中响应逆境的时间和强度不同,可能与2个品种本身抗逆性差异有关。

图4 部分物种ICE转录因子氨基酸序列的系统进化树
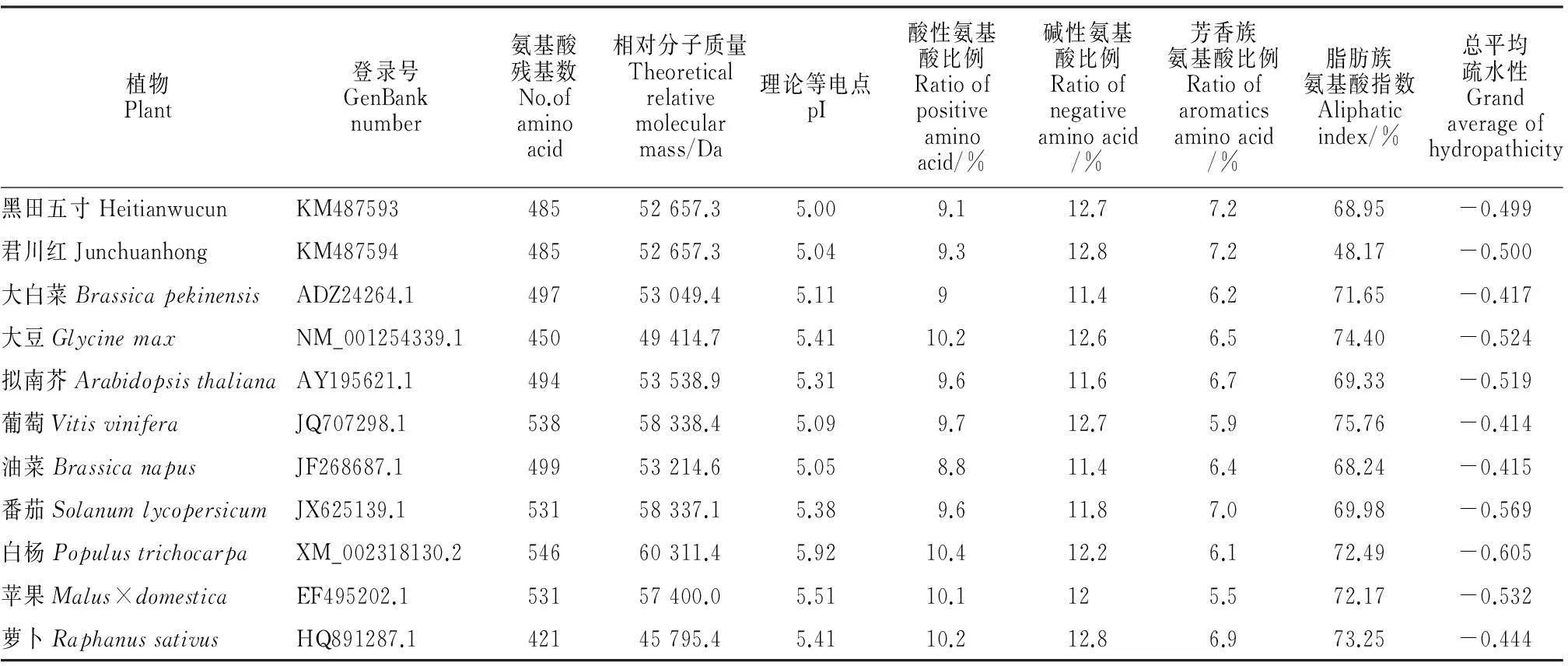
植物Plant登录号GenBanknumber氨基酸残基数No.ofaminoacid相对分子质量Theoreticalrelativemolecularmass/Da理论等电点pI酸性氨基酸比例Ratioofpositiveaminoacid/%碱性氨基酸比例Ratioofnegativeaminoacid/%芳香族氨基酸比例Ratioofaromaticsaminoacid/%脂肪族氨基酸指数Aliphaticindex/%总平均疏水性Grandaverageofhydropathicity黑田五寸HeitianwucunKM48759348552657.35.009.112.77.268.95-0.499君川红JunchuanhongKM48759448552657.35.049.312.87.248.17-0.500大白菜BrassicapekinensisADZ24264.149753049.45.11911.46.271.65-0.417大豆GlycinemaxNM_001254339.145049414.75.4110.212.66.574.40-0.524拟南芥ArabidopsisthalianaAY195621.149453538.95.319.611.66.769.33-0.519葡萄VitisviniferaJQ707298.153858338.45.099.712.75.975.76-0.414油菜BrassicanapusJF268687.149953214.65.058.811.46.468.24-0.415番茄SolanumlycopersicumJX625139.153158337.15.389.611.87.069.98-0.569白杨PopulustrichocarpaXM_002318130.254660311.45.9210.412.26.172.49-0.605苹果Malus×domesticaEF495202.153157400.05.5110.1125.572.17-0.532萝卜RaphanussativusHQ891287.142145795.45.4110.212.86.973.25-0.444

图5 胡萝卜DcICE1基因在不同逆境处理下不同时间的表达情况
3讨论
在植物生长发育过程中,会不断受到各种非生物胁迫而严重影响作物的产量和品质[28-29],低温就是一类重要的非生物影响因子。进一步深入了解植物抗寒机制对于农业生产有着重要的意义。胡萝卜生产中,低温会影响胡萝卜的生长发育,也是制约胡萝卜栽培区域的重要因素之一。
本实验分别从‘黑田五寸’和‘君川红’2个品种中克隆并分析了胡萝卜DcICE1基因,发现有2个核苷酸位点差异,编码氨基酸有2个位点差异。氨基酸序列及同源进化树分析显示,2个胡萝卜品种中的DcICE1转录因子在氨基酸性质如氨基酸数目、酸碱氨基酸比例、等电点、疏水性/亲水性等方面高度一致,但不同植物氨基酸理化性质有一定的差异,这可能与不同物种进化过程中的亲缘关系有关[30]。伞形科胡萝卜的DcICE1转录因子与蝶形花科大豆的ICE转录因子同属一个分支,亲缘关系较为密切。十字花科的大白菜、油菜、拟南芥及萝卜的ICE转录因子属于另外一个分支,而葡萄科的葡萄、蔷薇科的苹果、茄科的番茄分属于不同的分支,这说明不同物种ICE转录因子的进化关系呈现明显的种属特性。
有研究表明,ICE1基因是CBF转录因子基因的正调控因子,在植物抵抗寒冷环境中发挥着重要的作用[8,15]。ICE1基因是一种诱发基因,即在正常条件下不启动基因表达,当低温胁迫时诱导CBF基因表达,进而启动一系列相关基因表达,增强植物的抗寒性,在植物的生长发育中起着重要的作用[31]。本研究通过荧光定量PCR检测胡萝卜DcICE1基因在不同逆境不同时间段处理的表达情况,结果表明高温及盐胁迫不会引起DcICE1基因过量表达,而4 ℃低温处理下2 h后胡萝卜‘黑田五寸’DcICE1转录因子基因的表达量明显增加,‘君川红’1 h后表达量增加,这可能与不同胡萝卜品种特性有关。
另外,高等植物的抵御非生物胁迫机制复杂,涉及到很多的生理生化过程,参与逆境调控的基因非常多[32]。本研究使用生产上常用的2个胡萝卜品种,抗热、抗病性能较强的‘黑田五寸’和抗寒、抗热及抗病性强的‘君川红’,2品种DcICE1转录因子基因在不同逆境处理下响应的时间和强度表现出较大的差异,这可能与2品种中DcICE1转录因子个别氨基酸位点差异有关,也可能与品种本身特性有关。另外,有研究表明,葡萄的VaICE1基因能被盐诱导,为正调控,与本实验结果有些差异,这可能与物种不同有关,也可能与植物生长的环境差异有关[33-34]。
参考文献:
[1]ZHONG K Y(钟克亚),YE M SH(叶妙水),HU X W(胡新文),etal.Role of the transcription factors CBF in plant cold tolerance[J].Hereditas(遗传),2006,28(2):249-251(in Chinese).
[2]THOMASHOW M F.Plant cold acclimation:freezing tolerance genes and regulatory mechanisms[J].AnnualReviewofPlantBiology,1999,50(1):571-599.
[3]THOMASHOW M F.Arabidopsisthalianaas a model for studying mechanisms of plant cold tolerance[J].ColdSpringHarborMonographArchive,1994,27:807-834.
[4]VISWANATHAN C,ZHU J K.Molecular genetic analysis of cold-regulated gene transcription[J].PhilosophicalTransactionsoftheRoyalSocietyofLondon.SeriesB:BiologicalSciences,2002,357(1 423):877-886.
[5]ZHANG T G(张腾国),CHANG Y(常燕),WANG J(王娟),etal.Cloning and expression analysis of aBnICE1 fromBrassicanapusL.[J].ScientiaAgriculturaSinica(中国农业科学),2013,46(1):205-214(in Chinese).
[6]GUY C L,NIEMI K J,BRAMBL R.Altered gene expression during cold acclimation of spinach[J].ProceedingsoftheNationalAcademyofSciencesUSA,1985,82(11):3 673-3 677.
[7]XIANG D J(向殿军),YIN K D(殷奎德),MAN L L(满丽莉),etal.Cloning and genetic transformation of low temperature transcription factor fromRaphanussativus[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2011,31(10):1 933-1 941(in Chinese).
[8]CHINNUSAMY V,OHTA M,KANRAR S,etal.ICE1:a regulator of cold-induced transcriptome and freezing tolerance inArabidopsis[J].Genes&Development,2003,17(8):1 043-1 054.
[9]ZARKA D G,VOGEL J T,COOK D,etal.Cold induction ofArabidopsisCBFgenes involves multiple ICE(inducer ofCBFexpression) promoter elements and a cold-regulatory circuit that is desensitized by low temperature[J].PlantPhysiology,2003,133(2):910-918.
[10]LIN Y ZH(林元震),GUO H(郭海),LIU CH X(刘纯鑫),etal.Molecular cloning and expression analysis of cold-resistant transcription factor ICE1 fromEucalyptuscamaldulensisL.[J].PlantPhysiologyJournal(植物生理学报),2011,47(5):488-494(in Chinese).
[11]BADAWI M,REDDY Y V,AGHARBAOUI Z,etal.Structure and functional analysis of wheatICE(inducer ofCBFexpression) genes[J].PlantandCellPhysiology,2008,49(8):1 237-1 249.
[12]YIN Y(尹盈),ZHANG Y(张玥),HU J Y(胡靖妍),etal.Subcellular localization and expression analysis of CsICE transcription factor related to cold stress in tea plant[J].ActaHorticulturaeSinica(园艺学报),2013,40(10):1 961-1 968(in Chinese).
[13]FENG X M,ZHAO Q,ZHAO L L,etal.The cold-induced basic helix-loop-helix transcription factor geneMdCIbHLH1 encodes an ICE-like protein in apple[J].BMCPlantBiology,2012,12(1):22.
[14]PENGP H,LIN C H,TSAI H W,etal.Cold response inPhalaenopsisaphroditeand characterization ofPaCBF1 andPaICE1[J].PlantandCellPhysiology,2014,55(9):1 623-1 635.
[15]CHEN Y,JIANG J,SONG A,etal.Ambient temperature enhanced freezing tolerance ofChrysanthemumdichrumCdICE1Arabidopsisvia miR398[J].BMCBiology,2013,11(1):121.
[16]LEE B,HENDERSON D A,ZHU J K.TheArabidopsiscold-responsive transcriptome and its regulation byICE1[J].PlantCell,2005,17(11):3 155-3 175.
[17]LIU L,DUAN L,ZHANG J,etal.Cucumber(CucumissativusL.) over-expressing cold-induced transcriptome regulator ICE1 exhibits changed morphological characters and enhances chilling tolerance[J].ScientiaHorticulturae,2010,124(1):29-33.
[18]HUANG J Q,SUN Z H.Agrobacterium-mediated transfer ofArabidopsisICE1 gene into lemon(Citruslimon(L.) Burn.F.cv.Eureka)[J].AgriculturalSciencesinChina,2005,4(9):714-720.
[19]WANG B Y(王北艳),YIN K D(殷奎德).Expression ofrd29A-ICE1 gene in transgenic rice plants show increased tolerance to freezing stress[J].JournalofNuclearAgriculturalScience(核农学报),2013,27(6):731-735(in Chinese).
[20]FENG H L,MA N N,MENG X,etal.A novel tomato MYC-type ICE1-like transcription factor,SlICE1a,confers cold,osmotic and salt tolerance in transgenic tobacco[J].PlantPhysiologyandBiochemistry,2013,73:309-320.
[21]ZHANG N(张宁),SI H J(司怀军),WANG D(王蒂).Cloning ofrd29Agene promoter fromArabidopsisthalianaand its application in stress-resistance transgenic potato[J].ActaAgronomicaSinica(作物学报),2005,31(2):159-164(in Chinese).
[22]RUBATZKY V E,QUIROS C F,SIMON P W.Carrots and related vegetable umbelliferae[M].New York:CABI Publish-ing,1999.
[23]MAO J H(毛笈华),MAO SH M(茅淑敏),ZHUANG F Y(庄飞云),etal.Heredity and environmental regulation of premature bolting in carrot[J].ActaAgriculturaeBoreali-Sinica(华北农学报),2013,28(3):67-72(in Chinese).
[24]章镇.园艺学各论[M].北京:中国农业出版社,2004.
[25]TAMURA K,PETERSON D,PETERSON N,etal.MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods[J].MolecularBiologyandEvolution,2011,28(10):2 731-2 739.
[26]GASTEIGER E,GATTIKER A,HOOGLAND C,etal.ExPASy:the proteomics server for in-depth protein knowledge and analysis[J].NucleicAcidsResearch,2003,31(13):3 784-3 788.
[27]TAN G F(谭国飞),WANG F(王枫),etal.Isolation and expression analysisof cinnamic acid 4-hydroxylase gene under different temperatures inCryptotaeniajaponicaHassk[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2014,34(7):1 298-1 304(in Chinese).
[28]WANG Y Q(王雅琴),SHI M(石淼),ZHANG X Y(张新宇),etal.Cloning and expression analysis ofGhMBY113 gene ingossypium hirsutum[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2013,33(5):878-884(in Chinese).
[29]PEI L L(裴丽丽),GUO Y H(郭玉华),XU ZH SH(徐兆师),etal.Research progress on stress-related protein kinases in plants[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2012,32(5):1 052-1 061(in Chinese).
[30]SOLTIS P S,SOLTIS D E.The origin and diversification of angiosperms[J].AmericanJournalofBotany,2004,91(10):1 614-1 626.
[31]GILMOURS J,ZARKA D G,STOCKINGER E J,etal.Low temperature regulation of theArabidopsisCBF family of AP2 transcriptional activators as an early step in cold-inducedCORgene expression[J].PlantJournal,1998,16(4):433-442.
[32]ZHUANG J,ZHANG J,HOU X L,etal.Transcriptomic,proteomic,metabolomic and functional genomic approaches for the study of abiotic stress in vegetable crops[J].CriticalReviewsinPlantSciences,2014,33(2-3):225-237.
[33]ZHOU M Q,SHEN C,WU L H,etal.CBF-dependent signaling pathway:a key responder to low temperature stress in plants[J].CriticalReviewsinBiotechnology,2011,31(2):186-192.
[34]HUANG G T,MA S L,BAI L P,etal.Signal transduction during cold,salt,and drought stresses in plants[J].MolecularBiologyReports,2012,39(2):969-987.
(编辑:宋亚珍)
Cloning and Expression Profiles Analysis under Abiotic Stresses of Cold
Stress Transcription Factor GeneDcICE1 fromDaucuscarota
HUANG Ying,XU Zhisheng,WANG Feng,LI Mengyao,MA Jing,XIONG Aisheng*
(State Key Laboratory of Crop Genetics and Germplasm Enhancement,Ministry of Agriculture Key Laboratory of Biology and Germplasm Enhancement of Horticultural Crops in East China,College of Horticulture,Nanjing Agricultural University,Nanjing 210095,China)
Abstract:In higher plant,ICE1 involved in the response to abiotic and biotic stresses by regulating the expression ofCBFgenes.The cDNA sequence of DcICE1 transcription factor was cloned from carrot cultivars ‘Heitianwucun’ and ‘Junchuanhong’ using RT-PCR method,respectively.In order to analyze the response ofDcICE1 gene to abiotic stress,the expression profiles ofDcICE1 gene in four different stresses(4 ℃,38 ℃,0.2 mol·L-1NaCl,200 g·L-1PEG) were detected by quantitative real-time PCR.Sequence analysis indicated that theDcICE1 gene contained 1 458 bp,encoding 485 amino acids.There were two nucleotide sites and two amino acid residue differences between two cultivars(G/A in site 139 and A/G in site 475;E/K in site 47,N/D in 159).The different amino acid residues may be involved into the different response in two carrot cultivars encountered the abiotic stresses.The DcICE1 transcription factor has a conserved domain.Quantitative real-time PCR detected the expression profiles ofDcICE1 gene under abiotic stresses.The results showed the expression ofDcICE1 gene significantly increased when exposed to 4 ℃,while the expression profiles ofDcICE1 gene was not obvious after treated by salt and PEG in the two carrot cultivars.
Key words:Daucuscarota;DcICE1 transcription factor;gene cloning;abiotic stress;expression analysis
中图分类号:Q789
文献标志码:A
作者简介:黄莹(1990-),女,在读硕士研究生,主要从事蔬菜分子生物学研究。E-mail:2013104056@njau.edu.cn*通信作者:熊爱生,教授,博士生导师,主要从事蔬菜分子生物学和遗传育种研究。E-mail:xiongaisheng@njau.edu.cn
基金项目:教育部新世纪优秀人才支持计划(NCET-11-0670);江苏省杰出青年基金(BK20130027);中国博士后基金(2014M551609);江苏高校优势学科建设项目(2011PAPD);江苏省双创计划(2011JSSC)
收稿日期:2014-07-08;修改稿收到日期:2014-10-11
文章编号:1000-4025(2015)01-0030-07
doi:10.7606/j.issn.1000-4025.2015.01.0030
