不同浓度红景天苷对成肌细胞体外分化的影响及机制初探
2015-02-14陈晓萍
罗 维,张 鹏,艾 磊,周 越,陈晓萍
不同浓度红景天苷对成肌细胞体外分化的影响及机制初探
罗 维1,张 鹏2,艾 磊3,周 越4,陈晓萍2
研究目的:探讨不同浓度红景天苷对成肌细胞株C2C12体外分化的影响及可能机制,有望为成肌细胞增殖分化寻求更有效的外源调控物,以促进成肌细胞的移植和临床应用,同时为进一步开展红景天苷在体内骨骼肌损伤中的作用及机制研究奠定基础。研究方法:研究1:红景天苷对成肌细胞体外分化的影响:1)在C2C12中加入不同浓度红景天苷处理,在诱导成肌分化过程中,应用相差显微镜观察不同浓度红景天苷对C2C12分化过程中汇聚、融合和形成多核肌管的影响;2)应用免疫荧光组化方法,分析不同浓度红景天苷对C2C12成肌分化融合率的影响;3)应用real-time PCR和Western Blotting方法,检测红景天苷对C2C12成肌分化特异基因和蛋白表达的影响;研究2:红景天苷影响成肌细胞体外分化的机制初探:应用Western Blotting方法,检测红景天苷处理后C2C12成肌分化中TGF-β/Smad信号通路的变化。结果:50 μg 红景天苷处理:研究1:1)光镜观察结果从形态学上显示,红景天苷处理显著抑制C2C12细胞的融合和多核肌管的形成;2)免疫荧光检测结果表明Myosin-neonatal(E-MHC)阳性面积和myogenin阳性核均显著减少(P<0.05),显著抑制C2C12细胞分化过程中肌管形成,促进细胞增殖;3)real-time PCR和Western blotting检测结果表明,成肌分化特异因子MyoD和myogenin基因和蛋白表达水平均显著下降(P<0.05),显著抑制C2C12细胞分化过程中细胞的融合和肌管的形成;研究2:Western blotting检测结果表明,红景天苷显著抑制C2C12细胞分化效应,伴有Smad2/Smad3磷酸化激活增加,尤其是Smad3激活水平显著增加(P<0.01);30 μg红景天苷处理作用不显著(P>0.05)。结论:1)红景天苷能显著抑制骨骼肌成肌细胞体外成肌分化,并促进骨骼肌成肌细胞激活增殖,促进卫星细胞库储备增加。2)红景天苷对骨骼肌成肌细胞体外分化增殖的影响存在剂量依赖性,以50 μg红景天苷干预能达到显著效果。3)红景天苷可能是通过促进TGF-β/Smad通路的关键介导子Smad2和Smad3(尤其是Smad3)磷酸化来发挥作用。
红景天苷;C2C12细胞; 成肌分化; TGF-β/Smad通路
骨骼肌损伤在运动医学领域极为常见,因其愈合质量不稳定而成为影响运动员运动寿命的重要因素。骨骼肌成肌细胞,即骨骼肌卫星细胞,经分裂融合为多核肌纤维,形成肌小管,最终分化为成熟的骨骼肌细胞[15],其作为骨骼肌再生中新生肌肉的来源在骨骼肌损伤后的修复中起着非常关键的作用[6]。在损伤的骨骼肌中,卫星细胞激活并通过增殖分化融合为新的骨骼肌细胞来修复受损的骨骼肌纤维。在临床上,外源性骨骼肌成肌细胞能较好的融入体骨骼肌细胞中[16],所以,通过成肌细胞移植来治疗运动性损伤具有广阔的应用价值和前景。然而,因为成肌细胞易于分化、难于扩增的特点,很大程度上影响种子细胞批量获取,从而阻碍成肌细胞移植的临床应用[5,18]。因此,研究成肌细胞体外分化的调控对于骨骼肌运动性损伤的治疗具有重要的理论与应用意义。目前国内、外公认的成肌细胞体外分化调控因子主要有胰岛素样生长因子-1( IGF-1)[13]和肝细胞生长因子( HGF)[7]等,但他们主要通过促生长来实现对成肌细胞体外分化的抑制,而且他们公认的成瘤作用也限制了其在临床上的应用[23]。因此,寻求更为有效的成肌细胞体外调控因子,仍然是该领域研究的一个重要课题。C2C12细胞是由C3H小鼠骨骼肌卫星细胞永生化而来的细胞株,因为细胞形态特性均一,可无限代培养,常被用作为骨骼肌卫星细胞发育分化的研究对象。
红景天苷是红景天最主要的有效活性成分,其除了有利于抗疲劳、抗衰老、免疫调节、清除自由基、增强记忆、改善睡眠等外,还能保护心脏、抵抗辐射对人体的伤害、保护造血系统等,对人体亚健康的改善和某些疾病的防治具有重要作用[1,20,21]。目前,有关红景天/红景天苷在离体细胞培养中的影响,国内外的研究主要包括红景天/红景天苷对脂肪细胞[19]、红细胞[22]、淋巴细胞[17]、肝细胞和血管平滑肌细胞[3]等影响的研究。而关于红景天/红景天苷对骨骼肌细胞的影响,目前国内、外研究极少,仅有个别聚焦于对骨骼肌细胞物质代谢方面影响的研究[12]。迄今为止,未见红景天/红景天苷在骨骼肌细胞增殖分化中作用及机制研究的报道。
基于以上研究背景,本研究在红景天苷对骨骼肌成肌细胞体外增殖分化的影响中展开研究,并提出假设:红景天苷可能通过调节TGF-β/Smad信号通路来作用于成肌细胞。本研究不仅在成肌细胞增殖分化外源性干预的基础研究方面有所创新,而且,有望为调控成肌细胞增殖分化潜能寻找到新的外源性化合物,从而促进成肌细胞的体内移植及临床应用。
1 材料与方法
1.1 实验对象与分组
成肌细胞株C2C12小鼠骨骼肌成肌细胞,购自中国北京协和医院细胞库。取一管C2C12细胞复苏,增殖传代到足够细胞量后统一冻存。选取同代细胞作为实验对象。
红景天苷,购自中国普思生物科技有限公司,10 mg红景天苷溶于1 ml无菌PBS配制红景天苷工作液,即红景天苷工作液浓度为10 μg/μl。
分组:对照组:1 ml分化液中加入5 μl不含红景天苷的无菌PBS;30 μg红景天苷组:1 ml分化液中加入3 μl红景天苷工作液;50 μg红景天苷组:1 ml分化液中加入5 μl红景天苷溶液。
1.2 主要仪器和试剂
细胞孵育箱(Thermo),倒置荧光显微镜(Leica),电泳槽、电转仪、电源、玻璃板等(Bio-rad),TCS SP5激光共聚焦显微镜(Leica),实时定量PCR仪(Eppendorf);DMEM、胎牛血清(Gibco),马血清(Hyclone),Pax7抗体、Myf5抗体、MyoD抗体、myogenin抗体、Smad2抗体、p-Smad2抗体、Smad3抗体、p-Smad3抗体(abcam),β-actin抗体、辣根酶标记山羊抗小鼠IgG、辣根酶标记山羊抗兔IgG(北京中杉金桥生物技术有限公司),Myosin-neonatal(E-MHC)抗体(SANTA)。
1.3 实验方法
1.3.1 成肌细胞株C2C12细胞增殖和诱导分化培养
C2C12细胞采用细胞生长液(高糖DMEM、10%FBS、青霉素 100 U/ml以及链霉素 100μg/ml),于5%CO2、37℃细胞孵育箱中孵育。培养过程中,维持细胞密度在70%~90%的状态,依细胞生长状态1~2天传代一次。
实验用细胞以16.5×104个/皿的密度接种于35 mm培养皿,待细胞生长至70%~80%密度时将细胞生长液更换为细胞分化液(高糖DMEM、2%HS、青霉素100 U/ml和链霉素100 μg/ml),诱导分化,每隔24 h更换分化液。每天在显微镜下观察多核肌管形成。
1.3.2 免疫荧光组化方法检测C2C12细胞诱导分化后肌管融合率
细胞诱导分化120 h后,1 ml 4%多聚甲醛固定细胞30 min,1 ml/皿 1/100 PBST破膜5 h,5%的羊血清封闭1 h;5%的羊血清配制一抗E-MHC和myogenin一抗浓度分别为1/100和1/200,过夜;5%的羊血清工作液配制二抗,山羊抗小鼠绿色二抗,二抗浓度均为1/200,二抗2 h;配制的DAPI工作液加入细胞培养conforcal皿500 ml/皿,避光储存于4℃。
激光共聚焦显微镜拍摄图像,导入Image-Pro Plus 6.0软件统计肌细胞融合率和肌细胞融合面积。
1.3.3 RNA提取与反转录及Real-Time PCR方法检测相关基因表达
细胞诱导分化过程中,每隔24 h收集细胞提取RNA并检测Pax7、Myf5、Myod和myogenin基因表达,即选取分化0 h、24 h、48 h、72 h和96 h细胞进行基因检测。
RNA提取与反转录1 ml TRIzol/皿使细胞消化裂解,细胞悬液吸入2 ml EP管裂解25 min,4℃离心取上清,加入氯仿200 μl/管,剧烈振荡摇晃15 s后静置30 min,4℃离心后取400 ml上端无色部分,加入等体积异丙醇-20℃放置20~30 min,4℃离心留沉淀,分光光度计测浓度,根据浓度采用试剂盒进行反转录,所得cDNA放置-20℃储存待用。
Real-Time PCR 根据NCBI数据库序列利用DNAMAN软件设计引物:Pax7 上游5′-GGCACAAGATAGTGGAAATGG-3′,下游5′- CGACTGCGACGCTGCTTAC-3′;Myf5 上游5-CCAGCCCCACCTCCAACT-3,下游5-GACCAGACAG-GGCTGTTACATTC-3;MyoD 上游5′-TGGCAGTGAGCACTACAGC-3′,下游5′-CATCTGGCAAAAGCAGCGAAG-3′;myogenin 上游5-GACCCTACAGACGCCCACAA-3,下游5-CCGTGATGCTGTCCACGAT-3;18S 上游5′GCACACATACATGCACGAA3′,下游5′AAAATGAAAGCCCATAGC-C3′; GAPDH 上游5-TTGTGATGGGTGTGAACCACGAGA-3,下游5- CATGAGCCCTTCCACAATGCCAAA -3;以cDNA 2 μl、上游引物和下游引物各1 μl、SYBR Green酶25 μl和注射用水21 μl的反应体系95℃预热15 min,95℃15 s、62℃15 s、72℃20 s,50个循环,72℃延伸10 min的反应条件进行PCR,采用2-△△Ct进行数据处理。
1.3.4 蛋白提取及Western Blotting 方法检测相关蛋白表达
蛋白提取 细胞诱导分化96 h后,将细胞刮下吸入2 ml EP管,4℃离心留沉淀,加入配置好的含蛋白裂解液、蛋白酶抑制剂和磷酸酶抑制剂的混合液150 μl/管,混匀,置冰上裂解25 min,期间不断吹打细胞沉淀;裂解好的蛋白液10 μl加入稀释过滤好的蛋白定性液,在提前制作的标准曲线上用分光光度计检测蛋白浓度;剩余140 μl蛋白液中加入等体积2×loading buffer,100℃加热10 min,4℃离心取上清。提取的蛋白样本储存于-20℃待用。
Western Blotting 蛋白上样量为30 μg,经100 V电压电泳和60 V电压电转后取出NC膜,立春红预染,5%的脱脂牛奶封闭,Pax7、Myf5、MyoD、myogenin、Smad2、p-smad2、Smad3和p-smad3封闭2 h,beta-Actin封闭过夜;5%的脱脂牛奶封闭液配制一抗,Pax7、Myf5、 Smad2、p-smad2、Smad3和p-smad3一抗浓度为1/500,MyoD和 myogenin一抗浓度为1/1 000,均为一抗过夜;beta-Actin一抗浓度为1/3 000,一抗2 h;5%的脱脂牛奶封闭液配制二抗,Pax7、myogenin和beta-Actin为鼠原一抗,采用山羊抗小鼠二抗,抗体浓度分别是1/500、1/1 000、1/3 000;Myf5、MyoD、Smad2、p-smad2、Smad3和p-smad3为兔原一抗,采用山羊抗兔二抗,抗体浓度均为1/1 000;X射线曝光胶片显影。
显影后的胶片扫描导入Image-Pro Plus 6.0进行灰度分析。
1.4 统计学处理
2 实验结果
2.1 不同剂量红景天苷干预对C2C12分化中汇聚、融合和形成多核肌管的影响
采用相差显微镜观察诱导分化120 h对照组、30 μg红景天苷组和50 μg红景天苷组细胞并随机选取视野进行拍照(图1),观察对照组和不同剂量红景天苷组细胞分化状态形态学差异(bar=100 μm)。
由图1可见,对照组C2C12细胞诱导分化120 h后有明显粗大的肌管形成,细胞汇聚、融合状态好。而与对照组相比,30 μg红景天苷组细胞也有细长肌管形成,虽然肌管体积较对照组小,但仍可见细胞开始融合,有较多肌管形成,而且细胞增殖状态不明显;而50 μg红景天苷组细胞成肌分化很大限度的被抑制,仅可见少量细小肌管形成,细胞汇聚融合不明显,细胞出现明显增殖。这从形态学上显示了红景天苷能抑制C2C12细胞分化并促进增殖,且存在剂量依赖性,以50 μg红景天苷干预时成肌抑制效果最明显。
2.2 不同剂量红景天苷对细胞融合率的影响
E-MHC( Myosin-neonatal )为新生肌球蛋白,是反应细胞分化状态及细胞融合率的标志性指标。
图2、表1显示,细胞进行E-MHC免疫荧光染色,随机选取3个14 mm2视野统计平均数,对照组E-MHC阳性肌管面积达到9.88 mm2,细胞融合率达到71%。而与对照组相比,30 μg红景天苷组细胞E-MHC阳性肌管面积为7.98 mm2,细胞融合率依然达到58%;50 μg红景天苷组细胞E-MHC阳性肌管面积仅2.67 mm2,细胞融合率仅19%。
myogenin为成肌调节因子家族的主要成员,是评价成肌分化状态以及肌管形成的重要指标。

图1 本研究不同剂量红景天苷在形态学上对C2C12成肌分化的影响细胞图
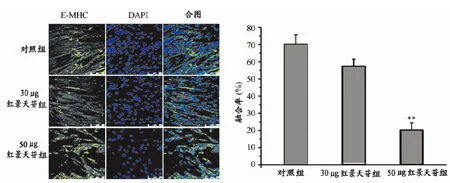
图2 本研究红景天苷对E-MHC表达的影响细胞图及柱状图
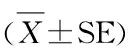
表1 本研究 E-MHC阳性面积及融合率一览表
注:*P<0.05; **P<0.01。下同。
从图3、表2细胞进行myogenin免疫荧光染色可见,对照组C2C12细胞分化120 h后形成明显粗大的肌管,myogenin阳性细胞核占视野内总细胞核的48%,30 μg红景天苷持续作用后,仍有肌管形成,myogenin阳性细胞核占视野内总细胞核的35%,而50 μg红景天苷持续作用后,仅有少量细小肌管形成,myogenin阳性细胞核仅占视野内总细胞核的25%。
同时,由以上相差显微镜观察和免疫荧光染色结果可见,红景天苷对C2C12骨骼肌细胞增殖分化的影响存在剂量依赖性,与30 μg红景天苷相比,50 μg红景天苷干预时作用更明显。因此,后续成肌分化特异基因和蛋白检测选取对照组和50 μg红景天苷组进行检测。
2.3 红景天苷(50 μg)对成肌分化特异基因表达的影响
表3、图4显示,随着分化时间的延长,Pax7基因表达水平逐渐下降,与对照组相比,红景天苷组Pax7下降幅度更大,且在各分化时程,其表达水平均低于对照组。在分化48 h时,红景天苷组和对照组差异最显著,到诱导分化96 h时,红景天苷组和对照组Pax7基因表达水平无显著差异;在诱导分化24 h和48 h时,对照组和红景天苷组Myf5表达无显著差异,对照组在分化48 h时,Myf5表达出现峰值,而红景天苷组Myf5的峰值出现在72 h,且远高于对照组,约为对照组的2.5倍,至分化96 h时,红景天苷组表达水平依然显著高于对照组(P<0.01);随着分化时长的增加,MyoD和myogenin表达水平都呈现递增趋势,其中对照组MyoD表达水平峰值出现在分化96 h(约为0 h表达水平的6倍),而myogenin表达水平峰值出现在分化72 h(约为0 h表达水平的19倍),且到分化96 h仍然高水平表达(约为0 h表达水平的15倍)。在诱导分化的任何时段,红景天苷组细胞MyoD和myogenin表达水平均低于对照组,而2组间MyoD表达水平差异在分化96 h时最显著,myogenin表达水平差异在分化72 h时最显著。

图3 本研究红景天苷对myogenin表达的影响细胞图及柱状图
表2 本研究myogenin阳性核数目一览表
表3 本研究红景天苷干预对成肌分化不同时程相关基因表达的影响一览表
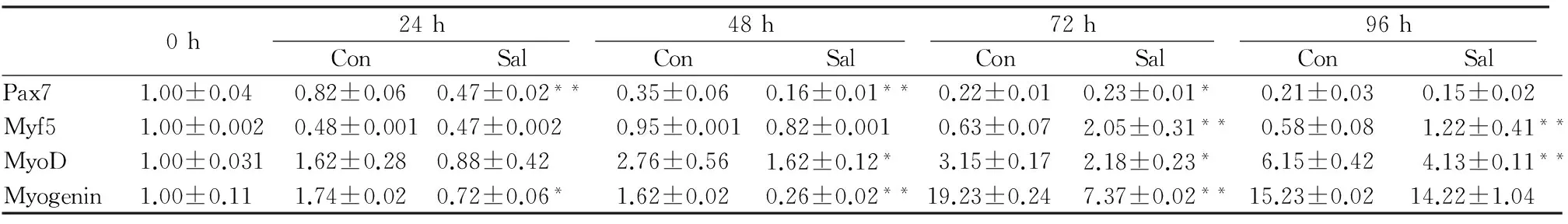
0h24h48h72h96hConSalConSalConSalConSalPax71.00±0.040.82±0.060.47±0.02**0.35±0.060.16±0.01**0.22±0.010.23±0.01*0.21±0.030.15±0.02Myf51.00±0.0020.48±0.0010.47±0.0020.95±0.0010.82±0.0010.63±0.072.05±0.31**0.58±0.081.22±0.41**MyoD1.00±0.0311.62±0.280.88±0.422.76±0.561.62±0.12*3.15±0.172.18±0.23*6.15±0.424.13±0.11**Myogenin1.00±0.111.74±0.020.72±0.06*1.62±0.020.26±0.02**19.23±0.247.37±0.02**15.23±0.0214.22±1.04

图4 本研究红景天苷对成肌分化相关基因表达的影响柱状图
2.4 红景天苷(50 μg)对成肌分化特异蛋白表达的影响
图5、表4显示,C2C12细胞诱导分化96 h后,红景天苷组细胞Pax7、MyoD和myogenin蛋白表达水平均低于对照组,而Myf5表达水平则显著高于对照组,红景天苷组Myf5表达水平约为对照组的2倍。
表4 本研究红景天苷干预对成肌分化相关蛋白表达的影响一览表
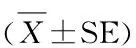
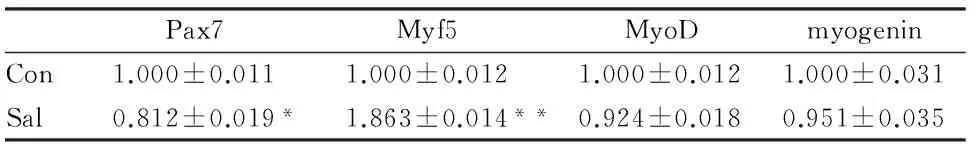
Pax7Myf5MyoDmyogeninCon1.000±0.0111.000±0.0121.000±0.0121.000±0.031Sal0.812±0.019*1.863±0.014**0.924±0.0180.951±0.035
2.5 红景天苷处理对C2C12成肌分化中TGF-β/Smad信号通路的影响
图6、表5显示,p-Smad3的表达水平随着红景天苷剂量的上升而不断递增,在30 μg红景天苷干预下,其表达略有上升,但是,无显著性差异,到50 μg红景天苷干预后,其表达激增至对照组的2.1倍,差异非常显著;而p-Smad2在30 μg红景天苷作用下,表达量略低于对照组, 50 μg红景天苷作用后,其表达增至对照组的1.4倍,差异非常显著,但响应强度不及p-Smad3。Smad2和Smad3表达量则随剂量增加递减,这与p-Smad2和p-Smad3的表达水平递增趋势是相呼应的。

图5 本研究红景天苷对成肌分化相关蛋白表达的影响电泳及柱状图
表5 本研究不同剂量红景天苷对TGF-β/Smad信号通路的影响一览表
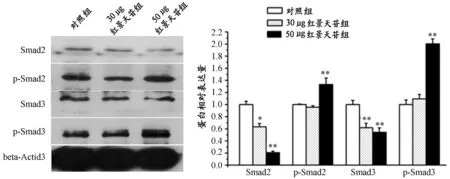
图6 本研究不同剂量红景天苷对TGF-β/Smad信号通路的影响电泳及柱状图
3 分析讨论
3.1 红景天苷对C2C12细胞体外分化的影响
本研究从形态学,细胞融合率,成肌分化特异基因和蛋白表达等几个方面探索不同浓度红景天苷对C2C12骨骼肌成肌细胞体外分化的影响。本研究以体外培养的成肌细胞株C2C12为研究对象,基本可以排除自体因素的干扰,更能直接说明红景天苷对骨骼肌成肌细胞的影响。
在形态学上,50 μg红景天苷能明显抑制C2C12细胞融合、汇聚和多核肌管的形成,且在诱导分化过程中仍然能明显促进骨骼肌细胞增殖。可以推断,红景天苷在抑制C2C12骨骼肌成肌细胞分化的同时,也能促进骨骼肌成肌细胞增殖,增加骨骼肌卫星细胞库储备。而30 μg红景天苷对成肌细胞的分化有一定的抑制,但效果并不如50 μg红景天苷明显,且没有观察到细胞增殖的迹象。因此,也可以从形态学上推断,红景天苷对成肌细胞的影响存在剂量依赖性,50 μg红景天苷效果更显著。
E-MHC为新生肌球蛋白,是反映细胞分化状态及细胞融合率的标志性指标。正常情况下,在细胞增殖期内E-MHC表达极少,而在细胞分化过程中,随着细胞分化时间的延长,细胞不断融合汇聚,肌管数目增多面积增大,则E-MHC阳性表达增加,细胞融合率也上升。myogenin为成肌调节因子家族的重要成员,调节着成肌细胞进一步终末分化为肌管肌纤维。可以把myogenin作为评价成肌分化状态以及肌管形成的重要指标[2]。本研究中E-MHC和myogenin免疫荧光染色结果可见,50 μg红景天苷显著降低细胞融合率,抑制粗大肌管生成,影响成肌分化,而30 μg红景天苷虽产生一定影响,但与对照组无显著差异。此时可以确定,与30 μg红景天苷干预相比,50 μg红景天苷能更有效的调控骨骼肌成肌细胞体外增殖分化以达到本研究的目的。因此,在接下来的研究中选取50 μg红景天苷干预浓度来进行成肌分化特异基因和蛋白的检测,以进一步验证红景天苷对骨骼肌成肌细胞体外增殖分化的影响。
Pax7是在成肌细胞中表达的标志性蛋白[11],Myf5、MyoD和myogenin同属于MRFs家族,是成肌细胞增殖分化的关键调控因子。静止的骨骼肌卫星细胞剧烈运动或受到创伤时,会大量激活Pax7,Pax7和MRFs家族相互作用,促进肌细胞增殖和分化,Myf5是MRFs家族中最早表达的蛋白,卫星细胞激活后,大量增殖并伴有Myf5和MyoD的表达,成肌细胞进入分化期后,Pax7和Myf5表达大量下降并开始表达myogenin[4,9,10]。简而言之,Pax7可以看作是静止状态成肌细胞增殖水平的标志蛋白,Myf5可以看作是激活状态成肌细胞增殖水平的标志蛋白,MyoD和Myogenin都可以看作是激活状态成肌细胞分化水平的标志蛋白,但MyoD先于myogenin表达。本研究通过每隔24 h实时检测以上4种基因在诱导分化成肌细胞中的表达验证了这一结论,即Pax7基因表达峰值出现在诱导分化即刻、Myf5基因表达峰值出现在诱导分化48 h和72 h之间、MyoD和myogenin基因表达峰值出现在诱导分化96 h左右(MyoD表达随分化时间的延长而逐渐递增,而myogenin在诱导分化初期表达极少,到分化72 h后出现大幅迅猛增长)。红景天苷作用后,Pax7基因和蛋白表达水平都显著低于对照组,Myf5基因和蛋白表达显著高于对照组,可知红景天苷不仅能有效促进静止态细胞的激活,且还能促进被激活卫星细胞的增殖,从而增加卫星细胞库的储备;在整个细胞诱导分化阶段,红景天苷干预后,MyoD和myogenin的基因表达水平均低于对照组,诱导分化96 h时,蛋白表达水平也低于对照组。
至此可以得出结论,红景天苷能有效促进骨骼肌成肌细胞的体外激活增殖,抑制骨骼肌成肌细胞的体外分化,从而促进卫星细胞库储备的增加。本结果一方面为成肌细胞的体内移植及临床应用奠定实验基础,提示,可以考虑将红景天苷作为骨骼肌成肌细胞体外增殖分化的外源性调控物,来解决成肌细胞自身易分化难扩增的问题,促进种子细胞的批量获取,进而推动成肌细胞移植的临床应用,达到通过成肌细胞移植来治疗骨骼肌损伤的目的;另一方面,为开拓红景天苷在体内骨骼肌损伤再生和修复领域的应用提供理论和实验基础,提示,红景天苷或许也可以促进体内骨骼肌卫星细胞库的储备,增加骨骼肌再生中新生肌肉的来源,从而有效帮助受损骨骼肌的体内再生和修复,不过这需要进一步的体内研究来佐证。
3.2 红景天苷影响C2C12细胞体外分化机制初探
TGF-β/Smad通路是由转化生长因子 β(transforming growth factor-β, TGF-β)、TGF-β受体(transforming growth factor-β receptor, TβR)以及受体底物Smad蛋白家族组成,在众多细胞调节过程中具有非常关键的作用。在成人骨骼肌中TGF-β/Smad通路可抑制卫星细胞的分化、肌纤维的融合以及特定基因的表达,Smad2和Smad3是该过程的关键调控因子[8,14]。因此,结合以上红景天苷抑制成肌细胞分化的结论,猜想红景天苷可能通过作用于TGF-β/Smad通路来影响成肌细胞增殖分化。所以,本研究检测了不同浓度红景天苷干预96 h后C2C12细胞中TGF-β/Smad信号通路的关键介导子Smad2和Smad3的磷酸化水平。由实验结果可知,Smad2和Smad3的磷酸化水平(尤其是Smad3)与红景天苷剂量变化和红景天苷成肌抑制作用均为正相关,即50 μg红景天苷作用于诱导分化的骨骼肌成肌细胞时,在抑制成肌分化促进细胞增殖的同时,也可以检测到Smad2和Smad3磷酸化水平的显著增加,且同时还检测到Smad2和Smad3表达水平与其磷酸化水平呈负相关。这说明,红景天苷可能是通过激活Smad2和Smad3蛋白磷酸化(主要是Smad3蛋白)来发挥成肌抑制作用,而没有增加总的Smad2和Smad3蛋白含量。不过该论断只是初步的探索,更确定的机制研究还有待后续进一步的研究,同时,30 μg红景天苷对Smad2和Smad3磷酸化的作用远不如50 μg红景天苷,与对照组无显著差异,这也进一步验证了本研究中关于红景天苷最佳作用浓度的结论。
4 结论
1.红景天苷能显著抑制骨骼肌成肌细胞体外成肌分化,并促进骨骼肌成肌细胞激活增殖,促进卫星细胞库储备增加。
2.红景天苷对骨骼肌成肌细胞体外分化的影响存在剂量依赖性,以50 μg红景天苷干预能达到显著效果。
3.红景天苷可能是通过促进TGF-β/Smad通路的关键介导子Smad2和Smad3(尤其是Smad3)磷酸化来发挥作用。
限于本研究条件有限,一些问题在所难免,需要更多后续研究来完善和证明:1)红景天苷对骨骼肌成肌细胞体外增殖分化的效果是否和其浓度存在线性关系,若浓度增加到50 μg以上是否会产生更好的效果;2)关于TGF-β/Smad通路在红景天苷的成肌调控中的作用还需进一步研究证实。
[1]杨卓,龙旭,兰泽伦.圣地红景天研究进展[J].现代临床医学,2011,37(4):243-244.
[2]曾缨,张成,刘克玄,等.成肌调节因子MyoD与myogenin在肌肉损伤修复过程的动态变化[J].第一军医大学学报,2004,24(5):542-545.
[3]张华,戴立里,曾维政,等.红景天甙对大鼠肝组织TGF-β1/Smads信号通路的作用[J].重庆医科大学学报,2011,36(6):699-703.
[4]BEYLKIN D H,ALLEN D L,LEINWAND L A.MyoD,Myf5,and the calcineurin pathway activate the developmental myosin heavy chain genes[J].Dev Biol,2006,294(2):541-553.
[5]BICK-FORRESTER J,LEE M S,MAKKAR R R,etal.Partial restoration of myocardial function and perfusion by cell therapy following myocardial infarction[J].Curr Opin Cardiol,2004,19(6):631-637.
[6]CHARGE S B,RUDNICKI M A.Cellular and molecular regulation of muscle regeneration[J].Physiol Rev,2004,84(1):209-238.
[7]DO M K,SUZUKI T,GERELT B,etal.Time-coordinated prevalence of extracellular HGF,FGF2 and TGF-beta3 in crush-injured skeletal muscle[J].Anim Sci J,2012,83(10):712-717.
[8]DROGUETT R,CABELLO-VERRUGIO C,SANTANDER C,etal.TGF-beta receptors,in a Smad-independent manner,are required for terminal skeletal muscle differentiation[J].Exp Cell Res,2010,316(15):2487-2503.
[9]GAYRAUD-MOREL B,CHRETIEN F,FLAMANT P,etal.A role for the myogenic determination gene Myf5 in adult regenerative myogenesis[J].Dev Biol,2007,312(1):13-28.
[10]KABLAR B,KRASTEL K,TAJBAKHSH S,etal.Myf5 and MyoD activation define independent myogenic compartments during embryonic development[J].Dev Biol,2003,258(2):307-318.
[11]KUANG S,CHARGE S B,SEALE P,etal.Distinct roles for Pax7 and Pax3 in adult regenerative myogenesis[J].J Cell Biol,2006,172(1):103-113.
[12]LI H B,GE Y K,ZHENG X X,etal.Salidroside stimulated glucose uptake in skeletal muscle cells by activating AMP-activated protein kinase[J].Eur J Pharmacol,2008,588(2-3):165-169.
[13]LIU T B,FEDAK P W,WEISEL R D,etal.Enhanced IGF-1 expression improves smooth muscle cell engraftment after cell transplantation[J].Am J Physiol Heart Circ Physiol,2004,287(6):H2840-2849.
[14]MENG X M,HUANG X R,CHUNG A C,etal.Smad2 protects against TGF-beta/Smad3-mediated renal fibrosis[J].J Am Soc Nephrol,2010,21(9):1477-1487.
[15]NAGATA Y,PARTRIDGE T A,MATSUDA R,etal.Entry of muscle satellite cells into the cell cycle requires sphingolipid signaling[J].J Cell Biol,2006,174(2):245-253.
[16]PAGANI F D,DERSIMONIAN H,ZAWADZKA A,etal.Autologous skeletal myoblasts transplanted to ischemia-damaged myocardium in humans.Histological analysis of cell survival and differentiation[J].J Am Coll Cardiol,2003,41(5):879-888.
[17]SKOPINSKA-ROZEWSKA E,SOKOLNICKA I,SIWICKI A K,etal.Dose-dependent in vivo effect of Rhodiola and Echinacea on the mitogen-induced lymphocyte proliferation in mice[J].Pol J Vet Sci,2011,14(2):265-272.
[18]TAMBARA K,SAKAKIBARA Y,SAKAGUCHI G,etal.Transplanted skeletal myoblasts can fully replace the infarcted myocardium when they survive in the host in large numbers[J].Circulation,2003,10( 1):259-263.
[19]WANG S H,WANG W J,WANG X F,etal.Effects of salidroside on carbohydrate metabolism and differentiation of 3T3-L1 adipocytes[J].Zhong Xi Yi Jie He Xue Bao,2004,2(3):193-195.
[20]YU S,LIU M,GU X,etal.Neuroprotective effects of salidroside in the PC12 cell model exposed to hypoglycemia and serum limitation[J].Cell Mol Neurobiol,2008,28(8):1067-1078.
[21]ZHANG L,YU H,SUN Y,etal.Protective effects of salidroside on hydrogen peroxide-induced apoptosis in SH-SY5Y human neuroblastoma cells[J].Eur J Pharmacol,2007,564(1-3):18-25.
[22]ZHENG K Y,GUO A J,BI C W,etal.The extract of Rhodiolae Crenulatae Radix et Rhizoma induces the accumulation of HIF-1alpha via blocking the degradation pathway in cultured kidney fibroblasts[J].Planta Med,2011,77(9):894-899.
[23]ZULOAGA R,FUENTES E N,MOLINA A,etal.The cAMP response element binding protein (CREB) is activated by insulin-like growth factor-1 (IGF-1) and regulates myostatin gene expression in skeletal myoblast[J].Biochem Biophys Res Commun,2013,440(2):258-264.

Effect of Different Concentrations of Salidroside on Myoblast Differentiation in Vitro and Its Preliminary Mechanism
LUO Wei1,ZHANG Peng2,AI Lei3,ZHOU Yue4,CHEN Xiao-ping2
Objective:This study is aim to explore the effect of different concentrations of salidroside on myoblast differentiation in vitro and its possible mechanism,which can help to find a more effective exogenous regulating substance for myoblast differentiation to promote the myoblast transplantation,and set the foundation for further study of the effect of salidroside on skeletal muscle in vivo.Methods:Part one:The effect of different concentrations of salidroside on myoblast differentiation in vitro:1)observe the morphology of differential myoblast C2C12 which were treated with 0,30,50 μg/ml salidroside to induce differentiation by Phase micrographs;2)Detect the cell fusion index affected by salidroside using immunofluorescence;3)Detect the expressions of myogenic differentiation-specific mRNAs and proteins affected by salidroside using Western Blotting and real time-PCR.Part two:The possible mechanism of above study :Detect TGF-β/Smad3 pathway protein expression in differential C2C12 cell treated with 0,30,50μg/ml Sal using Western Blotting. Results:Treated with 50 μg/ml salidroside:Part one:1)Salidroside inhibit C2C12 fusion and myotube form in morphological by Phase micrographs;2)Myosin-neonatal Positive area and myogenin Positive Nuclei decreased significantly by immunofluorescence(P<0.05),which inhibited myotube form notably;3)The expressions of MyoD and myogenin in mRNA and protein both decreased notably by real-time PCR and Western blotting(P<0.05),which inhibited cell fusion and myotube form during C2C12 differentiation.Part two:Salidroside inhibit C2C12 differentiation associated with increases in Smad2/Smad3 phosphorylation,especially p-Smad3(P<0.01).The results treated with 30 μg/ml salidroside is insignificant (P>0.05). Conclusion:1)Salidroside inhibit myogenic differentiation of C2C12 cell significantly,and promote cell proliferation,to add satellite cells band reserve.2)Salidroside inhibits myogenic differentiation in a dose-dependent manner,50μg/ml Salidroside is the best.3)Salidroside inhibits myogenic differentiation possibly by activating Smad2/Smad3(a key mediator in TGF-β/Smad pathway) phosphorylation.
Salidroside;C2C12cells;myogenicdifferentiation;TGF-β/Smadpathway
2015-03-24;
2015-08-10
国家自然科学基金面上项目(81272177);南京体育学院科研项目(YJ1423)。
罗维(1989-),女,湖北荆门人,硕士,主要研究方向为运动生理学,E-mail:luoweihyd@163.com。
1.南京体育学院 运动健康科学系,江苏 南京 210014;2.中国航天员科研训练中心,航天医学基础与应用国家重点实验室,北京 100094;3.江苏省体育科学研究所,江苏 南京 210033;4.北京体育大学,北京 100084 1.Nanjing Sport Institute,Nanjing,210014,China;2.State Key Laboratory of Space Medicine Fundamentals and Application,China Astronaut Research and Training Center,Beijing 100094,China;3.Jiangsu Research Institute of Sports Science,Nanjing 210033,China;4.Beijing Sport University,Beijing 100084,China.
1000-677X(2015)09-0050-08
10.16469/j.css.201508000
G804.5
A
