Nesfatin-1在抑郁症发病中的作用研究进展
2015-02-26徐亚运葛金芳陈飞虎
徐亚运,葛金芳,陈飞虎
(安徽医科大学药学院,安徽合肥 230032)
Nesfatin-1在抑郁症发病中的作用研究进展
徐亚运,葛金芳,陈飞虎
(安徽医科大学药学院,安徽合肥 230032)
摘要:摄食抑制因子Nesfatin-1是由核组蛋白NUCB2的N端水解产生的由82个氨基酸组成的肽段。自2006年日本Oh-I教授首次报道Nesfatin-1广泛表达于下丘脑以来,关于其生理功能的研究不断深入。近期研究表明,除了参与摄食及能量代谢的调节之外,Nesfatin-1还参与情绪反应的中枢调控,与抑郁症的发生密切相关。该文就Nesfatin-1的分布及其在抑郁症发病机制的作用做一综述,旨在明确Nesfatin-1的生理功能,也为进一步揭示抑郁症的发病机制提供依据。
关键词:Nesfatin-1;摄食行为调节;应激反应;下丘脑-垂体-肾上腺轴;抑郁;快感缺失;胰岛素信号通路
网络出版时间:2015-7-22 10:42 网络出版地址:http://www.cnki.net/kcms/detail/34.1086.R.20150727.0901.006.html
Nesfatin-1是新近发现的摄食抑制因子,是由nucleo-bindin2(NUCB2)N端水解产生的由82个氨基酸组成的肽段,2006年日本教授Oh-I最先报道其广泛表达于下丘脑[1]。众所周知,下丘脑是调节摄食、能量代谢和情绪反应的主要中枢结构之一,关于Nesfatin-1生物学活性的早期研究也多聚焦于摄食及能量代谢的调节。近期研究表明,Nesfatin-1可参与情绪反应的中枢调控,与抑郁症的发病密切相关。本文主要就Nesfatin-1在抑郁症发病中的作用综述如下。
1 Nesfatin-1的发现和体内分布
Oh-I等[1]利用PPAR-γ激动剂曲格列酮作用于Q-5细胞株,发现9种基因可以被激活,其中编码NEFA(DNA-bind-ing/EF-hand/acidic protein)/核组蛋白2(nucleobindin-2,NU-CB2)5’端非翻译区的基因序列受其调节最为明显。随后将NUCB2蛋白注入大鼠侧脑室,发现可以剂量和时间依赖性减少大鼠摄食量,遂将其命名为核组蛋白NUCB2。在转化酶的作用下,NUCB2可以裂解为3个片段,将其N段82个氨基酸片段命名为Nesfatin-1。Nesfatin-1又可以分为3个片段,其中中间片段M30是Nesfatin-1发挥厌食作用的核心部位。
下丘脑室旁核(paraventricular nucleus,PVN)、弓状核(arcuate nucleus,ARC)、室上核(supraoptic nucleus,SON)和下丘脑外侧区(lateral hypothalamic area,LHA)被首先证明是Nesfatin-1产生的主要部位[1],而且Nesfatin-1在PVN和SON神经元中与抗利尿激素、促肾上腺皮质激素释放激素(corticotropin releasing hormone,CRH)以及催产素共表达,在ARC神经元中与可卡因-苯丙胺调节转录肽(cocaine-and amphetamine-regulated transcript,CART)及酪氨酸羟化酶共表达,在LHA神经元与CART及黑素细胞凝集素(melanin-concentrating hormone,MCH)共表达[2]。随后的研究表明[2-3],Nesfatin-1还广泛存在于中脑等其他脑区,包括丘脑背内侧、迷走神经运动背核(dorsal vagal complex,DVC)、动眼神经副核(Edinger-Westphal nucleus,E-W核)等。随后,Stengel等[3]通过免疫组化发现大鼠的岛叶皮质、中央杏仁核、延髓腹外侧、小脑浦肯野细胞、脑的副交感神经和交感神经节前神经元以及腰椎和骶骨的脊髓中亦广泛表达Nesfa-tin-1。并且在脑干的E-W核和DVC核Nesfatin-1与乙酰胆碱转移酶共表达,在脑干的孤束核(nucleus of the solitary tract,NTS)与酪氨酸羟化酶共表达,在中缝核则与5-羟色胺(5-hydroxytryptamine,5-HT)共表达[2]。
除了广泛表达于中枢神经系统之外,Nesfatin-1还被证明在胃肠道、脂肪组织、胰腺、睾丸和心脏等外周组织中表达,并且Nesfatin-1可以通过血脑屏障[4],提示了Nesfatin-1具有广泛的生理作用。
2 Nesfatin-1与抑郁症
抑郁症是一种常见的情感障碍性疾病,主要表现为情绪低落、摄食行为改变、活动减少以及高度的自杀倾向。世界卫生组织的全球疾病负担(global burden of disease,GBD)合作研究预测在2020年抑郁症将成为全球首位疾病负担。抑郁症发病机制的“单胺神经递质学说”认为包括5-HT在内的单胺类神经递质的耗竭是导致抑郁症发生的主要原因[5]。但以提高突触间隙的单胺类神经递质为目标的治疗措施并非对所有的抑郁症患者有效,而且存在治疗潜伏期长、副作用大等缺陷[6]。因而,深入探索抑郁症发病的机制成为神经科学重点关注的领域之一。
现代神经生物学研究结果表明,伴随下丘脑-垂体-肾上腺轴(hypothalamic-pituitary-adrenal axis,HPA轴)过度激活的下丘脑多受体平衡紊乱是导致抑郁症发病的主要原因之一[7],参与摄食调节的下丘脑相关信号分子可直接或通过调节HPA轴的活性参与抑郁症的发病机制。研究表明,促黑素(melanocyte stimulating hormone,MSH)等摄食抑制因子与焦虑抑郁行为的产生密切相关,而促食素(orexin)等促进摄食因子则可改善焦虑抑郁相关行为。鉴于摄食抑制因子Nesfatin-1的主要生理学功能是参与摄食和代谢调控,近年来针对其与抑郁症相互关系的研究逐渐增多。
2.1Nesfatin-1与摄食行为调节 摄食行为改变及其引发的体重改变及代谢紊乱是抑郁症的主要临床表现之一。研究发现[1],侧脑室注射Nesfatin-1可剂量依赖性抑制大鼠对食物的摄取并导致体重减轻,且这一效应可被Nesfatin-1的抗体阻断。Stengel等[8]的研究也证实,第4脑室以及脑池内注射Nesfatin-1可在较长时间内持续抑制大鼠夜间摄食行为。Shimizu等[9]的研究结果则表明,长期外周注射(腹腔注射或皮下注射)Nesfatin-1也可以剂量依赖性抑制自由饮食的ICR小鼠夜间摄食并减少体重增长。这些研究结果表明,无论是中枢还是外周注射Nesfatin-1都可抑制摄食行为,而且中枢效应远远强于外周[10]。
研究表明,Nesfatin-1可以在瘦素受体突变的Zurker大鼠中[1]以及瘦素抵抗的高脂肪饮食导致肥胖的大鼠[9]中发挥厌食效应,提示Nesfatin-1的摄食抑制作用可能不依赖瘦素信号途径。进一步研究发现,Nesfatin-1的抑制摄食作用与应激反应以及情绪调控因子存在关联。Oh-I等[1]通过中枢注射a-MSH发现PVN中NUCB2基因表达增加,并且该过程能够被黑皮质素3/4受体特异性拮抗剂SHU9119抑制,提示Nesfatin-1可能通过中枢黑皮质素系统发挥其抑制摄食的作用。形态学研究结果证实,Nesfatin-1在下丘脑与MCH共定位表达,80%Nesfatin-1神经元分泌MCH,提示Nesfatin-1的抑制摄食效应可能通过抑制MCH的促进摄食行为实现[11]。然而,黑皮质素系统介导的厌食效应可以被CRH受体拮抗剂所阻断,而且CRH的受体拮抗剂astressin-B2可以解除侧脑室注射Nesfatin-1引起的饱腹感。因而,CRH可能作为中枢黑皮质素系统的下游因子来介导Nesfatin-1的抑制摄食行为[8]。
新近研究表明[12],Nesfatin-1可以增加下丘脑中CRH和促甲状腺激素释放激素(thyrotropin-releasing hormone,TRH)的水平,Nesfatin-1的厌食效应在使用CRH受体拮抗剂、TRH抗体以及敲除组胺受体的小鼠明显减弱,而且在结节乳头核(tuberomammillary nucleus,TMN)组胺神经元细胞膜上存在CRH受体1和TRH受体2,提示Nesfatin-1对摄食行为的抑制作用可能与其对CRH神经元、TRH神经元及TMN组胺神经元的调控有关。
2.2Nesfatin-1与抑郁相关行为
2.2.1应激刺激与Nesfatin-1表达 动物实验及临床研究[13]证实,应激性生活事件及其引发的HPA轴过度激活是抑郁症发生的主要原因之一。研究表明,Nesfatin-1在中枢和外周均不同程度地参与了应激反应。束缚应激可导致大鼠SON、PVN、蓝斑核(locus coeruleus,LC)、延髓中缝核(ros-tral raphe pallidus,rRPa)、NTS、延髓腹外侧(ventrolateral me-dulla,VLM)中Nesfatin-1阳性神经元c-FOS表达上调,提示Nesfatin-1可能是应激反应中的响应因子[14]。急性束缚应激还能够促进E-W核神经元共表达的尿皮质素、CART、Nes-fatin-1明显升高。Konczol等[14]的研究结果则证实,Nesfatin-1在延髓可通过NA和催产素等信号传递分子与应激反应中心PVN产生联系。其他几种急性应激反应如强迫游泳[3]、腹部手术、腹腔注射脂多糖也能够增加Nesfatin-1在中枢系统的表达(Tab 1)。这些研究结果表明,Nesfatin-1参与应激反应的中枢调节。
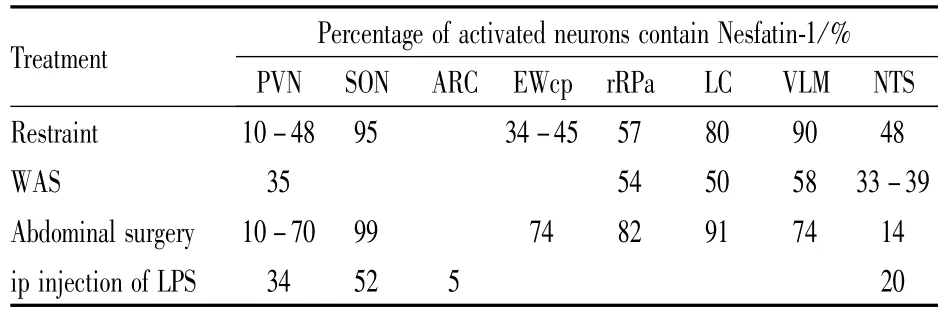
Tab 1 Activation of Nesfatin-1 containing neurons in brain induced by different stressors
慢性温和不可预知应激模型(chronic unpredictable mild stress,CUMS)能够较好地模拟人类抑郁症发病的病理生理机制,再现抑郁相关行为,是研究抑郁症发病机制和治疗药物筛选的主要动物模型之一。Xu等[15]采用冷水游泳、潮湿鼠笼、禁食禁水等慢性温和应激刺激构建了大鼠CUMS模型,并且发现中脑E-W核中Nesfatin-1和CART神经元活动性增强,提示可能通过CART参与了慢性应激反应过程。
Albayrak等[16]的研究表明,烧伤感染的应激患者血浆中Nesfatin-1水平较对照组明显降低,但随烧伤治疗后逐渐回升。Yoshida等[17]的研究则表明,急性束缚应激可导致下丘脑PVN、SON及中缝核、蓝斑核等脑区Nesfatin-1表达上调,但血浆Nesfatin-1水平则无明显改变。新近研究发现[18]腹腔注射Nesfatin-1可减少强迫游泳和束缚应激导致的胃酸分泌以及胃黏膜损伤。提示Nesfatin-1在外周也参与应激反应的病理生理过程。
以上研究结果表明,急性和慢性应激反应均可导致Nes-fatin-1的表达改变,而且Nesfatin-1参与应激反应的中枢和外周机制可能存在差异。
2.2.2Nesfatin-1干预与抑郁相关行为 行为学实验研究证实,Nesfatin-1可诱导焦虑抑郁样行为改变。Merali等[19]观察了脑室内注射Nesfatin-1后大鼠在高架十字迷宫和新环境摄食抑制实验中的表现,结果表明,中枢注射Nesfatin-1后大鼠在开放臂的逗留时间明显减少,在新环境中对食物探索的潜伏期明显升高,对食物的摄取量也明显减少。提示中枢注射Nesfatin-1可诱导大鼠焦虑抑郁样行为改变。
研究人员通过给大鼠侧脑室注射Nesfatin-1发现可以激活CRH、去甲肾上腺素(noradrenaline,NA)和5-HT神经元,并且血浆ACTH和糖皮质激素的水平明显升高,双侧肾上腺切除后PVN中NUCB2表达升高[17]。结果显示Nesfatin-1激活了HPA轴,而HPA轴亢进作为目前抑郁症发病机制中公认的机制之一,提示Nesfatin-1可能通过激活HPA轴来诱导抑郁行为的产生。
2.3Nesfatin-1与抑郁症的临床研究 临床研究进一步探讨了Nesfatin-1与抑郁症之间的联系。Ari等[20]的研究发现,与健康对照组比较,重症抑郁症患者血浆Nesfatin-1升高至接近2倍水平,而且血浆Nesfatin-1水平与患者在汉密顿抑郁量表(hamilton depression scale,HAMD)中的得分呈正相关。提示Nesfatin-1与抑郁存在联系。
采用患者健康问卷(patient health questionnaire,PHQ-9)、广泛性焦虑量表(generalized anxiety disorder question-naire,GAD-7)和感知应激量表(perceived stress question-naire,PSQ-20),Hofmann等[21]的研究表明,在焦虑和抑郁评价中得分高的患者其血浆NUCB2/Nesfatin水平明显升高,血浆NUCB2/Nesfatin水平与PHQ-9、GAD-7和PSQ-20量表中的得分呈正相关,而与体质指数(body mass index,BMI)无关。进一步提示Nesfatin-1参与情绪反应的调节,可能参与抑郁症的发病。
由于实验对象的限制,并没有大量资料对人类中枢的Nesfatin-1改变进行研究,所以这种现象的机制还不是很清楚。然而,最近一项研究发现[22],抑郁症自杀患者的中枢Nesfatin-1水平发生了明显改变,并表现出性别差异。与对照组比较,抑郁自杀的男性患者E-W核Nesfatin-1升高了2倍,而女性患者则降低了3倍。与动物实验结果相一致的是,抑郁症患者E-W核中Nesfatin-1与CART存在共表达,进一步证实Nesfatin-1可能参与了抑郁症的发病,且其机制可能与CART相关。
2.4Nesfatin-1参与抑郁症发病的可能机制 无论是Nes-fatin-1注射产生抑郁相关行为,还是抑郁症患者体内Nesfa-tin-1含量的升高都显示出Nesfatin-1与抑郁症发病的密切联系,针对其可能涉及的神经生物学机制的研究也逐渐深入。
CRH、CART、Urocortin-1、NPY、oxytocin等神经肽的表达改变被证明是参与应激反应,并导致摄食行为改变及抑郁症发生的可能机制之一。大量研究证实[2,15,23],应激刺激后Nesfatin-1与上述神经肽存在共定位表达(Tab 2),这为Nes-fatin-1参与抑郁症发病的机制研究提供了思路。

Tab 2 Co-localization of Nesfatin-1 with stress sensitive substances in the brain
2.4.1Nesfatin-1通过调节HPA轴参与抑郁症发病 如前所述,侧脑室注射Nesfatin-1可导致HPA轴过度激活[17],诱发大鼠焦虑恐惧行为[19],形态学研究结果也证实Nesfatin-1 与CRH能神经元等共表达,CRH还被证明是Nesfatin-1的下游信号分子[17]。因而,调节HPA轴的活性可能是Nesfatin-1参与抑郁症的主要机制之一。
2.4.2Nesfatin-1通过中枢奖赏通路参与抑郁症发病 中枢奖赏通路,尤其是腹侧被盖区(ventral tegmental area,VTA)投射至伏隔核(nucleus accumbens,NAc)的多巴胺能神经元功能障碍是导致抑郁症患者快感缺失的主要神经生物学机制之一[24]。下丘脑摄食相关信号因子瘦素、胃饥饿素等可以通过调控中枢奖赏通路多巴胺能神经元的活性,影响摄食及快感缺失行为[25],多巴胺受体激动剂也可调节下丘脑摄食相关信号分子的表达[26]。新近研究表明[24],下丘脑弓状核的POMC能神经元可直接投射至伏隔核,α-MSH信号通过Melanocortin 4受体(MCR4)参与奖赏通路多巴胺能神经元活性的调控,更进一步地证实了下丘脑摄食调节因子与中枢奖赏通路之间的紧密联系。
研究表明,Nesfatin-1引起的摄食行为障碍可通过激活黑皮质素系统介导[1],侧脑室注射α-MSH可导致下丘脑Nesfatin-1表达上调,形态学研究结果也证实Nesfatin-1与POMC/CART能神经元的共表达。这些研究结果表明Nesfa-tin-1与黑皮质素系统之间存在密切的内在联系。另外,在ARC中,Nesfatin-1可以通过超极化来抑制NPY神经元,而促进摄食因子NPY被证明可以改善焦虑抑郁相关行为[27]。2.4.3 Nesfatin-1通过调节下丘脑胰岛素信号通路参与抑郁症发病 动物实验和临床研究结果表明,下丘脑摄食与代谢调节的稳态失衡,尤其是糖代谢调节紊乱与抑郁相关行为密切相关。胰岛素信号通路功能障碍导致的胰岛素抵抗被认为是导致抑郁症患者糖代谢紊乱及神经功能障碍的主要机制之一。研究表明,胰岛素信号通路参与突触结构及功能可塑性的调节,下调下丘脑胰岛素受体的表达可诱导大鼠抑郁相关行为的发生,慢性应激可导致大鼠下丘脑弓状核的胰岛素信号通路功能障碍,并与抑郁相关行为和HPA轴过度激活密切相关[28]。而且,与抑郁症密切相关的5-HT等单胺类神经递质也参与胰岛素分泌及糖代谢的调控,胰岛素可剂量依赖性降低中脑腹外侧被盖区突触内多巴胺浓度,降低享乐感觉,胰岛素抵抗可改变多巴胺受体及5-HT受体的敏感性,氟西汀等选择性5-HT再摄取抑制剂可抑制胰岛素分泌并改善抑郁大鼠的抑郁相关行为和HPA轴过度激活[28]。
近期研究表明,Nesfatin-1可与胰岛素共定位表达于胰岛β细胞[29],下丘脑靶向性Nesfatin-1敲除可调控胰岛素受体与胰岛素信号通路关键分子的表达,应用胰岛素信号通路关键分子的拮抗剂可以阻断Nesfatin-1对血糖的调控效应,提示Nesfatin-1的作用机制与胰岛素信号通路密切相关。另有研究表明,Nesfatin-1血浆浓度与糖尿病患者的胰岛素抵抗状态及血糖水平呈正相关[30],而胰岛素抵抗与高血糖都是导致快感缺失等抑郁表现的重要因素。因此,有理由相信,应激导致的下丘脑Nesfatin-1水平增高可能通过影响下丘脑胰岛素信号通路的活性,诱发并加重胰岛素抵抗,继而参与抑郁症的发生。
3 结语与展望
综上所述,Nesfatin-1的改变与抑郁症的摄食行为调节、快感缺失、行为绝望及糖代谢紊乱密切相关,Nesfatin-1可能参与抑郁症的发生。然而,由于Nesfatin-1的特异性受体至今仍未发现,Nesfatin-1作用的具体机制有待进一步明确。
参考文献:
[1] Oh-I S,Shimizu H,Satoh T,et al.Identification of nesfatin-1 as a satiety molecule in the hypothalamus[J].Nature,2006,443 (7112):709-12.
[2] Foo K S,Brismar H,Broberger C.Distribution and neuropeptide coexistence of nucleobindin-2 mRNA/nesfatin-like immunoreactiv-ity in the rat CNS[J].Neuroscience,2008,156(3):563-79.
[3] Goebel-Stengel M,Wang L,Stengel A,Tache Y.Localization of nesfatin-1 neurons in the mouse brain and functional implication [J].Brain Res,2011,1396:20-34.
[4] Pan W,Hsuchou H,Kastin A J.Nesfatin-1 crosses the blood-brain barrier without saturation[J].Peptides,2007,28(11):2223-8.
[5] 张亭亭,薛 瑞,李云峰,等.单胺转运蛋白与单胺重摄取抑制剂研究进展[J].中国药理学通报,2013,29(6):741-4.
[5] Zhang T T,Xue R,Li Y F,et al.Research progress in monoamine transporters and monoamine transmitter reuptake inhibitors[J].Chin Pharmacol Bull,2013,29(6):741-4.
[6] 刘佳莉,苑玉和,陈乃宏.抑郁症的治疗研究进展[J].中国药理学通报,2011,27(9):1193-6.
[6] Liu J L,Yuan Y H,Chen N H.Research progress in treatment of depression[J].Chin Pharmacol Bull,2011,27(9):1193-6.
[7] Ge J F,Peng L,Cheng J Q.et al.Antidepressanlike effect of res-veratrol:involvement of antioxidant effect and peripheral regulation on HPA axis[J].Pharmacol Biochem Behav,2013,114-115:64-9.
[8] Stengel A,Goebel M,Wang L,et al.Central nesfatin-1 reduces dark-phase food intake and gastric emptying in rats:differential role of corticotropin-releasing factor 2 receptor[J].Endocrinology,2009,150(11):4911-9.
[9] Shimizu H,Oh I S,Hashimoto K.et al.Peripheral administration of nesfatin-1 reduces food intake in mice:the leptin-independent mechanism[J].Endocrinology,2009,150(2):662-71.
[10]Stengel A,Tache Y.Minireview:nesfatin-1-an emerging new play-er in the brain-gut,endocrine,and metabolic axis[J].Endocri-nology,2011,152(11):4033-8.
[11]Fort P,Salvert D,Hanriot L,et al.The satiety molecule nesfatin-1 is co-expressed with melanin concentrating hormone in tuberal hypothalamic neurons of the rat[J].Neuroscience,2008,155(1):174-81.
[12]Gotoh K,Masaki T,Chiba S,et al.Nesfatin-1,corticotropin-re-leasing hormone,thyrotropin-releasing hormone,and neuronal his-tamine interact in the hypothalamus to regulate feeding behavior [J].J Neurochem,2013,124(1):90-9.
[13]Swaab D F,Bao A M,Lucassen P J.The stress system in the hu-man brain in depression and neurodegeneration[J].Ageing Res Rev,2005,4(2):141-94.
[14]Konczol K,Bodnar I,Zelena D,et al.Nesfatin-1/NUCB2 may participate in the activation of the hypothalamic-pituitary-adrenal axis in rats[J].Neurochem Int,2010,57(3):189-97.
[15]Xu L,Bloem B,Gaszner B,et al.Stress-related changes in the activity of cocaine-and amphetamine-regulated transcript and nes-fatin neurons in the midbrain non-preganglionic Edinger-Westphal nucleus in the rat[J].Neuroscience,2010,170(2):478-88.
[16]Albayrak A,Demiryilmaz I,Albayrak Y,et al.The role of dimin-ishing appetite and serum nesfatin-1 level in patients with burn wound infection[J].Iran Red Crescent Med J,2013,15(5):389 -92.
[17]Yoshida N,Maejima Y,Sedbazar U,et al.Stressor-responsive central nesfatin-1 activates corticotropin-releasing hormone,nora-drenaline and serotonin neurons and evokes hypothalamic-pituitary-adrenal axis[J].Aging(Albany NY),2010,2(11):775-84.
[18]Szlachcic A,Sliwowski Z,Krzysiek-Maczka G,et al.New satiety hormone nesfatin-1 protects gastric mucosa against stress-induced injury:mechanistic roles of prostaglandins,nitric oxide,sensory nerves and vanilloid receptors[J].Peptides,2013,49:9-20.
[19]Merali Z,Cayer C,Kent P,Anisman H.Nesfatin-1 increases anx-iety-and fear-related behaviors in the rat[J].Psychopharmacology (Berl),2008,201(1):115-23.
[20]Ari M,Ozturk O H,Bez Y,et al.High plasma nesfatin-1 level in patients with major depressive disorder[J].Prog Neuropsycho-pharmacol Biol Psychiatry,2011,35(2):497-500.
[21]Hofmann T,Stengel A,Ahnis A,et al.NUCB2/nesfatin-1 is as-sociated with elevated scores of anxiety in female obese patients [J].Psychoneuroendocrinology,2013,38(11):2502-10.
[22]Bloem B,Xu L,Morava E,et al.Sex-specific differences in the dynamics of cocaine-and amphetamine-regulated transcript and nesfatin-1 expressions in the midbrain of depressed suicide victims vs.controls[J].Neuropharmacology,2012,62(1):297-303.
[23]Brailoiu G C,Dun S L,Brailoiu E,et al.Nesfatin-1:distribution and interaction with a G protein-coupled receptor in the rat brain [J].Endocrinology,2007,148(10):5088-94.
[24]Lim B K,Huang K W,Grueter B A,et al.Anhedonia requires MC4R-mediated synaptic adaptations in nucleus accumbens[J].Nature,2012,487(7406):183-9.
[25]Chuang J C,Perello M,Sakata I,et al.Ghrelin mediates stress-induced food-reward behavior in mice[J].J Clin Invest,2011,121(7):2684-92.
[26]van Zessen R,van der Plasse G,Adan R A.Contribution of the mesolimbic dopamine system in mediating the effects of leptin and ghrelin on feeding[J].Proc Nutr Soc,2012,71(4):435-45.
[27]Bjornebekk A,Mathe A A,Brene S.The antidepressant effects of running and escitalopram are associated with levels of hippocampal NPY and Y1 receptor but not cell proliferation in a rat model of de-pression[J].Hippocampus,2010,20(7):820-8.
[28]Pan Y,Hong Y,Zhang Q Y,Kong L D.Impaired hypothalamic insulin signaling in CUMS rats:restored by icariin and fluoxetine through inhibiting CRF system[J].Psychoneuroendocrinology,2013,38(1):122-34.
[29]Gonzalez R,Tiwari A,Unniappan S.Pancreatic beta cells colocal-ize insulin and pronesfatin immunoreactivity in rodents[J].Bio-chem Biophys Res Commun,2009,381(4):643-8.
[30]Zhang Z,Li L,Yang M,et al.Increased plasma levels of nesfatin-1 in patients with newly diagnosed type 2 diabetes mellitus[J].Exp Clin Endocrinol Diabetes,2012,120(2):91-5.
Advances in studies on role of Nesfatin-1 in depression
XU Ya-yun,GE Jin-fang,CHEN Fei-hu
(School of Pharmacy,Anhui Medical University,Hefei 230032,China)
Abstract:Nesfatin-1,discovered in 2006 by Oh-I as an 82-ami-no-acid peptide derived from the precursor protein nucleobindin2 (NUCB2),has been identified to play an important role in the regulation of food intake and energy metabolism.Recently,it has also been found that Nesfatin-1 might be associated with the pathogenesis of depression.This article reviewed the advances in related studies on Nesfatin-1 at home and abroad,which should throw light in expliciting the physiological function of Nesfatin-1 and understanding the neurobiological mechanism of depression.
Key words:Nesfatin-1;feeding behavior;stress;hypothalamic-pituitary-adrenal axis;depression;anhedonia;insulin signaling
作者简介:徐亚运(1990-),男,硕士生,研究方向:分子药理学,E-mail:413271974@qq.com;葛金芳(1975-),女,副教授,研究方向:神经药理学,通讯作者,Tel:0551-65161115,E-mail:aygjf@mail.ustc.edu.cn;陈飞虎(1962-),男,博士,教授,博士生导师,研究方向:分子药理学、基因工程药物的表达与活性、中药药理学,通讯作者,Tel:0551-65161116,E-mail:cfhchina@sohu.com
基金项目:教育部高等学校博士点基金资助项目(No 20133420120005);安徽省自然科学基金资助项目(No 1408085MH154)
收稿日期:2015-06-10,修回日期:2015-07-08
文献标志码:A
文章编号:1001-1978(2015)08-1044-05中国图书分类号:R-05;R341.6;R749.41;R977.6
doi:10.3969/j.issn.1001-1978.2015.08.003