hsa-miR-30a在人骨肉瘤细胞U2-OS中的表达及其对细胞凋亡的影响
2014-12-01丁江水邓英虎
丁江水,邓英虎
(1.青阳县人民医院 骨科,安徽 池州 242800;2.铜陵市人民医院 骨科,安徽 铜陵 244000)
目前骨肉瘤治疗方法通常为手术治疗并辅以化疗,5年生存率为50% ~80%[1]。但迄今为止我们仍不完全清楚骨肉瘤的分子发病机理。微小RNA(microRNA,miRNA)是一种高度保守的长度约22~24个碱基的非编码单链小RNA,它能通过与互补的mRNA序列结合,从而导致目的mRNA的降解和抑制蛋白的翻译[2]。研究表明,有98种miRNA基因定位于与肿瘤相关的脆弱位点或基因组区域[3-4]。它通过调控靶基因参与信号通路,从而影响肿瘤的细胞周期、细胞凋亡、细胞迁移和血管生成,影响肿瘤的发生、发展和预后[5-6]。
近期研究发现,miR-30a在人多种正常组织中均高表达,但在非小细胞肺癌、结肠癌、乳腺癌等恶性肿瘤组织中表达下降或不表达,揭示miR-30a在恶性肿瘤中是一种潜在的抑癌基因[8-10]。本研究检测了人骨肉瘤组织中miR-30a的表达水平,构建miR-30a前体的真核表达载体并转染至人骨肉瘤细胞U2-OS中,应用荧光定量PCR检测了miR-30a在U2-OS细胞中的表达,和流式细胞技术检测稳转株的凋亡情况,分析miR-30a对骨肉瘤细胞凋亡的影响。
1 材料与方法
1.1 主要材料 人骨肉瘤组织和正常肌肉组织由铜陵市人民医院和青阳县人民医院骨科提供。所有组织标本的处理均经过铜陵市人民医院或青阳县人民医院伦理委员会讨论通过。RIPA-1640培养基(Gibco)、南美胎牛血清(Gibco)、转染用无血清培养基 OPTI-MEM(Gibco),Trizol(Invitrogen)、Purelink HiPure Plasmid Miniprep Kit(Invitrogen)、X-treme GENE HP DNA transfection reagent(Roche)、Cell Counting Kit-8(日本同仁)、miScript Reverse Transcription Kit(Qiagen)、miScript SYBR Green PCR Kit(Qiagen)、miScript Primer Assays(MS00007350)(Qiagen)、Guava Nexin Reagent(Millipore),premiR-30a真核表达载体由广州复能基因有限公司构建,以插入一段无效序列的载体作为无效对照。
1.2 实验方法
1.2.1 hsa-miR-30a在人骨肉瘤组织中的表达水平检测 术中将切除的骨肉瘤组织和正常肌肉组织分别剪碎,部分组织于液氮中速冻后-80℃冰箱保存备用。用Trizol一步法提取骨肉瘤组织和正常肌肉的总RNA(包括microRNA),琼脂糖凝胶电泳检测RNA质量,并测RNA浓度。按照miScript Reverse Transcription Kit说明书将细胞总RNA逆转录为cDNA。以cDNA为模板,用Qiagen miScript Primer Assays及 miScript SYBR Green PCR kit扩增 miRNA-30a片段,并使用RNU6B作为内参。反应条件为:95℃预变性 15 min,94℃变性 15 s,55℃退火 30 s,70℃荧光检测30 s,重复40个循环。70~95℃绘制溶解曲线,根据Cp值进行数据分析。
1.2.2 无内毒素质粒提取 将构建的真核表达载体pEZX-MR04-pre-miR-30a及无效对照质粒转化大肠杆菌DH5α感受态细胞,涂布于LB平板(Amp+)上,并在37℃条件下倒置培养12~16 h。次日从平板上挑取菌落至LB液体培养基(Amp+)中培养,并按Invitrogen Purelink HiPure Plasmid Miniprep Kit试剂盒说明书提取质粒,最后用NaRo Drop 2000核酸测定仪(Thermo)测定质粒浓度。
1.2.3 细胞转染 U2-OS细胞用含10%胎牛血清的RIPA-1640培养基置于37℃、5%CO2的恒温培养箱中培养。向6孔板各孔中加入浓度为1×105细胞/ml的U2-OS细胞2 ml,培养24 h待贴壁细胞达到70% ~80%汇合度时,按照Roche X-treme GENE HP DNA转染试剂说明书进行质粒转染,设无效对照组,转染试剂对照组。转染后16 h更换含10%胎牛血清的RIPA-1640培养基,继续培养48 h后检测miR-30a的表达。
1.2.4 检测 hsa-miR-30a在 U2-OS细胞中的表达pEZX-MR04载体中有eGFP标签,发绿色荧光,所以用荧光显微镜观察 eGFP的表达情况,可检测miR-30a的表达。另外,可用荧光定量PCR检测U2-OS细胞miR-30a的表达。在转染pEZX-MR04-pre-miR-30a载体48 h后,用Trizol一步法提取细胞总RNA,并反转录成cDNA,然后进行荧光定量PCR检测,操作步骤同 1.2.1。
1.2.5 流式细胞术检测U2-OS细胞的凋亡 U2-OS细胞于6孔板转染pEZX-MR04-pre-miR-30a载体72 h后,用胰酶消化液将细胞消化下来,800 g离心后用100 μl含10%胎牛血清的RIPA-1640培养基轻轻重悬,然后加入100 μl Guava Nexin Reagent,避光孵育20 min。然后上Guava easyCyte 6-2L流式细胞仪(Millipore)检测细胞凋亡情况。
1.2.6 统计学处理 采用SPSS 13.0软件统计分析,实验数据以s表示,荧光定量PCR结果采用方差分析,两两比较用SNK-q检验,P<0.05为差异有统计学意义。实验数据均重复3次。
2 结果
2.1 miR-30a在人骨肉瘤组织和U2-OS细胞中的表达 与5例正常肌肉组织相比,5例骨肉瘤组织的miR-30a表达水平降低(P<0.01,图1)。骨肉瘤细胞U2-OS在转染pre-miR-30a表达载体和无效载体后,荧光显微镜下可见大量发绿色荧光的阳性细胞,转染效率高达90%以上(图2)。pre-miR-30a转染组细胞的miR-30a表达水平显著增高,与无效对照组及转染试剂对照组细胞相比,差异具有统计学意义 (P <0.01,图3)。
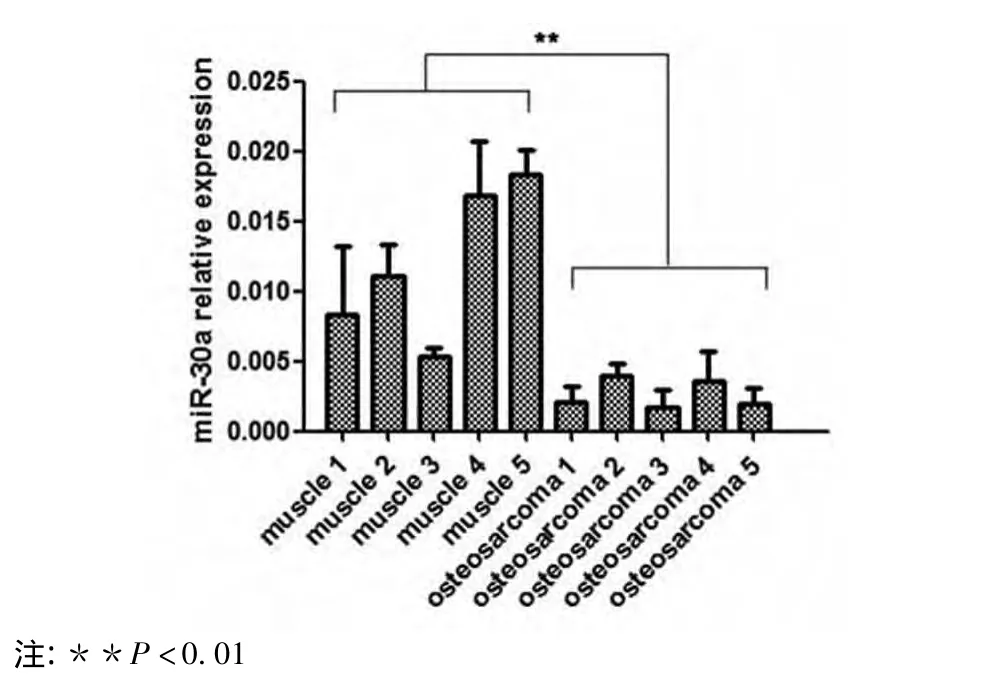
图1 miR-30a在正常肌肉组织和骨肉瘤组织中的表达水平Fig 1 Expression levels of miR-30a in normal muscle tissue and in osteosarcoma
2.2 上调miR-30a表达对骨肉瘤细胞U2-OS凋亡的促进作用 转染pre-miR-30a表达载体72 h后,骨肉瘤细胞U2-OS经Annexin V和7-AAD染色后用流式细胞仪检测,早期凋亡率分别为(26.32±2.54)%,与无效对照载体组的凋亡率(9.35±1.64)%,和转染试剂对照组细胞的凋亡率(0.21±0.06)% 相比,差异具有统计学意义(P <0.05,图4)。

图2 荧光显微镜观察U2-OS细胞转染pre-miR-30a表达载体后情况(×100)Fig 2 Expression of U2-OS cells transfected with the expression vector pre-miR-30a by fluorescence microscopy

图3 荧光定量PCR检测miR-30a在骨肉瘤细胞U2-OS中的表达Fig 3 Detection of miR-30a expression in osteosarcoma cells in U2-OS by fluorescence quantitative PCR
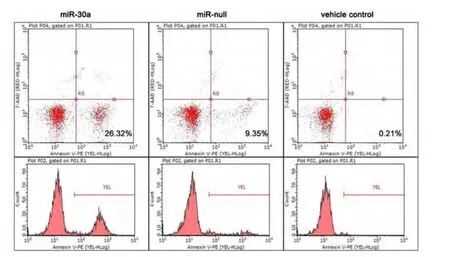
图4 流式细胞仪检测上调miR-30a表达水平后对U2-OS细胞凋亡的影响Fig 4 Effect on apoptosis of U2-OS cells in flow cytometry after up-regulated miR-30a expression level
3 讨论
miRNA是一种内源性表达的非编码单链小RNA,其前体:初级 miRNA(primary miRNA,pri-miRNA)和前体 miRNA(precursor miRNA,pre-miRNA)可在Drosha复合体和Dicer酶的作用下在胞浆中由剪切成为成熟miRNA,并与靶基因mRNA的3'非翻译区结合并降解靶mRNA,从而降低靶基因mRNA水平或直接抑制其蛋白翻译过程[2],调控生命过程的多个环节,包括器官发育、物质代谢、细胞分化、凋亡和癌变。近年来发现在乳腺癌、结肠癌、肺癌、甘肝癌等多种恶性肿瘤中部分miRNA的表达水平异常,揭示miRNA在肿瘤发生发展过程中具有重要调控作用[11-15]。
近期研究表明,miR-30a参与了多种癌细胞的增殖、凋亡、侵袭和转移等过程。Baraniskin等发现恢复miR-30a在结肠癌细胞中的表达后可抑制癌细胞的增殖,诱导细胞的凋亡,并使细胞停留在G1期,提示miR-30a可能可作为治疗结肠癌的分子靶标之一[8]。Cheng等上调miR-30a在乳腺癌细胞株中的表达水平后,细胞的迁移和侵袭能力显著降低,且miR-30a低表达的乳腺癌病人大多存在癌症晚期、淋巴结转移、高病死率和高复发率等现象[9]。Kumarswamy等还发现了miR-30a与非小细胞肺癌的上皮-间质转化过程相关,高表达miR-30a能抑制癌细胞的侵袭和转移[10]。本研究发现与正常肌肉组织相比,miR-30a在骨肉瘤组织中的表达水平明显下降,与其他肿瘤中的报道一致。本研究构建了miR-30a前体的真核表达载体并成功将其转染至骨肉瘤细胞U2-OS中,检测出骨肉瘤细胞中miR-30a的上调表达。
细胞凋亡过程是一个多步骤、多基因失调的过程,有许多蛋白参与其中。研究证实,miR-30a的靶蛋白之一为denticleless protein homolog(DTL),可诱导细胞凋亡;DTL的表达下调可增强TP53和CDKN1A的表达,从而诱导细胞凋亡及使细胞停留在G1期[8]。推测miR-30a可能直接或间接锚定其中某些抗凋亡蛋白的mRNA 3'UTR区,使蛋白合成抑制或减少,从而诱导凋亡发生。本研究中,流式细胞技术检测成功转染后的U2-OS细胞,发现骨肉瘤细胞的凋亡明显增加。
综上所述,本实验成功将miR-30a真核表达载体转染骨肉瘤细胞U2-OS,且稳转株可有效表达,研究结果表明,骨肉瘤细胞中miR-30a的过表达可明显增加U2-OS细胞的凋亡,但其具体机制尚未清楚,且miR-30a与骨肉瘤细胞增殖、侵袭等生物学行为的关系亦未见相关报道,值得进一步研究,以便为骨肉瘤的治疗提供新的靶点。
[1]OTTAVIANI G,JAFFE N.The epidemiology of osteosarcoma[J].Cancer Treat Res,2010,152:3 -13.
[2]DOENCH JG,SHARP PA.Specificity of microRNA target selection in translation repression [J].Genes Dev,2004,18(5):504 -511.
[3]CALIN GA,DUMITRU CD,SHIMIZU M,et al.Frequent deletions and down-regulation of micro-RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia [J].Proc Natl Acad Sci USA,2002,99(24):15524 -15529.
[4]CALIN GA,SEVIGNANI C,DUMITRU CD,et al.Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers[J].Proc Natl Acad Sci USA,2004,101(9):2999-3004.
[5]SKAFTNESMO KO,PRESTEGARDEN L,MICKLEM DR,et al.MicroRNAs in tumorigenesis[J].Curr Pharm Biotechnol,2007,8(6):320-325.
[6]SHI XB,TEPPER CG,DEVERE WHITE RW.Cancerous miRNAs and their regulation[J].Cell Cycle,2008,7(11):1529 -1538.
[7]BARTEL DP.MicroRNAs:genomics,biogenesis,mechanism,and function[J].Cell,2004,116(2):281 -297.
[8]ALEXANDER B,KARIN BD,ABDELOUAHID M,et al.MiR-30a-5p suppresses tumor growth in colon carcinoma by targeting DTL[J].Carcinogenesis,2012,33(4):732 -739.
[9]CHENG CHUNWEN,WANG HSIAOWEI,CHANG CHIAWEI,et al.MicroRNA-30a inhibits cell migration and invasion by downregulating vimentin expression and is a potential prognostic marker in breast cancer[J].Breast Cancer Research and Treatment,2012,134(3):1081-1093.
[10]KUMARSWAMY R,MUDDULURU G,CEPPI P,et al.MicroRNA-30a inhibits epithelial-to-mesenchymal transition by targeting Snai1 and is downregulated in non-small cell lung cancer[J].Int J Cancer,2012,130(9):2044 -2053.
[11]SIEUWERTS AM,MOSTERT B,BOLT-DE VRIES J,et al.mRNA and microRNA expression profiles in circulating tumor cells and primary tumors of metastatic breast cancer patients[J].Clin Cancer Res,2011,17(11):3600 -3618.
[12]TANG JIETING,FANG JINGYUAN.MicroRNA regulatory network in human colorectal cancer[J].Mini Rev Med Chem,2009,9(8):921-926.
[13]LI XINHUA,ZHANG GUIYING,LUO FEIJUN,et al.Identification of aberrantly expressed miRNAs in rectal cancer[J].Oncol Rep,2012,28(1):77 -84.
[14]YANG YE,LI XIAOFEI,YANG QI,et al.The role of microRNA in human lung squamous cell carcinoma[J].Cancer Genet Cytogenet,2010,200(2):127 -133.
[15]方基勇,刘影,毕富勇.靶向Survivin基因microRNA诱导肝癌细胞凋亡的实验[J].皖南医学院学报,2008,27(6):401-402.
