内源性H2S抑制angiotensin II引起的神经元活性氧水平的升高*
2014-08-08曹冬青刘小妮徐海艳金惠铭
曹冬青, 刘小妮, 徐海艳, 陶 然, 黄 莺, 金惠铭, 王 睿, 卢 宁
(复旦大学基础医学院生理与病理生理学系,上海 200032)
硫化氢(hydrogen sulfide,H2S)作为一种新的气体信号分子,对心血管系统有着重要的调节作用,参与了多种病理生理过程,如降低血压、减慢心率、抑制血管平滑肌细胞增殖等。但其对高血压的具体调节机制尚不明确。近年研究证实在高血压和心衰的发病过程中,中枢活性氧(reactive oxygen species,ROS)介导了血管紧张素II(angiotensin II,Ang II)的心血管效应。中枢ROS水平升高会导致延髓头端腹外侧(rostral ventrolateral medulla,RVLM)等区神经元放电频率增加,最终引起交感兴奋,血压升高[1-3]。ROS是机体氧化应激的主要来源之一,而H2S能够通过促进抗氧化物质谷胱甘肽的生成,降低ROS水平,保护神经元[4]。本课题组前期在整体实验证实:H2S能降低自发性高血压大鼠(spontaneously hypertensive rat,SHR)的ROS水平,继而降低血压,减慢心率[5]。同时我们在细胞水平上证实:外源性H2S可抑制Ang II引起的延髓神经元ROS水平的升高[6]。在此基础上,本工作旨在通过给予内源性H2S生成酶胱硫醚β-合成酶(cystathionine β-synthetase,CBS)的激动剂——丁酸钠(sodium butyrate,NaBu)[7],提高神经元内源性H2S水平,进一步观察内源性H2S对Ang II引起的ROS水平的影响并对可能的作用机制进行探讨。
材 料 和 方 法
1 主要试剂与仪器
Ang II、二氢乙啶(dihydroethidium,DHE)和NaBu均购自Sigma;微管相关蛋白2(microtubule-associated protein 2, MAP-2)抗体(小鼠来源和兔来源)购自Abcam;小鼠来源CBS抗体购自Abnova;神经元培养基、B27神经细胞生长添加剂、胎牛血清(fetal bovine serum,FBS)、2.5% 胰蛋白酶和DMEM/F12培养基均购自Gibco;DAPI、FITC标记山羊抗兔IgG (H+L)、Cy3标记山羊抗小鼠IgG (H+L)、蛋白裂解液和超氧化物歧化酶(superoxide dismutase,SOD)活性检测试剂盒购自碧云天;SYBR Green和逆转录试剂盒购自Toyobo;GAPDH引物和CBS引物由上海桑尼生物科技有限公司合成;二氧化碳恒温细胞培养箱(上海力申科学仪器有限公司),无菌超净台(上海博迅实业有限公司医疗设备厂),激光共聚焦系统(Zeiss),酶标仪(Tecan)等。
2 原代培养及鉴定延髓神经元
2.1原代延髓神经元的培养 孕14~16 d SD大鼠购于上海斯莱克实验动物中心,取胎鼠,在解剖显微镜下取延髓组织,剪碎,0.125% 胰酶37 ℃ 消化12 min左右,10% FBS终止消化3次,吹打3次,上清移至新的离心管,1 000 r/min 离心8 min,弃上清,加入神经元培养基与B27的混合培养液(比例为49∶1),重悬细胞。用0.4% 台盼蓝对神经元进行计数,稀释到所需密度,接种到35 mm的培养皿或96孔板,1周换2次液,每次半量换液,培养7 d左右,待神经元发育成熟,进行以下实验。
2.2原代延髓神经元的鉴定 将已培养7 d左右的延髓神经元进行鉴定,具体步骤如下:(1)0.01 mol/L PBS 洗3次;(2)4%多聚甲醛固定细胞20 min;(3)0.01 mol/L PBS洗3次;(4)5% FBS 封闭30 min;(5)加Ⅰ抗MAP-2抗体(1∶400),37 ℃ 1 h,4 ℃ 过夜;(6)0.01 mol/L PBS洗3次;(7)加Ⅱ抗Cy3(1∶200),37 ℃ 避光1 h;(8)0.01 mol/L PBS洗3次;(9)DAPI染核5 min;(10)0.01 mol/L PBS洗3次;(11)激光共聚焦系统拍照(×200)鉴定。
3 神经元胞内ROS水平的检测
本实验采用DHE荧光探针法检测延髓神经元胞内的ROS (主要为超氧阴离子)水平。神经元给药后,加DHE(5 μmol/L),37 ℃ 孵育30 min,酶标仪检测荧光强度,激发波长515 nm,发射波长585 nm。
4 总SOD活性的测定
延髓神经元预先给予Ang II(1 μmol/L)处理后,(1)用细胞裂解液裂解,细胞刮刀收集至1.5 mL离心管中,16 000×g离心10 min;(2)将上清移入新的离心管;(3)采用BCA法测蛋白浓度;(4)按试剂盒说明测定总SOD的活性。
5 CBS与MAP-2的荧光双标
将延髓神经元培养至7 d左右进行以下实验:(1)0.01 mol/L PBS 洗3次;(2)4% 多聚甲醛固定细胞20 min;(3)0.01 mol/L PBS洗3次;(4)5% FBS 封闭30 min;(5)加MAP-2抗体(1∶400)与CBS抗体(1∶400)的混合液,37 ℃ 1 h,4 ℃ 过夜;(6)0.01 mol/L PBS洗3次;(7)加抗FITC与抗Cy3混合液(均为1∶200),37 ℃ 避光1 h;(8)0.01 mol/L PBS洗3次;(9)DAPI染核5 min;(10)0.01 mol/L PBS洗3次;(11)激光共聚焦系统拍照(×200)鉴定。
6 Ang II 对CBS 表达的影响
采用real-time PCR观察Ang II对CBS mRNA表达的影响。GAPDH上游引物 5’-TGACAACTTTGGCATCGTGG- 3’,下游引物5’ -TGCAGGGATGATGTTCTGGG-3’;CBS上游引物5’ -CCCATCCTTGCAAAGCTTGT-3’,下游引物 5’ -TTGGGGTATCAGGCGAGATC-3’。
7 统计学处理
采用SPSS 17.0 统计软件分析。数据用均数±标准误(mean±SEM)表示,两组间比较应用独立样本t检验,多组间比较应用单因素方差分析(One-way ANOVA),以P<0.05为差异有统计学意义。
结 果
1 原代培养神经元的鉴定
MAP-2是神经元的特异性标志物。由于本课题组原代培养延髓神经元的方法较为成熟,所以本实验仅对神经元进行了标记,以观察所培养细胞是否为神经元。如图1所示,左上红色为MAP-2标记的神经元,左下蓝色为DAPI标记的细胞核,右图为左侧两图的叠加,可见原代培养的细胞90%以上为神经元,可进行下一步实验。

Figure 1. Confocal microscopic demonstration of medullary neurons.
2 Ang II引起神经元ROS的升高
为明确Ang II是否可升高延髓神经元的ROS水平,我们给予Ang II(1 μmol/L),37 ℃孵育30 min后,DHE荧光法检测延髓神经元ROS水平。结果显示,Ang II可显著升高延髓神经元的ROS水平,见图2。

Figure 2. Ang II increased ROS level in medullary neurons. Bar graphs summarizing ROS level after treatment with or without Ang II (1 μmol/L) for 30 min. Con: control. Mean±SEM. n=6.**P<0.01 vs con.
3 Ang II引起延髓神经元ROS升高的机制
3.1Ang II降低神经元总SOD的活性 如图3所示,Ang II显著抑制神经元总SOD的活性,提示Ang II可能通过抑制总SOD的活性,提高神经元ROS水平。

Figure 3. Activity of total SOD was inhibited by Ang II in medullary neurons. Neurons were pretreated with vehicle or Ang II (1 μmol/L) for 30 min. Con: control. Mean±SEM. n=3.**P<0.01 vs Con.
3.2CBS在神经元中的表达 如图4所示,绿色为MAP-2标记的神经元,红色为CBS阳性的细胞,蓝色为DAPI标记,叠加图中黄色为MAP-2与CBS共表达的细胞。结果显示,约90%原代培养神经元胞体均呈现黄色,提示延髓神经元是有CBS表达的,并主要表达在胞浆,为内源性H2S的研究提供了结构基础。
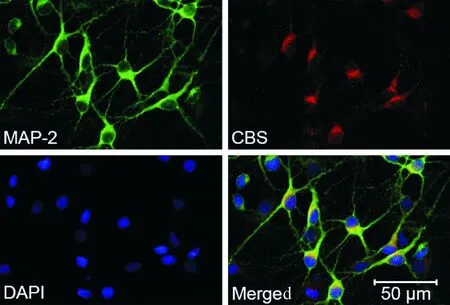
Figure 4. Expression of cystathionine β-synthase (CBS) in the rat medullary neurons.
3.3Ang II 抑制CBS mRNA的表达 如图5所示,Ang II显著抑制CBS mRNA的表达,提示Ang II可能通过抑制CBS的表达,阻断内源性H2S产生,升高神经元的ROS水平。

Figure 5. Expression of CBS mRNA was inhibited by Ang II in medullary neurons. Neurons were pretreated with vehicle or Ang II (1 μmol/L) for 30 min.Con: control. Mean±SEM. n=3.*P<0.05 vs Con.
4 内源性H2S抑制 Ang II引起的神经元ROS水平的升高
4.1单独给予不同浓度NaBu对神经元ROS水平的影响 延髓神经元单独给予NaBu (100 μmol/L、250 μmol/L和500 μmol/L),酶标仪检测结果显示,神经元胞内的ROS水平没有明显变化,见图6A,提示NaBu对于延髓神经元基础ROS水平没有显著作用。
4.2NaBu抑制Ang II引起的神经元ROS水平的变化 结果表明,100 μmol/L NaBu对Ang II引起的ROS水平没有明显作用,250 μmol/L及500 μmol/L NaBu 均可显著抑制Ang II引起的神经元ROS水平升高,并且500 μmol/L NaBu作用更明显,见图6B,提示NaBu抑制Ang II引起的神经元ROS水平的升高存在剂量依赖性。
讨 论
H2S作为第三类气体信号分子,对心血管系统有着重要的调节作用。研究表明:侧脑室注射低浓度的H2S可降低正常大鼠的血压和心率,注射不同浓度的S-腺苷甲硫氨酸(S-adenosine methionine,SAM,一种CBS激动剂)引起相同的效应并呈剂量依赖性;而高浓度的H2S则产生升压效应[8-9]。本课题组研究显示:向SHR大鼠RVLM区微量注射外源性H2S(供体NaHS)可降低血压和心率;注射CBS激动剂SAM,升高内源性H2S水平,引起相同的效应[5],且均引起ROS水平的降低。研究表明,中枢ROS介导了Ang II的心血管效应[2]。基础和临床研究也证实:血压升高常伴有氧化应激的增强[10]。
ROS是生物有氧代谢过程中的一种副产品,包括氧离子、过氧化物和含氧自由基等。过高的活性氧水平会对细胞和基因结构造成损坏。Ang II引起ROS升高的可能机制有哪些呢?中枢ROS的主要来源是NADPH氧化酶。文献报道,Ang II与AT1受体结合,可激活相关信号通路提高NADPH氧化酶的活性,升高机体的ROS水平,继而升高血压[2, 11]。在兔的侧脑室注射Ang II,可使RVLM区部分通过升高NADPH氧化酶亚单位p40phox、p47phox、p67phox和gp91phox高表达,活性氧水平升高[12]。前期工作表明,Ang II可提高延髓神经元p47phoxmRNA和p67phoxmRNA的表达量[13]。因此本实验神经元给予Ang II,作为高ROS水平的细胞模型,以与整体实验相呼应。我们首先证实了Ang II可升高延髓神经元的ROS水平。

Figure 6. NaBu inhibited ROS level in medullary neurons induced by Ang II with a dose-dependent manner. A: medullary neurons were treated with various concentrations (100, 250 and 500 μmol/L) of NaBu alone for 30 min; B: neurons were treated with vehicle, Ang II, or various concentrations (100, 250 and 500 μmol/L) of NaBu 30 min before Ang II (1 μmol/L) was used. Mean±SEM. n=6.*P<0.05,**P<0.01 vs Con (0 μmol/L Ang II and 0 μmol/L NaBu).
机体内ROS的动态平衡与SOD有着紧密的联系。SOD具有特殊的生理活性,是生物体内清除自由基的首要物质。现已证实:SOD可对抗与阻断因氧自由基对细胞造成的损害,并及时修复受损细胞,复原因自由基造成的细胞伤害。SHR大鼠RVLM区微量注射SOD可降低血压[10]。因此,ROS的升高也可能与SOD有一定关系。本研究表明,Ang II降低了延髓神经元总SOD的活性,至少部分引起了ROS的升高。
我们的整体实验中,外源性H2S(NaHS)和内源性H2S(SAM)可使SHR大鼠的ROS水平降低。前期细胞实验证实,外源性H2S(NaHS)通过抑制丝裂原激活的蛋白激酶家族中磷酸化细胞外信号调节激酶1/2蛋白的表达量,抑制Ang II引起的ROS的升高[6]。那内源性H2S是否能抑制Ang II引起的延髓神经元ROS的升高呢?
内源性H2S主要由2种酶催化生成:一种是磷酸吡多醛5-磷酸依赖性酶,包括CBS和胱硫醚γ-裂解酶(cystathionine γ-lyase, CSE)。CBS主要在海马、小脑、脑干和皮层等部位,而CSE主要分布于神经系统以外的组织。另一种是3-巯基丙酮酸硫转移酶(3-mercaptopyruvate sulfurtransferase),主要分布于肾、心、肝和脑。本课题组和近期的形态学研究显示,CBS在SD大鼠的RVLM、下丘脑室旁核等区有表达[14]。患高血压6周的Wistar 大鼠网状核神经元的CBS表达水平显著降低[15]。RVLM区是心血管交感活动的基本中枢,是维持心血管中枢紧张性活动的关键部位。本课题组前期整体实验证实,SHR大鼠RVLM区催化H2S生成的酶CBS的表达水平低于WKY大鼠[5]。本实验结果显示,CBS在延髓神经元有表达,为我们对内源性H2S的研究提供了结构基础。
CBS是中枢H2S的主要来源,Ang II与CBS之间的作用鲜有报道。本研究发现,Ang II可降低CBS mRNA的表达量,而它对于CBS蛋白水平的作用还有待进一步验证。然后我们给予CBS激动剂NaBu,升高内源性H2S,再给予Ang II观察延髓神经元ROS水平的变化。结果表明,内源性H2S(NaBu)与外源性H2S(NaHS)有相同的效应,均可抑制Ang II引起的延髓神经元ROS水平的升高。但其具体机制还需进一步探讨。
[参 考 文 献]
[1] Sun C, Sellers KW, Sumners C, et al. NAD(P)H oxidase inhibition attenuates neuronal chronotropic actions of angiotensin II[J]. Circ Res, 2005, 96(6): 659-666.
[2] Chan SH, Hsu KS, Huang CC, et al. NADPH oxidase-derived superoxide anion mediates angiotensin II-induced pressor effect via activation of p38 mitogen-activated protein kinase in the rostral ventrolateral medulla[J]. Circ Res, 2005, 97(8): 772-780.
[3] Gao L, Li Y, Schultz HD, et al. Downregulated Kv4.3 expression in the RVLM as a potential mechanism for sympathoexcitation in rats with chronic heart failure[J]. Am J Physiol Heart Circ Physiol, 2010, 298(3): H945-H955.
[4] Tay AS, Hu LF, Lu M, et al. Hydrogen sulfide protects neurons against hypoxic injury via stimulation of ATP-sensitive potassium channel/protein kinase C/extracellular signal-regulated kinase/heat shock protein 90 pathway[J]. Neuroscience, 2010, 167(2): 277-286.
[5] 于海云, 马 红, 曹银祥,等. 硫化氢对SHR大鼠的中枢心血管效应及其机制探讨[J], 中国分子心脏病学杂志, 2011, 11(4): 235-240.
[6] 马 红, 于海云, 于 燕,等. H2S抑制AngⅡ引起的神经元活性氧水平升高的机制研究[J]. 中国病理生理杂志, 2012, 28(5): 865-869.
[7] Cao Q, Zhang L, Yang G, et al. Butyrate-stimulated H2S production in colon cancer cells[J]. Antioxid Redox Signal, 2010, 12(9): 1101-1109.
[8] Dawe GS, Han SP, Bian JS, et al. Hydrogen sulphide in the hypothalamus causes an ATP-sensitive K+channel-dependent decrease in blood pressure in freely moving rats[J]. Neuroscience, 2008, 152(1): 169-177.
[9] Ufnal M, Sikora M, Dudek M. Exogenous hydrogen sulfide produces hemodynamic effects by triggering central neuroregulatory mechanisms[J]. Acta Neurobiol Exp (Wars), 2008, 68(3): 382-388.
[10] Keith M, Geranmayegan A, Sole MJ, et al. Increased oxidative stress in patients with congestive heart failure[J]. J Am Coll Cardiol, 1988, 31(6): 1352-1356.
[11] Hirooka Y, Sagara Y, Kishi T, et al. Oxidative stress and central cardiovascular regulation: pathogenesis of hypertension and therapeutic aspects[J]. Circ J, 2010, 74(5): 827-835.
[12] Gao L,Wang W, Li YL, et al. Sympathoexcitation by central Ang II: roles for AT1 receptor upregulation and NAD(P)H oxidase in RVLM[J]. Am J Physiol Heart Circ Physiol, 2005, 288(5): H2271-H2279.
[13] 马 红. H2S抑制Ang II引起的延髓神经元活性氧水平升高的机制研究[D]. 上海:复旦大学, 2012.
[14] Guo Q, Jin S, Wang XL, et al. Hydrogen sulfide in the rostral ventrolateral medulla inhibits sympathetic vasomotor tone through ATP-sensitive K+channels[J]. J Pharmacol Exp Ther, 2011, 338(2): 458-465.
[15] Chertok VM, Kotsiuba AE. Immunocytochemical characterization H2S-positive neurons in the nuclei of bulbar cardiovascular center in the development of renovascular hypertension[J]. Vestn Ross Akad Med Nauk, 2012, (4): 50-54.
