五节芒与荻人工杂交种F1群体开花物候与生殖特性研究
2014-04-10艾辛朱玉叶蒋建雄龙卫李莎莎易自力
艾辛,朱玉叶,蒋建雄,龙卫,李莎莎,易自力*
(1.湖南农业大学园艺园林学院,湖南 长沙410128;2.湖南农业大学生命科学学院,湖南 长沙410128;3.湖南省常德市农业科学研究所,湖南 常德415000)
芒属(Miscanthus)植物作为极具开发潜力的能源植物之一备受关注[1-4]。中国是芒属植物重要的起源与分布中心,中国有7个种[5],其中芒(M.sinensis)、五节芒(M.floridulus)、荻(M.sacchariflorus)、南荻(M.lutarioriparus)4个种作为能源植物开发利用的价值最大,资源分布范围广、蕴藏量大,称为芒属能源植物(Miscanthusenergy plant,MEP),其多样性丰富[6-9]。芒属植物的4个种各自具有优良的特性,但作为能源植物直接利用都存在不足,通过种间杂交将分散在各个种中的优良性状聚合在一起,选育出生物产量高,适应不良环境能力强的新品种是芒属能源植物育种的主要方向。荻是我国芒属植物中分布纬度最北、经度最西的种类,适应恶劣生态环境的能力强,地下根状茎低于-30℃也能安全越冬,而地上部分在低温来临前已自然衰老。研究表明,荻还具有耐中度干旱和瘠薄的能力[10-15],耐受盐碱的能力较强[16],对重金属污染的土壤也有一定的修复能力[17],其根系的扩张能力有利于水土固定[18],综合评价,荻是一种适应恶劣环境能力强但生物产量较低的芒属能源植物。五节芒主要生长在我国长江中下游流域的广大地区,对其生态生物学特性[19]、遗传特性[20-21]、生态修复[22-23]及利用价值[24-25]等方面所做的研究工作表明,五节芒生物产量高,具有较好的能源利用开发前景。但五节芒具有不易衰老的特性,虽然利用催熟剂能促进其衰老[26],但这种方法对五节芒今后的生长是否有不良影响还有待观察。芒属能源植物的自然衰老性状是一个重要性状,随着植物的衰老,植物体内的水分和矿物质的含量明显减少,有利于收获、储藏和燃烧[27]。为了充分利用荻和五节芒优良特性,克服各自的缺点,开展了荻和五节芒的种间杂交工作,首次在全球获得了荻和五节芒的种间杂交种群体[28],杂交种在生长速度,生物产量、耐旱、耐瘠薄等方面充分地显示了远缘杂种的优势。植物远缘杂交往往存在不亲和、杂种不育和杂种后代不稔三大障碍,杂交试验和天然种间杂交种的研究表明芒属植物种间杂交的障碍不明显[29],目前欧美国家普遍利用的能源植物“奇岗”就是荻和芒的天然三倍体杂交种[30]。芒属植物种间杂交种虽然具有较大的杂种优势,但通过对杂交种后代基因的分离重组和选择,能实现优良性状的聚合,筛选出性状优良的新品种,对杂交种的开花物候和生殖特性的了解对开展杂交种的进一步遗传改良研究非常重要,但到目前为止,有关芒属植物种间杂交种在开花物候和生长特性方面的研究还未见报道,本研究利用五节芒和荻的人工杂交种群体进行开花物候、结实能力和花粉母细胞减数分裂行为等方面的观察,旨在为今后进一步开展芒属植物种间、种内及亲子代间有性杂交育种积累基础资料。
1 材料与方法
1.1 供试材料与试验地环境条件
试验材料为本研究者创建的五节芒和荻的种间杂交种F1群体,经过SSR分子标记和田间形态学鉴定的五节芒与荻杂种植株和2个亲本。亲本荻采集于甘肃泾川县窑店镇,亲本五节芒采集于湖南邵阳市洞口县。杂种植株群体为112株,只有103株在大田种植。2个亲本均为二倍体(2n=38)。试验材料于2011年种植于浏阳芒草试验基地,植株生长旺盛,冬季衰老后割除地上部分,安全越冬。第1年由于受栽培时间的影响,当年8月开花,不作为物候期信息进行分析,2012年进行正式的开花物候期和生殖特性观测,2013年重复观测1次物候期和其他农艺学性状。
浏阳芒草试验基地坐落在湖南省浏阳市永和镇石佳村,栽培地为山地,经过机械施工平整,土质属于贫瘠的黄壤土,气候条件为中亚热带季风湿润气候[31]。
1.2 实验方法
1.2.1 杂交种F1群体开花物候观察与开花数统计 从2012年5月初开始观察杂交种和2个亲本的开花情况,观测指标主要有:始花日期、开花持续时间和终花日期。个体(株丛)始花日期为单株第1朵花开放的日期,终花日期为单株最后1朵花开放的日期。个体总花期长度为第1朵花开到最后1朵花开的时间。群体水平的开花物候参数以所观测的全部个体的平均值计算,其中,始花日指第1个植株开花日期。参照Dafni[32]的方法,群体水平上5%的个体开花时视为始花,50%的个体达到开花高峰时视为群体开花高峰期,95%的植株开花结束时视为群体花期结束。芒属植物是一种多分蘖植物,形成丛生状,大部分的分蘖都能孕穗开花,此处所说的个体水平是指杂交种和它的亲本株丛。2013年继续观测,但只观测每个株丛的孕穗及始花日期,与2012年数据进行比较验证,由于花期基本一致,而分蘖数又太多,未进行开花花序数的记数,故相关计算以2012年的数据为准。
用同步指数(synchrony index,Si)检测孕抽穗和开花同步性高低,具体方法见参考文献[33],同步指数计算公式如下:

式中,ej表示个体i和j花期重叠时间(d),fi表示个体i开花的总时间(d),n表示群体中个体总数。Si的变异范围为0~1,“0”表示种群内个体花期无重叠,“1”则表示完全重叠,所有个体的平均值为该群体的开花同步性指数。
从5-10月对F1各株丛植株每月开花分蘖数进行统计,根据植株每月开花分蘖数的数量计算植株的平均开花振幅(mean flowering amplitude),每月开花比例=(每月开花分蘖数/开花分蘖总数)×100%。
1.2.2 花粉育性观测 采用I2-KI溶液进行花粉育性的测定。在亲本和杂交种的开花期,每天早上7点至10点之间选取每株当天开花的花序。每次随机从花序上选取花药未开裂散粉的小穗,从中取出3个花药放在载玻片上。在载玻片上滴1~2滴I2-KI溶液,用镊子将花药轻轻捣碎,去掉肉眼可见的杂质,加上盖玻片,在Olympus显微镜下(100×)观察。全部染成棕黑色的花粉被认为是可育花粉,不染色或染色很浅的花粉为不育花粉。每份材料制作3个效果优良的制片,每片观察3个视野,每个视野能观察花粉数30粒以上。可育花粉百分率=(染色花粉数/观测花粉数)×100。
1.2.3 自花授粉和异花授粉结实率的测定 每株丛选取3个花序,用定制的羊皮纸袋分别对其套袋,其后不再做任何处理。同时每株丛随机选取3个花序作为对照让其自然授粉。待种子完全成熟时,将套袋处理的花序和自然授粉的花序分开收集,将3个自交花序上小穗混合均匀,随机取300个小穗进行计数,重复3次,自然授粉的花序照此法同样处理,分别统计结实率,结实率=(结实数/300)×100%。
1.2.4 杂交种F1植株花粉母细胞减数分裂的显微观察 取材与后处理:在亲本和杂交种的开花期,于早上7点左右在田间取材,以旗叶与下一叶叶耳间距为标准,选取不同长度的幼穗用卡诺氏固定液(无水乙醇∶冰醋酸=3∶1)固定24h左右,用95%酒精清洗3次,转入70%酒精中低温4℃保存备用。
制片:用镊子将1朵小花中雄蕊取出,放在洁净的载玻片上,滴1滴45%的醋酸将其软化几秒,擦去多余液体,再滴1~2滴的改良卡宝品红染液[34]进行染色,加盖玻片进行压片,吸水纸吸去多余液体。
镜检:在Olympus普通光学显微镜(400×或1000×)下镜检,寻找减数分裂中的各个分裂相并拍照。减数分裂各时期分别选取30个视野,至少观察200个细胞,统计各时期的染色体行为异常率,异常率(%)=(染色体行为异常细胞数/各时期观察的细胞总数)×100%。
1.3 数据统计与分析
采用Microsoft Excel进行花粉育性和结实率的描述性统计分析,用DPS 6.5进行开花物候、花粉育性与结实率的相关性分析。
2 结果与分析
2.1 五节芒与荻杂交种F1群体的开花物候
个体和群体的开花物候特征如表1所示,F1群体一般为5月下旬开始孕穗,6月中旬开始开花,花期持续到10月底,花期持续时间长。F1个体在始花日期和终花日期上略有不同,始花日期为6月7日至6月21日之间,终花期为10月11日至10月27日之间,开花情况具很高同步性。由图1可见,亲本五节芒的花期分为2个时期,第1个开花高峰出现在7月(开花分蘖数占全年开花分蘖数比例为43.75%),第2个开花高峰出现在9月(开花分蘖数比例为50.00%),2个花期间分隔明显,不表现为连续性;亲本荻只在6月(开花比例为100%)出现1个开花高峰;F1杂种从6月上旬开始连续开花直至10月下旬结束,但仍然可以分为2个阶段即2个开花高峰,呈双峰曲线,第1次开花高峰出现在6月(开花分蘖数平均值为36.34%)与亲本荻重合,1个在9月(开花分蘖数平均值为39.38%)与亲本五节芒重合(图1)。从孕穗期和始花期的时间看,杂交种的始花期偏向于早花的荻亲本,66.07%的个体(株丛)花期与亲本荻相近,31.25%的个体花期与亲本五节芒相近,只有2.68%的花期晚于晚花的亲本,说明早花性状的遗传力高。
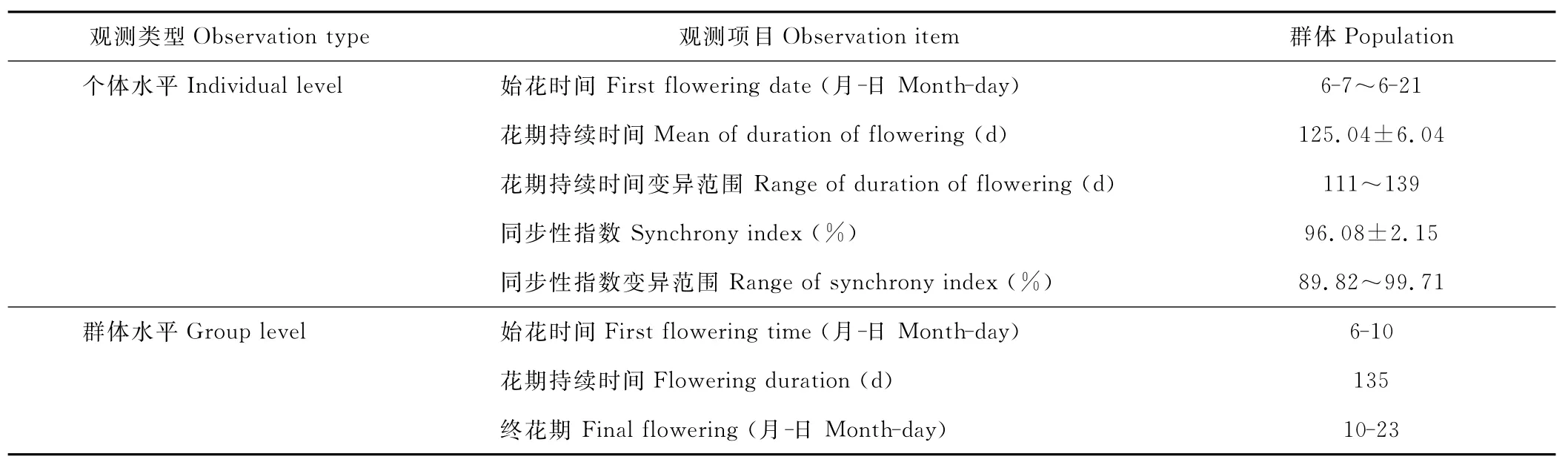
表1 F1群体个体水平(n=103)和群体水平的开花物候Table 1 Flowering phenology characteristics of F1population and individual plants(n=103)
2.2 杂交种F1植株花粉育性研究
利用I2-KI染色法测定2个亲本和103株F1植株的花粉育性(表2),发现芒属植物花粉经I2-KI染色后呈较深的颜色,说明亲本和F1植株花粉中淀粉的含量比较高。亲本荻的花粉染色率为(96.11±2.05)%,亲本五节芒第1次开花高峰花粉染色率为(75.38±12.44)%,第2次开花高峰花粉染色率为(92.32±4.70)%,明显高于第1次开花高峰花粉染色率。F1群体植株平均花粉染色率低于2个亲本,2次开花高峰的花粉染色率分别为(50.79±18.61)%,(65.42±14.71)%,第2次开花高峰时的花粉染色率大于第1次开花高峰时的花粉染色率。F1群体个体间有较大的差异,2次开花高峰可育花粉率的变异系数分别为36.64%,22.48%,杂交种育性降低说明了荻和五节芒的亲缘关系相对较远,可能存在较大的生理差异。

图1 亲本荻、五节芒和杂种F1个体开花振幅曲线Fig.1 Flowering amplitudes curves of F1population and its parents M.floridulus and M.sacchariflorus
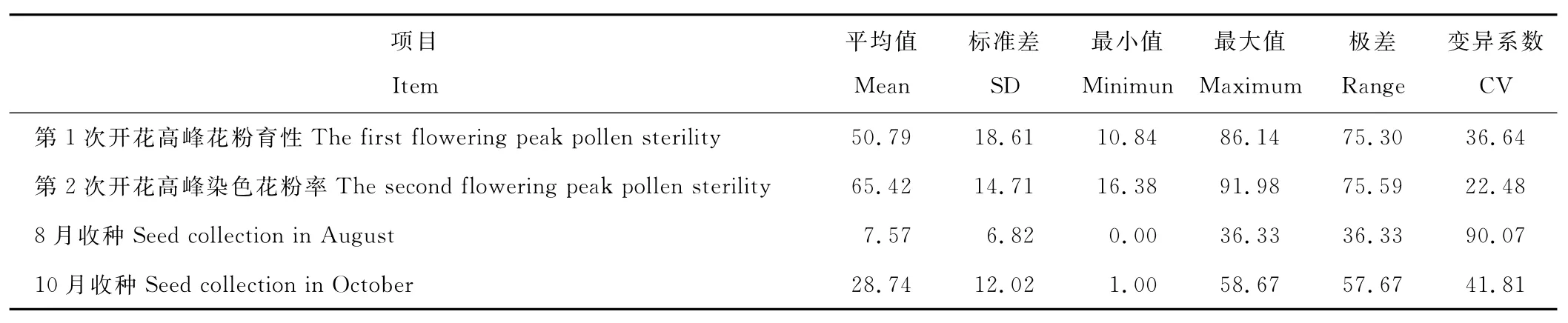
表2 杂交种F1群体花粉育性与结实性Table 2 Pollen fertility and natural seed setting rate of F1population %
2.3 杂交种自交结实率与天然结实率测定
荻、五节芒和它们的杂交种植株的花序套袋自交结实率均为0,说明荻和五节芒杂交种F1也是高度自交不亲和。开放授粉的情况下,荻的天然结实率为0,五节芒第1次开花高峰期的花序结实率极低,为0.33%,第2次开花高峰期的花序结实率为22.00%。种植在湖南农业大学校园内芒属植物资源圃的2个亲本却有很高的天然结实率[亲本荻为(57.22±11.69)%,亲本五节芒为(62.94±4.47)%]。F1群体内株间的天然结实率有较大的差异(表2),第1次开花高峰期的花序平均天然结实率为(7.57±6.82)%,最低为0.00%,最高为36.33%,变异系数为90.07%,第2次开花高峰期的花序平均天然结实率为(28.74±12.02)%,最低为1.00%,最高为58.67%,变异系数为41.81%,说明杂交种株间有较大的遗传差异,群体多样性高。

表3 F1植株个体水平的物候指标及结实率的相关分析Table 3 Correlation analysis between flowering phenology parameters and seed setting rate of F1individual plants
2.4 杂交种开花物候指数与结实率的相关性
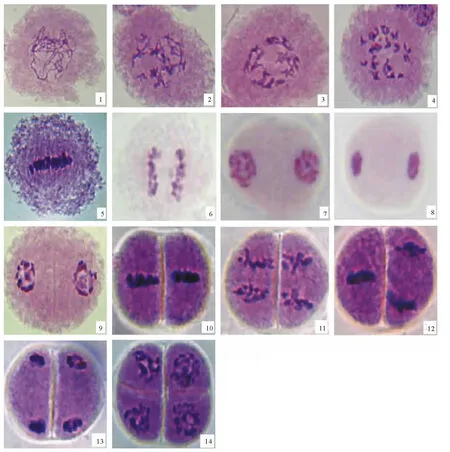
图2 五节芒与荻人工杂交种F1花粉母细胞减数分裂(在400×下拍摄)Fig.2 Meiosis of pollen mother cell of F1plants(400×)
五节芒与荻人工杂交种F1植株个体水平的始花时间、花期持续时间、开花数目与结实率的相关分析显示(表3),开花数与花期持续时间呈极显著正相关,与始花时间和结实率呈正相关关系;花期持续时间与始花时间呈极显著负相关,与结实率呈正相关关系;始花时间与第1次开花高峰结实率呈负相关,与第2次开花高峰结实率呈显著负相关关系。这表明开花数目多的个体比开花数目少的个体花期持续时间更长,结实率更高;花期持续时间越长的个体,始花时间越早,结实率更高;始花时间早的个体比始花时间晚的个体花期持续时间更长,第2次开花高峰后的结实率更高。
2.5 杂交种花粉育性与结实率的关系
杂交种花粉的染色率与结实率的关系并不密切,第1次开花高峰的花粉染色率与结实率之间为负相关(r=-0.1473),第2次开花高峰花粉育性与结实率之间虽然为正相关(r=0.1754),但也未达到显著性水平,花粉I2-KI染色率与植株结实率之间没有相关性,其可能原因是I2-KI染色法不能有效地反映杂交种的花粉育性和活力水平,且由于芒属植物自交不亲和,自花花粉是否可育与雌蕊能否受精结实也没有关系,雌蕊的发育是否正常值得关注。
2.6 杂交种F1植株花粉母细胞减数分裂观察
利用根尖压片法和流式细胞仪观测亲本荻和五节芒的染色体数目,2个亲本的倍性均为二倍体,花粉母细胞减数分裂未见异常。选取叶枕距范围在3~6cm的健壮幼穗可以获得较好的减数分裂时期的各个分裂相。如图2所示,观测结果表明F1植株的花粉母细胞减数分裂过程基本正常,胞质分裂为连续型,即减数第1次分裂后产生细胞板,将母细胞分为2个子细胞,即二分体。接着进行第2次分裂,形成四分体,四分体为两侧对称型。少数花粉母细胞在第2次减数分裂后期出现染色体分裂不同步(图2-12),但这一现象出现的比例很小,不同步差异也不大,到第2次分裂末期又趋于同步,即不会导致出现三分体的现象。
个别花粉母细胞减数分裂过程中染色体的异常行为(图3)包括:在中期I出现部分染色体没有排列在赤道板上,而是随机分布于赤道板附近(图3-15),行为异常率为1.5%;后期I正常染色体已趋向两级,而落后染色体则分布于两级之间形成染色体桥(图3-16),行为异常率为1;末期I仍可见染色体片段位于两级之间(图3-17),行为异常率为0.5%。
3 讨论
根据中国植物志[35]记载,荻在原产地的花果期为每年8-10月,五节芒的花果期为每年的5月到10月。本试验的亲本荻在浏阳基地观察到的始花期在6月上中旬,花期呈单峰曲线,而在湖南农业大学芒草基地其花期延续至9月上旬,开花花序数呈双峰曲线,高峰分别出现在6月上旬和9月上旬。五节芒花果期从7月一直持续到10月,呈双峰曲线,高峰分别出现在7和9月。出现这些情况的原因在于荻是北种南移,并且有研究表明荻是短日照响应植物[36],因此出现了提前开花的现象,但在浏阳基地没有出现二次开花高峰,可能是由于土壤贫瘠缺水所致,没有持续新长出的分蘖,吴文新和王洪铭[37]研究表明,肥料与水分因素对菊花(Dendranthemamorifolium)的开花期有很大影响。五节芒从7月开始开花,出现2次开花高峰,这与朱明东等[29]发现五节芒的生殖分蘖中下部的节位会发生第2次抽穗开花的现象相吻合。五节芒与荻的杂交种F1群体呈双峰曲线,花期持续时间长,同步性指数高,季孔庶等[38]在进行鹅掌楸属(Liriodendron)树种物候观测时也发现杂种花期持续时间比双亲的长,并且杂种和亲本的花期之间有较长交叉时间。
开花物候对植物的生殖成功具有一定影响[39-41]。Augspurger[42]认为,植物的开花数是决定其结实率的重要因子,在观察的6种新热带灌木的开花物候中,有5种植物的个体开花数与结实率呈正相关。肖宜安等[43]研究发现濒危植物长柄双花木中,始花日期与结实率均呈显著负相关关系,始花日期早的比始花日期迟的结实率高,而开花数与结实率表现出线性正相关关系。通过对浏阳基地五节芒与荻人工杂种个体水平的花期持续时间、开花数与结实率的关系及相关分析发现:开花数和花期持续时间分别与结实率存在正相关关系,此结果与李新蓉等[41]和Augspurger[42]的研究结果一致。由于芒属植物存在自交不亲和现象,花粉寿命较短,柱头可授时间较长的特性[44],五节芒与荻人工杂种在群体开花数多,花期持续时间较长的情况下,增加了接受其他基因型花粉的机会,从而提高结实的机会。
2个亲本均是二倍体,而且减数分裂正常,F1植株减数分裂基本正常,但因为是种间杂交种,也有个别花粉母细胞出现了染色体异常行为,染色体行为异常率均低于2.0%,这说明五节芒与荻种间基因组具有很高的同源性,可以相互配对,说明五节芒与荻有很近的亲缘关系。张春等[45]通过对拟鹅观草属(Pseudoroegneria)与鹅观草属(Roegneria)和披碱草属(Elymus)属间及种间杂种的花粉母细胞减数分裂观察,确定了披碱草属种的St染色体组与Y染色体组具有一定程度的同源性,亲缘关系较近,而与H染色体组的关系较远。对于五节芒与荻种间杂种的花粉母细胞减数分裂行为研究还未见报道,有文献指出[46]芒属的19个连锁群与高粱(Sorghumbicolor)的10个连锁群以2∶1呈相同的线性关系,表明芒属植物基因组中存在着完整的全基因组的染色体复制。而高粱染色体基数为10[47],本实验也不能鉴别杂种F1中配对的染色体为同源配对还是异源配对,可借助基因组原位杂交(GISH)技术加以鉴别[48],可能也存在一些片段的重排和变异,导致种间杂种染色体在减数分裂过程中的异常行为,其减数分裂过程还有待进一步深入的研究。从杂交种的花粉育性来看,杂交种的花粉育性普遍降低,说明两者由于亲缘关系的差异有可能影响到花粉发育,这与杂交种染色体在减数分裂过程的异常行为是否有某种关系,也有待今后进一步的研究。
[1] Sang T,Zhu W X.China’s bioenergy potential[J].Global Change Biology Bioenergy,2011,3(2):79-90.
[2] Hastings A.Future energy potential ofMiscanthusin Europe[J].Global Change Biology Bioenergy,2009,1(2):180-196.
[3] Lewandowski I,Clifton-Brown J,Scurlock J M O,etal.Miscanthus:European experience with a novel energy crop[J].Biomass and Bioenergy,2000,19(4):209-227.
[4] Lewandowski I,Scurlockb J M O,Lindvallc E,etal.The development and current status of perennial rhizomatous grasses as energy crops in the US and Europe[J].Biomass and Bioenergy,2003,25(4):335-361.
[5] Chen S L,Renvoize S A.MiscanthusAndersson[A].Flora of China,Vol.22[M].St.Louis.:Missouri Botanical Garden Press,2006:581-583.
[6] 薛德,肖亮,艾辛,等.五节芒表型性状和SSR标记遗传多样性分析[J].草业学报,2012,21(5):96-106.
[7] 肖亮,蒋建雄,易自力,等.五节芒种质资源的表型多样性分析[J].湖南农业大学学报(自然科学版),2013,39(2):150-154.
[8] 刁英,胡小虎,郑兴飞,等.利用SRAP和ISSR标记分析五节芒(Miscanthusfloridulus)的遗传多样性[J].武汉大学学报(理学版),2010,(5):578-583.
[9] 肖亮,蒋建雄,易自力,等.广西省芒野生居群表型多样性研究[J].草业学报,2013,22(4):43-50.
[10] 聂刚,张新全,黄琳凯,等.中国西南区野生芒居群表型变异研究[J].草业学报,2013,22(5):52-61.
[11] 黄平,左海涛,韩烈保,等.拔节期水分胁迫对荻生长和生物质特性的影响[J].草地学报,2007,15(2):153-157.
[12] 黄杰,黄平,左海涛.栽培管理对荻生长特性及生物质成分的影响[J].草地学报,2008,16(6):646-651.
[13] 侯新村,范希峰,左海涛,等.氮肥对挖沙废弃地能源草生长特性与生物质产量的影响[J].草地学报,2010,18(2):268-273,279.
[14] 李强,李加鹏,何淼.荻实生苗生物构件对土壤水分胁迫的响应[J].草业科学,2013,30(6):893-897.
[15] 何淼,赵保成,李强,等.PEG胁迫对芒和荻种子萌发的影响[J].草业科学,2013,30(4):577-582.
[16] 宗俊勤,陈静波,聂东阳,等.我国不同地区芒和荻种质资源抗盐性的初步评价[J].草地学报,2011,19(5):803-807.
[17] 张杰,周守标,黄永杰,等.能源植物荻对铜胁迫的耐性和积累特性[J].水土保持学报,2013,27(2):168-172,188.
[18] 杨朝东,张霞,万勇,等.荻草根系统扩展能力和固土作用的研究[J].长江大学学报(自然科学版),2009,6(3):19-23.
[19] 陈慧娟,宁祖林,张卓文.五节芒生物学特性及能量生产动态变化[J].草业学报,2012,21(6):252-257.
[20] 邓果特,刘清波,蒋建雄,等.五节芒基因组大小测定[J].植物遗传资源学报,2013,14(2):339-341,346.
[21] 陈少风,何俊,周朴华,等.芒和五节芒的核型研究[J].江西农业大学学报,2008,30(1):123-126.
[22] 秦建桥,夏北成,赵鹏,等.五节芒(Miscunthusfloridulus)不同种群对镉积累与转运的差异研究[J].农业环境科学学报,2011,30(1):21-28.
[23] 秦建桥,赵华荣,胡萌,等.铅胁迫下不同生态型五节芒(Miscanthusfloridulus)的抗氧化系统的差异研究[J].生态环境学报,2011,20(3):525-531.
[24] 陈慧娟.五节芒生物量及其热值的动态研究[D].武汉:华中农业大学,2009.
[25] 龚道生.五节芒栽培花菇技术与效益的分析[J].防护林科技,2011,(3):40-42.
[26] 易镇邪,王禹,王学华,等.3种催熟剂对芒属植物的催熟效果比较[J].草业科学,2013,30(7):1052-1056.
[27] Robson P,Mos M,Clifton-Brown J,etal.Phenotypic variation in senescence inMiscanthus:towards:optimising biomass quality and quantity[J].Bioenergy Research,2012,5(1):95-105.
[28] 朱玉叶,艾辛,蒋建雄,等.五节芒与荻人工杂交种的创建与鉴定研究[J].中国草地学报,2013,35(2):31-36.
[29] 朱明东,蒋建雄,肖亮,等.基于形态性状及Adhl基因序列的芒与五节芒自然杂交现象研究[J].草业学报,2012,21(3):132-137.
[30] 席庆国,洪浩.外来植物奇岗的生物学特征[J].草业科学,2008,25(2):26-28.
[31] 李良勇,谢鹏飞,刘峰,等.湖南浏阳烟区气候土壤因素和烟叶质量特点[J].湖南农业大学学报(自然科学版),2006,32(5):497-501.
[32] Dafni A.Pollination Ecology[M].New York:Oxford University Press,1992:1-57.
[33] Mcintosh M E.Flowering phenology and reproductive output in two sister species of Ferocactus(Cactaceae)[J].Plant Ecology,2002,159:1-13.
[34] 冯静仪,张振宇,李兴诗,等.一种优良的减数分裂实验材料及其制片[J].生物学通报,1986,(09):18.
[35] 刘亮.芒属,双药芒属,荻属[A].陈守良,庄体德,方文哲,等.中国植物志[M].北京:科学出版社,1997:4-26.
[36] Jensen E,Robson P,Norris J,etal.Flowering induction in the bioenergy grassMiscanthussacchariflorusis a quantitative short-day response,whilst delayed flowering under long days increases biomass accumulation[J].Journal of Experimental Botany,2013,64(2):541-552.
[37] 吴文新,王洪铭.菊花花期调控技术的研究概况及展望[J].福建农业科技,2001,3:21-23.
[38] 季孔庶,杨秀艳,杨德超,等.鹅掌楸属树种物候观测和杂种家系苗光合日变化[J].南京林业大学学报(自然科学版),2002,26(6):28-32.
[39] Ollerton J,Lack A.Relationships between flowering phenology,plant size and reproductive success inLotuscorniculatus(Fabaceae)[J].Plant Ecology,1998,139(1):35-47.
[40] Ollerton J,Diaz A.Evidence for stabilizing selection acting on flowering time inArummaculatum(Araceae):the influence of phylogeny on adaptation[J].Oecologia,1999,119(3):340-348.
[41] 李新蓉,谭敦炎,郭江.迁地保护条件下两种沙冬青的开花物候比较研究[J].生物多样性,2006,14(3):241-249.
[42] Augspurger C K.Phenology,flowering synchrony,and fruit set of six neotropical shrubs[J].Biotropical,1983,15(4):257-267.
[43] 肖宜安,何平,李晓红.濒危植物长柄双花木开花物候与生殖特性[J].生态学报,2004,24(1):14-21.
[44] 赵元杰.芒繁育系统研究[D].长沙:湖南农业大学,2010.
[45] 张春,王晓丽,于海清,等.拟鹅观草属与鹅观草属和披碱草属属间及种间杂种的细胞学研究[J].草业学报,2009,18(3):86-93.
[46] Ma X F,Jensen E,Alexandrov N,etal.High resolution genetic mapping by genome sequencing reveals genome duplication and tetraploid genetic structure of the diploidMiscanthussinensis[J].PLoS One,2012,7(3):1-11.
[47] 詹秋文,高丽,张天真.苏丹草与高粱染色体核型比较研究[J].草业学报,2006,15(2):100-106.
[48] 于卓,云锦风,马有志,等.加拿大披碱草×野大麦三倍体杂种染色体的分子原位杂交鉴定[J].遗传学报,2004,31(7):735-739.
